
红苞凤梨嵌合体组织细胞学观察与叶绿素合成特性研究
2019-01-09 01:44刘和平杨转英张俊丽林剑波何业华
广东农业科学 2018年11期
刘和平,曹 莉,杨转英,张俊丽,林剑波 ,何业华
(1.阳江职业技术学院食品与环境工程系,广东 阳江 529566;2.华南农业大学园艺学院,广东 广州 510642;3.广东海洋大学农学院,广东 湛江 524088;4.茂名市水果科学研究所,广东 茂名 525000)
叶绿素是植物进行光能吸收与转化的主要色素,其合成途径主要为:Glu(Glutamate,谷氨酸) →ALA(δ-Aminolevulinic acid,δ-氨基酮戊)→PBG(Porphobilinogen,胆色素原)→UroⅢ(Uroorphyrinogen Ⅲ,尿卟啉原 Ⅲ)→ProtoⅨ(Protoporphyrin Ⅸ,原卟啉 Ⅸ) →Mg-ProtoⅨ(Mg-Protoporphyrin Ⅸ,镁原卟啉Ⅸ)→Pchl原脱植基叶绿素Pch(Protochlorophyll,原叶绿素酸)→Chl a(Chlorophylla,叶绿素a)→Chl b(Chlorophyll b, 叶绿素 b),以上任何一个步骤受阻都会影响叶绿素正常的生物合成,随之引起一系列的生长表型变化[1-2]。冬小麦[1]、水稻[3-4]、也门铁[6]、红苞凤梨[7]等多种植物具有叶绿素合成前体物质发生变化导致叶绿素合成受阻而表现出花叶等嵌合现象。
红苞凤梨(Ananas bracteatus)又名艳凤梨、斑叶凤梨,为凤梨科凤梨属观赏植物,因其冠芽和叶片白、绿相间的嵌合特点,颇具观赏价值,为国际上重要的盆栽花卉和鲜切花[8-9]。红苞凤梨等花叶植物不仅具有观赏价值,也是研究叶色嵌合形成机理的重要材料,在观赏植物育种和开发利用中占重要地位[10-11]。近年来,曹莉等[8,12-16]分别从红苞凤梨嵌合性状变异规律和组织细胞学、转录组信息、叶绿素前体物质及叶片白化关键基因筛选、内参基因筛选、胚性悬浮系及转基因等方面开展研究,并取得一定成果,但鲜见结合组织细胞学与叶绿素合成两个方面研究红苞凤梨嵌合形成特性的报道。
本研究通过对红苞凤梨嵌合体进行组织细胞学观察,结合分析叶绿素及其合成前体物质含量变化,初步确定红苞凤梨嵌合叶片的组织细胞学特性及其叶绿素合成受阻位点,阐述其嵌合性状形成的组织细胞学基础及叶绿素合成生理因素,为揭示红苞凤梨嵌合性状形成机理、提高红苞凤梨嵌合体繁殖的稳定性提供理论基础,为植物花叶嵌合性状理论研究提供参考。
1 材料与方法
1.1 试验材料
供试材料为红苞凤梨金边、金心、全白与全绿4个品种,来源于华南农业大学园艺科研基地。其中红苞凤梨金边、金心及全绿植株均为生长良好、长势一致的2年生盆栽植株,全白材料为嵌合体组培苗全白突变芽发育而成。
1.2 试验方法
1.2.1 切片 选取红苞凤梨金边和全绿植株幼嫩叶片,截取较宽部位2~3 cm置于载玻片上,用两片新双面刀片贴紧垂直向下横向切取获得15~20 μm薄片,将其展于清水中,做成简易切片并在显微镜下观察。
1.2.2 叶绿素及其合成前体物质含量测定 分别剪取红苞凤梨各品种植株叶片的绿色和白色部分测定叶绿素及其合成前体物质含量。设全绿株叶片为对照,并设定其叶绿素及合成前体物质含量为100%。3次重复,每个重复取自3棵不同植株。
(1)ALA、PBG、UroⅢ含量测定。参照徐培洲等[3]方法并稍作改进。
ALA测定:取0.5 g叶片,用4%三氯乙酸(15 mL)及少量石英砂研磨,合并上清液并定容至20 mL,18 000 r/min离心15 min,取5 mL上清液,加入2.35 mL 1 mol/L醋酸钠和0.15 mL乙酰丙酮,沸水浴10 min后冷却至室温。取2 mL加入显色液(Ehrlich-Hg 试剂:1 g P- 二甲氨基苯甲醛溶解于30 mL冰醋酸中,加入8 mL 70%高氯酸,冰醋酸稀释到50 mL,最后加入0.2 g 氯化汞)黑暗下显色15 min,然后测定OD553值。
PBG 含量测定:0.5 g叶片用液氮研磨后,加入5 mL提取缓冲液(0.6 mol/L Tris,0.1 mol/L EDTA,用盐酸调节 pH至8.2),过滤或18 000 r/min离心 10 min。加入2 mL Ehrlich-Hg 试剂,黑暗中显色15 min,测定OD553值。
UroⅢ含量测定:1 g叶片用液氮研磨后,加入10 mL 提取缓冲液(0.067 mol/L磷酸缓冲液,pH 6.8),过滤或18 000 r/min 离心10 min。取5 mL加入0.25 mL 1%Na2S2O3,剧烈振荡后强光照射20 min。加入1 mol/L 甲酸(或冰醋酸)至pH 值3.5。用10 mL乙醚萃取3次,分层后测定水相的OD405.5值。
(2)ProtoⅨ、Mg- ProtoⅨ、Pchl含量测定。采用Hodgins等[17]方法。取0.3 g去主脉鲜叶,加25 mL 80%碱性丙酮,研磨提取后,分别在波长575、590、628 nm处测得OD值,计算其含量。
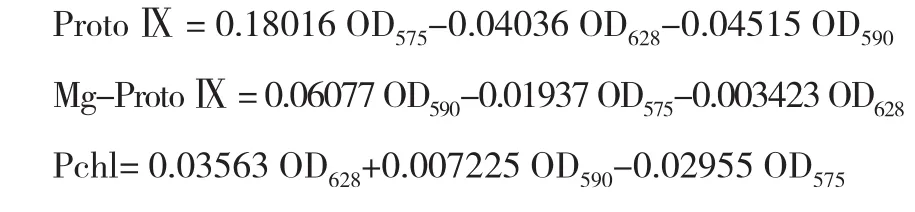
(3)叶绿素(Chl)含量测定。取新鲜叶片0.5 g,置于2 mL丙酮∶乙酸=1∶1中浸泡24 h,浸提液在663、645 nm处测OD值,计算叶绿素的浓度和含量。
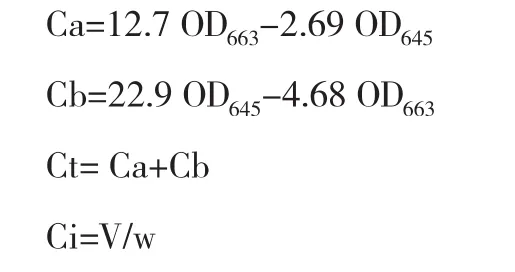
式中,Ca为叶绿素a浓度(mg/L),Cb为叶绿素b浓度(mg/L),Ct为叶绿素总浓度(mg/L),V为浸提液体积(mL),w为材料质量(g)。
2 结果与分析
2.1 嵌合体植株叶片的组织细胞学观察
红苞凤梨叶片切片如图1~图3(封三)所示。图1(封三)是红苞凤梨金边嵌合体叶片红色物质较少时(图4,封三)的切片,图2(封三)是该植株红色物质较多时(图4B,封三)的叶片切片,图3(封三)是全绿植株叶片切片。嵌合体植株叶片细胞表现出含叶绿体、不含叶绿体和含红色物质等3种类型。组织细胞学观察表明,红苞凤梨金边嵌合体植株叶片白色部分(即金边)的细胞中几乎不含叶绿体(图1A、图1B、图2A,封三)。红苞凤梨金边嵌合体植株叶片中部的海绵细胞含有丰富的叶绿体(图1C、图1D、图2A,封三)。叶片中部上表皮下有两层栅栏细胞,栅栏细胞都几乎不含叶绿体。其中上层栅栏细胞排列紧密,形状稍短;下层栅栏细胞具典型形态,形状稍长。从图1E~F(封三)可以观察到,该状态下红苞凤梨金边嵌合体植株叶片白色部分的上下表皮中分布少量的含红色物质细胞。从图2B~D可以看出,该状态下红苞凤梨嵌合植株的整个叶片上下表皮中都分布了丰富的红色物质细胞。从图3(封三)可以看出,红苞凤梨全绿植株整个叶片含有丰富的叶绿体,但基本无红色细胞。全绿植株叶片中部上表皮下有2层栅栏组织,但第一层栅栏组织细胞基本不含叶绿体(图3C,封三)。
红苞凤梨金边嵌合体植株叶片切片结果与田间观察嵌合表型相一致。金边嵌合体植株叶片白色部分几乎不含叶绿体(图1、图2,封三),田间观察即表现为白色;嵌合体植株叶片中部含有丰富的叶绿体(图1、图2,封三),田间观察即表现为绿色。全绿植株叶片含有丰富的叶绿体,田间观察即全叶呈现绿色,即红苞凤梨嵌合体植株叶片的绿色部分主要是该部位细胞含有叶绿体所致,白色部分主要是该部位细胞几乎不含有叶绿体所致。
2.2 红苞凤梨各品种叶绿素及其合成前体物质含量
由图5可知,红苞凤梨金边嵌合体植株叶片绿色和白色部位的叶绿素含量均低于对照,田间观察也表明叶片绿色部位的颜色比全绿株叶片要浅。红苞凤梨金边嵌合体植株叶片绿色部位的叶绿素合成前体物质含量在Pchl合成前一直高于对照,且ALA、PBG含量显著高于对照;而白色部位仅ALA和PBG含量高于对照,其中ALA含量约为对照的2.5倍,但从UroⅢ开始叶绿素合成前体物质含量就迅速下降,且显著低于对照。
从图6可以看出,红苞凤梨金心嵌合体植株叶片绿色和白色部位的叶绿素含量都低于对照,其中白色部位叶绿素含量很低,绿色部位含量显著高于白色部位,与田间表型一致。红苞凤梨金心嵌合体植株叶片白色部位的ALA和PBG含量高于绿色部位,但从UroⅢ开始,绿色部位的叶绿素及其合成前体物质含量呈上升趋势,且都显著高于白色部位。红苞凤梨金心嵌合体植株叶片绿色和白色部位的叶绿素合成前体物质PBG含量高于对照,此后其前体物质都低于对照。其中白色部位的UroⅢ含量不到对照的1/10,ProtoⅠ、Mg-protoⅨ和Pchl含量接近对照的1/5。
从图7可以看出,红苞凤梨全白植株叶片的叶绿素及其合成前体物质含量一直都显著低于对照。
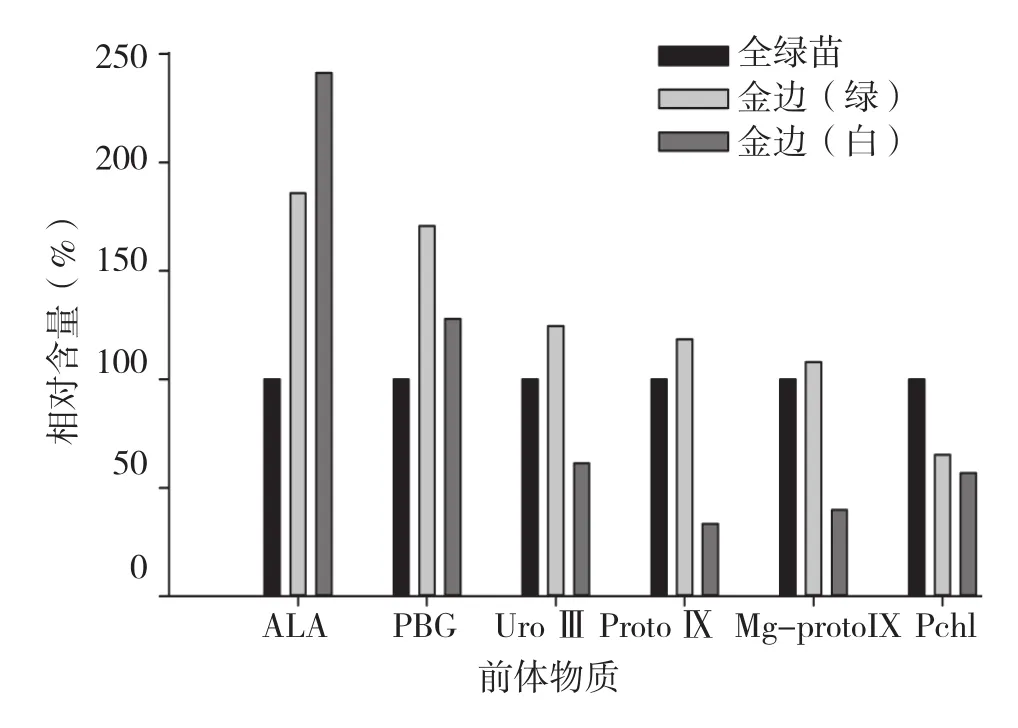
图5 红苞凤梨金边嵌合体叶绿素及前体物质含量
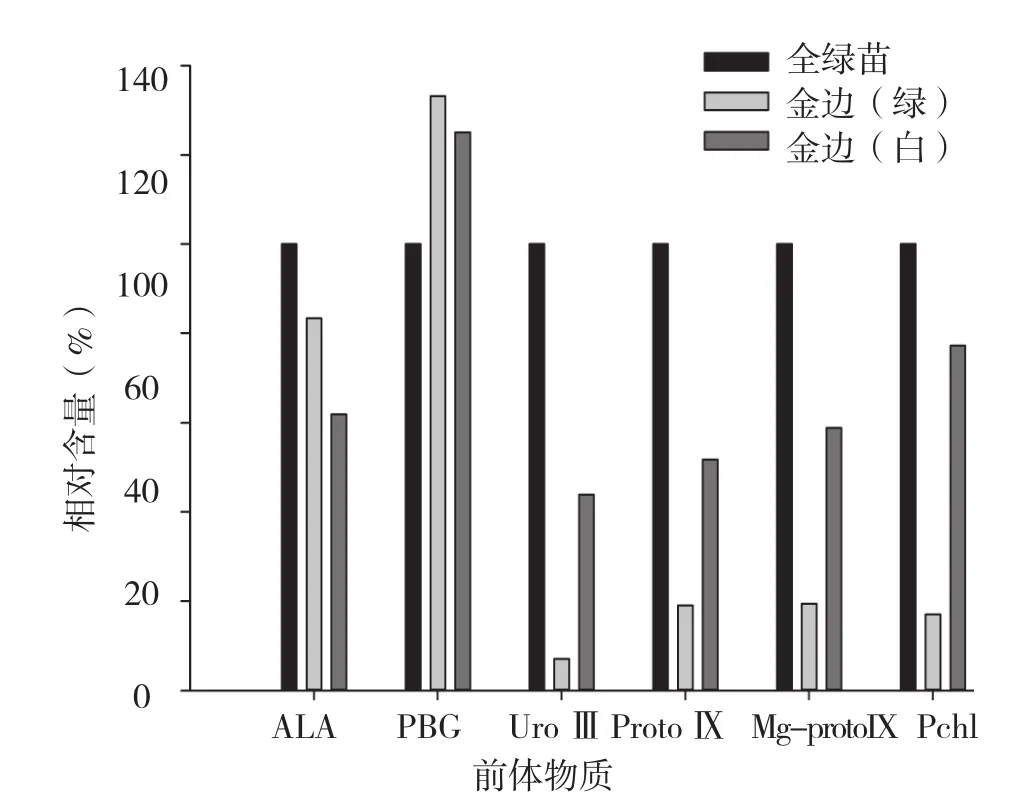
图6 红苞凤梨金心嵌合体叶绿素及前体物质含量
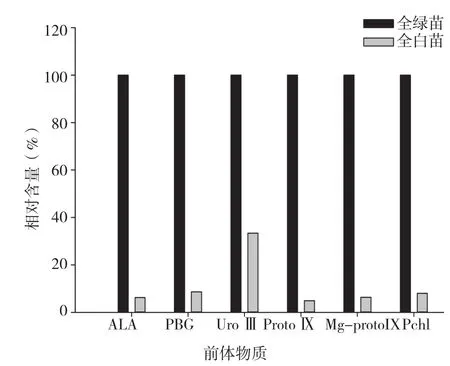
图7 红苞凤梨全白株叶绿素及前体物质含量
2.3 红苞凤梨各品种叶绿素含量比较结果
由图5~ 图7可知,红苞凤梨金边、金心、全白植株叶片的各部位都含有叶绿素,但含量丰富程度不同。以对照叶绿素含量为100%,各品种叶片叶绿素相对含量表现为全绿株>金心绿色部位>金边绿色部位>金边白色部位>金心白色部位>全白株,可见,叶绿素含量与田间观察结果相一致。
3 结论与讨论
叶绿素生物合成受阻会导致叶绿素合成量减少,从而引起黄化甚至白化。本研究结果表明,红苞凤梨全绿、金边、金心、全白植株叶片各部位的叶绿素含量丰富程度不同,各品种叶绿素相对含量表现为全绿株>金心绿色部位>金边绿色部位>金边白色部位>金心白色部位>全白株,叶绿素含量与植株叶片田间表型对应。结合切片结果可以确定,植株叶片绿色部分细胞含有丰富的叶绿素,而白色部分细胞叶绿体含量少是红苞凤梨叶片嵌合性状形成的主要原因,这与也门铁研究结果[11]相一致。
红苞凤梨金边嵌合体植株叶片绿色部位的叶绿素合成前体物质含量在Pchl合成之前一直比全绿苗(对照)高;白色部位的ALA和PBG含量高于对照,但从UroⅢ开始叶绿素合成前体物质含量迅速下降,且显著低于对照。初步确定红苞凤梨金边嵌合体植株叶片白色部位的叶绿素合成在UroⅢ合成时受阻,导致叶绿素合成代谢下降。
红苞凤梨金心嵌合体植株叶片绿色部位和白色部位的叶绿素合成前体物质PBG含量高于对照,从UroⅢ开始合成前体物质都低于对照,其中白色部位的UroⅢ含量不到对照的1/10,ProtoⅨ、Mg-protoⅨ和Pchl含量仅接近对照的1/5。初步确定红苞凤梨金心嵌合体植株叶片白色部位的叶绿素合成也是在UroⅢ合成时受阻,导致叶绿素合成代谢下降。该研究结果与水稻[5]、金边也门铁[6]等植物一致。
红苞凤梨全白植株叶绿素及合成前体物质含量均显著低于对照,说明红苞凤梨白化植株的叶绿素合成是在前体物质ALA合成时受阻,导致叶绿素合成代谢下降,这与冬小麦等研究结果[1]一致。
红苞凤梨各品种叶绿素合成受阻,表明在叶绿素合成途径中存在某个调控因子,导致叶绿素合成代谢下降,叶绿素含量低的部位出现白化现象,最终导致红苞凤梨金边、金心等品种植株叶片表现出条斑等花叶嵌合性状。但调控机理还需进一步深入研究。
猜你喜欢
科学与社会(2021年3期)2021-12-02
幸福·婚姻版(2020年7期)2020-09-02
环球人物(2020年13期)2020-07-13
孩子(2020年4期)2020-06-08
意林(2020年9期)2020-06-01
大自然探索(2020年3期)2020-05-03
短篇小说(原创版)(2020年12期)2020-03-24
短篇小说(2020年12期)2020-03-11
中国-东盟博览(旅游版)(2018年10期)2018-05-14
发明与创新·中学生(2017年6期)2017-06-12
