
Uptake of Micro and Macronutrients in Relation to Increasing Mn Concentrations in Cistus salvifolius L.Grown in Hydroponic Cultures
2019-01-09 02:37FiglioliMemoliMaistoSpagnuoloGiordanoLeidiRossiniOliva
F.Figlioli,V.Memoli,G.Maisto,V.Spagnuolo†,S.Giordano,E.O.Leidi,S.Rossini Oliva
1Dipartimento di Biologia,Università Federico II,Via Cinthia 4,80126 Napoli,Italy
2IRNAS-CSIC,Av R.Mercedes 10,41012 Seville,Spain IRNAS-CSIC,Av R.Mercedes 10,41012 Seville,Spain
3Dpto.Biología Vegetal y Ecología,Avda.Reina Mercedes,41080 Sevilla,Spain
Keywords Manganese Plant metal stress Plant mineral uptake SEM-XS ray microanalysis
Abstract Mining and smelting activities can alter the ecosystem degrading vegetation and landscape,causing loss of soil fertility and changes in hydrology and microclimate.The mining area of Rio Tinto is one of the largest metallic sulfide deposits in the world,extending to southern Portugal and the Rio Tinto region(Huelva,SW Spain).Soils,characterized by low pH,are strongly impoverished in macro-and micronutrients essential to the plant metabolism and contain very high concentrations of As,Cu,Fe,Mn,and Pb.The aim of this study was to evaluate the effects of increasing Mn concentrations(0,50,100,200,and 300 mg/L)on the uptake of a set of micro and macro nutrients in Cistus salvifolius L.,a species native of the Rio Tinto region.The plants,grown in hydroponic culture,were analyzed by AAS for elemental content and by SEM-XS ray microanalysis for element localization.The results of this study showed a stunted growth and ultrastructuralalterations in the root of C.salvifolius,with the most evident damages occurring at the highest Mn concentration.Chemical analyses confirm that the higher the concentration in culture medium,the higher the uptake of Mn in plant tissue;both lower and higher Mn concentrations influence the absorption of other essential nutrients,as Fe,Zn,K,and Mg.The visible state of stress observed in plants grown with addition of 300 ppm Mn may therefore be due to such variations in the absorption of micronutrientsand/orto the Mn itself.Futurestudies shouldfocuson possible synergisticandantagonisticactivities of Mnversus otheressential elements for proper plant development.
1 Introduction
Mining and smelting activities induce landscape degradation,vegetation disruption,soil fertility loss,and hydrological and microclimate changes,deeply disturbing the ecosystems(Runolfsson and Arnalds,2004).
The abandoned mining area of Rio Tinto is included in the Iberian Pyrite Belt(IPB)one of the largest metallic sulfide deposits in the world,extending in Southern Portugal and the Rio Tinto district(Huelva,SW Spain).The exploitation of this region has been going on for about 5000 years,producing a total of about 1600 million metric tons of waste material(Davis et al.,2000);the area is currently occupied by open pits,tailing deposits and other mining wastes.As a result of those activities and intensive deforestation,natural landscape disruption and vegetation lack occur at large areas of the region.The average mineralogical composition found in the area consists of pyrite(83.1%),sphalerite(5.4%),galena(2.1%),chalcopyrite(1.4%),and arsenopyrite(0.9%),the remaining fraction(7.1%)corresponds to unproductive minerals(Almodovar et al.,1998).The area has a Mediterranean climate,with annual rainfall from 600 to 800 mm and mean annual temperature of 18◦C.Summers are very hot and dry and rainfall occur mostly during autumn and winter.
Usually,soils developing on mine wastes are spontaneously colonized by pioneer plant species,which provide an important contribution for land recolonization and natural rehabilitation(Anawar et al.,2013).These plants tolerate the increased availability of potentially toxic metals in the soils through genetically-based mechanisms which allow either to prevent root metal uptake and translocation or detoxification and compartmentalization of absorbed metals.The tolerance in these plants is not metal specific,varying among different genotypes(ecotypes,physiotypes or races)of the same species(Bargagli,1998).Several families of vascular plants,such as Caryophyllaceae,Cyperaceae and Ericacaeae,include species with morphological and physiological plasticity able to evolve tolerance mechanisms and,to a certain extent,to modulate their evolution according to substrate characteristics(Rossini-Oliva et al.,2016).A number of studies(Kidd et al.,2004;Freitas et al.,2004;Santos et al.,2009;de la Fuente et al.,2010;Abreu et al.,2012;Jiménez et al.,2011)show that several Cistus species(e.g.C.salvifolius L.,C.monspeliensis L.,C.albidus L.,C.crispus L.,C.populifolius L.and C.ladanifer L.)are able to survive in very hostile habitats.
Manganese is an essential element for plant growth,activating some enzymes involved in citric acid cycle(tricarboxylic acid cycle);besides,a central role of manganese cluster complexes in oxidation of water to oxygen has been reported(Dharmendra et al.,2013).However,its excess,especially in acidic soils,can affect plant survival disturbing physiological functions and mineral uptake(Kochian et al.,2004;Ducic and Polle,2005).Manganese in plants participates in the structure of photosynthetic proteins and enzymes.Its deficit is dangerous for chloroplasts because it affects the water-splitting system of photosystem II(PSII),which provides the necessary electrons for photosynthesis(Buchanan et al.,2000).However,its excess seems also to be particularly damaging to the photosynthetic apparatus(Mukhopadhyay and Sharma,1991).Toxic Mn levels fall in the range of 1000-12000 mg kg-1,depending on the plant species;some species have been found with Mn contents in the range 1000-5000 mg kg-1both on soils with Mn concentrations higher than 1%and on soils with lower Mn concentrations.Ultrama fic soils may have 1000-5000 mg kg-1,which is not regarded as strongly abnormal.Manganese toxicity is favored in acid soils(Pendias and Pendias,1992);with low pH values,the amount of exchangeable Mn–mainly Mn2+–increases in the soil solution.
In the present study the effect of increasing Mn concentrations on the uptake of selected micro and macronutrients,such as Fe,Zn,K,Mg,and Mn itself,was evaluated in Cistus salvifolius,a plant native of the Rio Tinto region.The aim of the experiment was to assess if mineral nutrition is affected by Mn concentrations in these plants,naturally growing in soil enriched in Mn.To this aim,the plants were cultured in hydroponic medium and analyzed by AAS for their element content;in addition,SEM microscopy X-ray spectrometer equipped was applied to follow Mn distribution in plant tissues and reveal eventual anatomical alterations possibly related to Mn stress.
2 Materials and methods
2.1 Plant material
Cistus salvifolius L.is a shrub belonging to the Cistaceae family,typical of Mediterranean scrub.It is a shrubby plant not exceeding 50-60 cm height(Farley and McNeilly,2000).Combined with other bushes,it can spread several meters and form impenetrable clusters.Itis resistant to prolonged drought conditions,and not demanding about soil pH.It grows well in neutral,slightly limestone or slightly sandy soils.The plant prefers a sunny exposure,but it also suits partially shady conditions,so it can also be found in open-wood forests;it is also well adapted to areas exposed to environmental disturbances such as recurrent fires and mine contamination.
2.2 Experimental design and plant culture
The seeds of C.salvifolius were treated in a stove at 40◦C for 24 hours to activate seed germination,and subsequently seeded on humid paper in the dark.The plantlets were then transferred and grown in hydroponic solution with a modified Hoagland solution(Hoagland and Arnon,1950;modified according to Rossini-Oliva et al.,2016).After obtaining seedlings of at least 5 cm long,the samples were exposed to different Mn concentrations for a period of 15 days.We tested the effect of 4 different Mn concentrations added to the culture medium in the form of MnSO4to obtain final concentrations of 50 ppm,100 ppm,200 ppm,300 ppm;Mnuntreated plants were also cultured in the same conditions and used as control.The environmental conditions provided were:16/8 hrs light/darkness;ambient temperature between 22 and 26◦C,and light intensity 150-200µE m-2s-1.
At the end of the growth period the samples were collected and prepared for the chemical and SEM-EDS analyses.
2.3 Chemical analysis
To determine the total concentration of the metals and their site of accumulation in the plant,samples were split in shoot and root,oven dried at 70◦C and pulverized in a miller(Retsch S 100)equipped with an agate pocket.An aliquot of 250 mg of each sample were digested with HF(50%)and HNO3(65%)at a ratio of 1:2,in a microwave oven(Milestone mls 1200-Microwave Laboratory Systems).To avoid the risk of contamination,blank samples(mineralization solutions without plant samples)were also analysed.Elemental concentrations were measured by AAS(SpectrAA-Varian)with graphite furnace and calculated considering the values of the blanks;the measurements were performed for each sample in triplicate;in addition,the mean value and standard error were calculated.A standard reference material(CTA-OTL 1,tobacco leaves)was analysed in parallel and used to calculate recover percentages and for analytical quality control.The content of Fe,K,Mg,Mn and Zn in the standard reference material showed recovery percentages in the range 86-109%,indicating a good accuracy of the chemical analysis.
2.4 SEM-EDS analysis
For SEM-EDS analysis,small pieces(1-3 mm)of leaves and roots of both control and Mn-treated plants of Cistus salvifolius were fixed in glutaraldehyde at 2%in phosphate buffer 65 mM at pH 7.2 for 2 hours.Samples were then dehydrated in 10 ml of alcohol at increasing concentrations(30,50,70,90 and 100%)for a time of 15 min for each concentration except 100%of alcohol left to act for 1 hour.After dehydration,the samples were dried in a stove at 40◦C for 1 hour,mounted on aluminum stubs and coated with a thin carbon layer to make conductive their surface.Samples were observed under a scanning electron microscope(SEM;JEOL JSM 5310).The samples were also analyzed by energy-dispersive X-ray spectroscopy(EDX;Oxford INCA).
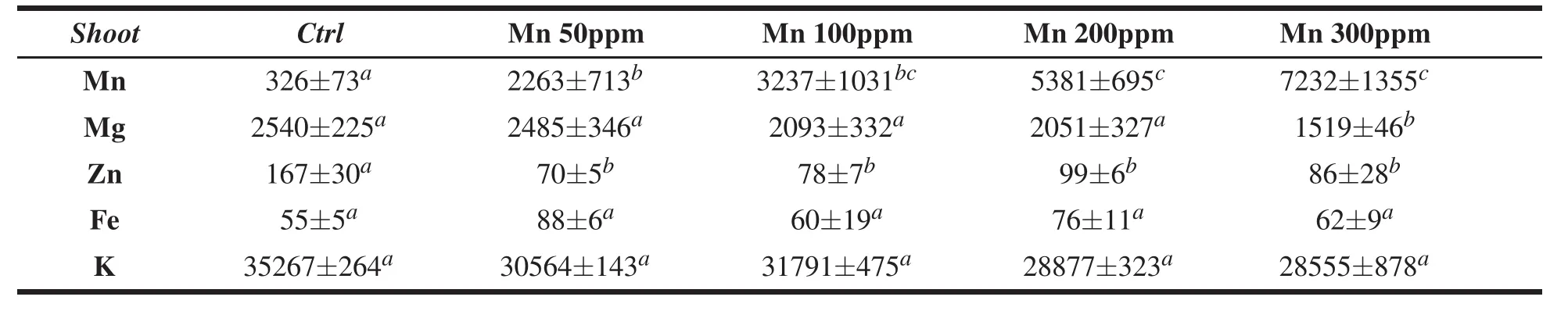
Table 1 Element concentrations(µg/g d.w.)in the shoots(mean± SE,n=3)of C.salvifolius.Different letters indicate significant differences(P<0.05).

Table 2 Element concentrations(µg/g d.w.)in the roots(mean± SE,n=3)of C.salvifolius.Different letters indicate significant differences(P<0.05).
2.5 Chemical data analysis and SEM-EDS acquisitions
To provide an adequate collection of data three samples(n=3)for each Mn treatment were analyzed by AAS,and three measurements were collected for each sample.We performed a one-way ANOVA test to evaluate the statistical significance(for P<0.05)of the differences recorded in the chemical analyses.The ANOVA tests were followed by the post hoc tests of Holm-Sidak.
SEM-EDS was performed in control samples and in both 50 Mn and 300 ppm Mn treated plants.Image and spectrum acquisitions were made in three to five leaf and root-samples for each treatment.For leaves,lower and upper epidermis,palisade and spongy parenchyma were observed and analyzed;for roots,epidermis,cortical cylinder and vascular cylinder were similarly observed and analyzed by SEM-EDS.
3 Results
3.1 Chemical analysis
Element contents(mean values and standard error)adjusted according to each recovery percentage,in the leaves and roots of C.salvifolius grown in the presence of different concentrations of Mn(0-300 ppm)are reported in Tables 1 and 2.
As expected,both in leaves and in roots,at increasing concentration of manganese in the culture medium,a parallel increase in Mn content was observed in plant tissues(Tables 1,2),with a distribution between shoot and root overall similar to control,which indicates that Mn translocation was not affected by its concentration in the liquid medium.
For the other investigated elements,in the roots,the uptake of Mg was enhanced by low Mn concentrations and did not undergo significant variations at high Mn concentrations.In the leaves,at increasing concentration of Mn,the Mg content decreased compared to control;this decrease was significant in the treatment with the highest Mnconcentration(300 ppm).A similar trend was observed for K;in fact,in the roots,a synergistic effect was observed only with low Mn supply,whereas,in the leaves,K content significantly decreased at increasing Mn concentrations(Table 2).In brief,Mn seemed to inhibit both Mg and K uptake in the leaves.
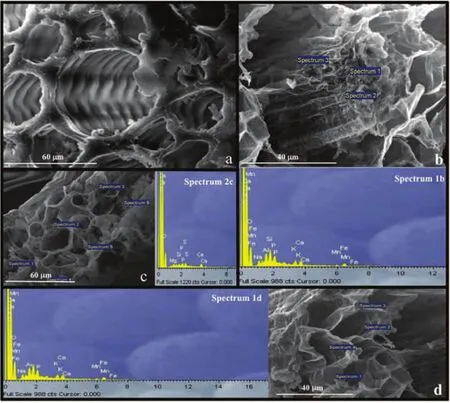
Fig.1 SEM-EDS analysis in C.salvifolius.a)control plant,vascular cylinder of the root;b)300 ppm Mn-treated plant:vascular cylinder of the root and relative spectrum(1b);c)300 ppm Mn-treated plant:leaf and relative spectrum(2c);d)300 ppm Mn-treated plant:root cortex and relative spectrum(1d).In all spectra measure unit of the X axis is KeV.
Theuptake ofZnsignificantly decreased in theroots and leaves inMn-treated plants compared tocontrol.On the contrary,an increasing trend was observed for Fe,both in roots and leaves of Mn-treated plants,particularly evident in the shoot grown with low supply of Mn(Tables 1 and 2).
3.2SEM-EDS
SEM-EDS analyses were performed on leaves and roots of C.salvifolius plants to investigate whether the examined elements were preferentially accumulated in specific tissues.Microanalysis of roots grown in control soil revealed the presence of nutrients in the analyzed tissues;Ca,Na,Mg and K are the most commonly found nutrients,likely deriving from the culture medium.The presence of Al in the spectra depended on the stubs on which the plant samples were mounted(Fig.1).
The addition of 50 ppm Mn did not determine changes in microanalysis spectra compared to the control;in fact,according to a semi-quantitative analysis in both control and treated roots,Mn was always present at mean concentrations between 0 and 0.002%.
With the addition of 300 ppm Mn,it increased in the roots,especially in the cortical cylinder.Mean concentrations of 0.5%and 0.7%were recorded in the central and cortical cylinders,respectively(Fig.1b and 1d;spectra 1b and 1d).Compared to control,Mn treated roots showed a reduced development of the central cylinder,particularly at the highest concentration;at 300 ppm Mn,the diameter of central cylinder was halved(from about 60µm to about 30µm)and the conducting cells appeared much narrower(Fig.1a and 1b).
As for leaves,no significant Mn concentrations were appreciated under SEM-EDS,either with 50 or with 300 ppm of element(Fig.1c,spectrum 1c).The average percentage concentrations were in the order of 0.01%.In addition,all leaves appeared scarcely differentiated,probably due to their early age.Being the treated plants smaller than the control ones,their leaves had a poorly differentiated mesophyll,without distinction between the palisade and the spongy parenchyma,probably because the plants reflect a slowdown in the development.
4 Discussion
Our results on the effect s of Mn on the uptake of mineral nutrients in C.salvifolius highlighted a similar increasing trend for Mg and K and an opposite,decreasing trend for Fe and Zn.The quite similar behavior of Mg and K uptake in response to increasing Mn concentrations well agrees with some effects induced by both nutrients.For example,it is well assessed that magnesium and potassium concentrations in the leaves affect photosynthesis efficiency,and a visible symptom of their deficiency is leaf chlorosis and necrosis,as observed in Pinus radiata D.Don,Morus alba L.and Gossypium hirsutum L.(Sun and Payn,1999;Zhao et al.2001;Tewari et al.2006);this result is coherent with the evident necrotic areas observed especially in older leaves of C.salvifolius at the end of Mn treatment(Fig.2).Moreover,both Mg and K deficiency are known to cause an alteration of carbohydrate partitioning between root and shoot(Cakmak et al.,1994;Zhao et al.,2001);further analyses are needed to confirm this effect also in C.salvifolius.Many micronutrients,including Fe and Zn,act as constituents of prosthetic groups and catalyze redox processes by electron transfer(Romheld and Marschner,1991).The opposite trend observed in the uptake of Fe and Zn in C.salvifolius could be interpreted as a compensative mechanism adopted under Mn stress to balance the two metal co-factors and avoiding a misfunctioning of redox processes.The interaction of heavy metals with the uptake and transport of macro and micronutrients was highlighted in many studies(Ebbs and Kochian,1997;Wenzel and Jockwer,1999;Liu et al.,2000).Manganese alters the supply of minerals,which often act as co-factors in different enzymatic pathways;this may result in a stunted growth and alterations even at the ultrastructural level,depending on Mn concentrations(Subrahmanyam and Rathore,2000;Fecht-Christoffers et al.,2003).Although synergistic and antagonistic effects did not follow a dose-dependent univocal trend,phenomena of nutritional antagonism generally involve the ions of the same charge.Chemical analysis confirms an increase in the transport of Mn at increasing concentrations and its in fluence on the absorption of other essential nutrients,as already reported in the literature(Shi et al.,2006).The visible state of stress that is observed in plants at high Mn concentrations with chlorotic and necrotic areas may,therefore,be due to such variation and not only to Mn concentrations(Fig.2).
SEM-EDS analyses were aimed to investigate the accumulation of Mn in the different plant structures and to relate Mn accumulation to eventual morpho-anatomical changes.Our results indicated a general slowdown of plant development under Mn stress,also evident in the leaves,poorly differentiated,despite Mn was not detectable in leaf tissues.The severe reduction of the vascular cylinder and its conducting cells can be interpreted as a morphological response to high concentrations of Mn and it is in line with previous observations reported by Millaleo et al.(2010),showing that high Mn concentrations inhibit growth.At the same age,Mn-treated plants grew less and showed less developed and thinner roots.Of course,it cannot be excluded that the morphological changes observed by SEM under Mn stress could be related to the altered mineral nutrition induced by Mn.

Fig.2 C.salvifolius 300 ppm Mn-treated plant at the end of the growth period(15 d).
5 Conclusion
In the present study the growth performance of the metallophyte C.salvifolius was evaluated with respect to its tolerance to increasing concentrations of bioavailable Mn in hydroponic culture;in addition,the interference of Mn with the uptake of micro and macronutrients(Fe,Zn,Mg and K)was screened as well.The accumulation of the investigated nutrients did not follow a dose-dependent univocal tendency.The similar trend observed for Mg and K uptake under Mn stress can be related to their common role in photosynthetic metabolism and the presence of necrosis at the end of the culture period strengthen this hypothesis.The opposite trend observed for Zn and Fe uptake could be a plant response to keep efficient redox processes under Mn stress.SEM-EDS highlighted that Mn stress determines a histological immaturity in leaves and a reduced development of vascular cylinder in root,which,based on semi-quantitative analysis,accumulated most of the supplied Mn.In future experiments other nutrients and growth/vitality parameters could be studied to reach a complete view on the response of C.salvifolius under metal stress.Plant mineral nutrition represents indeed,one of the key factors affecting metal tolerance and biomass production;therefore,these studies can provide useful knowledge to plan and promote recovery actions of metal polluted soils using native flora.
 Journal of Environmental Accounting and Management2018年4期
Journal of Environmental Accounting and Management2018年4期
- Journal of Environmental Accounting and Management的其它文章
- Light Fertilization Affects Growth and Photosynthesis in Mung Bean(Vigna radiata)Plants
- Capture Rate of Selected Heavy Metals In Q.Ilex L.Leaves Collected At Two Sites With Different Land Uses
- Impact of Biochar Amendment on Soil Quality and Crop Yield in a Greenhouse Environment
- Regional Redistribution Effects of Renewable Energy Subsidies
- Phytotoxic Extracts as Possible Additive in Subsurface Irrigation Drip for Organic Agriculture
- Ecotoxicological Assessment of Virgin Plastic Pellet Leachates in Freshwater Matrices