
Pb胁迫对水稻繁殖策略的影响
2019-01-02 03:36:26陈嘉伟黄一格张居奎王立宁
东北师大学报(自然科学版) 2018年4期
王 颖,陈嘉伟,黄一格,张居奎,王立宁
(1.吉林建筑大学市政与环境工程学院,吉林 长春 130118;2.吉林省第五地质调查所,吉林 长春 130061)
重金属污染近年来已成为全球影响环境和威胁人类健康最重要的环境问题之一[1].土壤重金属积累破坏土壤生态系统的同时,也导致重金属在植物中大量积累,严重影响植物的生长发育,而且重金属经食物链可危及动物和人类健康[2].进入土壤中的重金属严重影响土壤的理化性质,破坏植物的生存环境,使植物生长、发育及生物量在不同器官的分配、种子特性等发生变化,最终导致植物繁殖策略发生改变;同时植物也会采取一定的生长、繁殖策略适应生态环境以增加其生存的适合度[3].因此,研究重金属污染对植物繁殖策略的影响有助于理解植物种群发展,也具有一定的应用价值.
植物种群的更新、个体生活史的完成依赖于植物的繁殖过程.不同环境下,植物通过改变繁殖策略格局协调生存、生长与繁殖间的协同进化关系,以更好地适应环境变化[4].每一种植物都有其特定的繁殖策略,同时环境变化对植物繁殖策略也具有重要影响.环境的差异会导致植物不同器官间的生长、发育存在一定的权衡关系[5],即控制资源在不同组织或器官间进行有效分配达到生长和繁殖的平衡,这种平衡关系反映了繁殖部分与非繁殖部分生物量或能量的比例关系[6].繁殖分配在很大程度上决定植物获取资源和适应环境的能力,也是决定种子特性的一个重要因素,种子特性包括种子产量、大小、数量等[7].
铅(Pb)是土壤环境最主要的重金属污染物.王永强等[8](2010)报道了高浓度Pb胁迫可显著降低水稻(Oryzasativa)穗数、结实率、千粒重、每穗总粒数和产量,低浓度Pb胁迫则使每穗总粒数增加,而且水稻籽粒中Pb含量与土壤中Pb浓度呈显著正相关.Pb胁迫还导致水稻成熟期推迟、空秕率增加,从而使产量大幅度下降[9].水稻是世界上一半以上人口的粮食作物,更是与我国人民生活息息相关的最主要的粮食作物之一.因此,以水稻为研究对象探讨Pb污染胁迫对其生长、发育的影响,不仅可为提高重金属污染地区农产品的产量和质量提供参考,而且可为环境变化对植物繁殖策略的影响研究提供一定的理论支持.
1 材料与方法
1.1 实验材料
供试水稻品种为吉农985,水稻秧苗来自吉林农业大学农学院水稻育苗基地.用于培育幼苗的土壤环境和水稻后期生长的土壤环境基本一致.
1.2 实验设计
实验在吉林建筑大学实验基地进行,用防雨篷控制降水.2016年4月1日开始,采用桶培方式进行实验,所用塑料桶直径25 cm、高20 cm.将取自农田表层0~30 cm土层的土壤碾碎、风干,过尼龙筛去除杂物、根系等,每桶装8 kg备用土.实验设置Pb(以纯Pb计,由Pb(NO3)2提供)胁迫处理的质量分数分别为:0,500,1 500,2 500 mg/kg.将每个处理的药品分别稀释在1 L水中,与事先称好的8 kg备用土壤充分混匀,每个处理4次重复.所有塑料桶盖上遮光布放置在防雨棚内1个月,5月1日秧苗约30 cm时移栽到塑料桶中.每桶保留6株秧苗进行胁迫处理,16个塑料桶随机排放在防雨篷内.水稻生长过程中,每5 d浇1次水,每次浇水1 L,并进行正常管理.
1.3 取样和数据收集
在水稻成熟期,计数每桶内每株总分蘖数和繁殖分蘖数.然后用剪刀将植株地上部分齐土面剪下,地上部分分为繁殖蘖和非繁殖蘖,繁殖蘖分为繁殖蘖营养部分、种子部分和繁殖支撑物部分(穗去掉种子部分),非繁殖蘖和繁殖蘖营养部分统称非繁殖部分.计数每株水稻的种子数,然后将每个塑料桶的所有种子混合,随机取50粒,在60℃烘箱中烘干至恒重,测定并计算单粒重.分别将植株各部分放入烘箱中烘干至恒重,并测量各部分干重.最后将每个花盆所有植株相同部分混合,烘干、粉碎,干样品密封保存,待测各部分重金属含量.采用ICP-MS法测定种子中的Pb含量.
1.4 数据分析
所有数据用SPSS17.0(version 17.0,SPSS Inc.,Chicago IL,USA)统计软件进行分析,对分蘖数、地上部分和种子生物量、生物量分配、单粒重及种子中Pb含量等进行单因素方差分析,统计分析中显著性检验均采用LSD检验,显著水平为P=0.05.
为了分析不同Pb胁迫下种子数量与植株高度、单粒重之间的异速生长关系,将收集的数据对数转换成lg(y)=lg(b)+alg(x).其中:b代表易速关系的截距;a为易速关系的斜率,当a=1时为等速生长,当a≠1时为异速生长.用标准主轴回归方程确定斜率和截距,用SMART(Standardized Major Axis Tests and Routines)软件分析并检测斜率间差异显著性.
2 结果与分析
不同质量分数Pb胁迫处理对水稻总分蘖数和繁殖分蘖数均具有显著影响(见表1).Pb质量分数为0~1 500 mg/kg时,总分蘖数无显著变化(P>0.05);Pb质量分数为2 500 mg/kg时,总分蘖数显著下降到最小值(P<0.05).繁殖分蘖数与总分蘖数的变化规律基本相同,高质量分数Pb胁迫条件下繁殖分蘖数显著下降.

表1 Pb胁迫对水稻分蘖数的影响
注:同列内相同字母表示P=0.05水平无差异.
Pb胁迫显著影响水稻地上部分、种子生物量及各部分生物量分配比率(见图1).0~1 500 mg/kg Pb胁迫处理地上部分植株生物量无显著变化(P>0.05),2 500 mg/kg Pb胁迫处理地上部分植株生物量显著下降到最小值(P<0.05);种子生物量与地上部分植株生物量具有相同的变化规律(见图1A).
水稻种子生物量分配比率在500 mg/kg Pb胁迫处理下达到最小,非繁殖部分生物量分配比率变化趋势与种子生物量分配比率具有相反的变化规律,在500 mg/kg Pb胁迫处理下达到最大;繁殖支撑物生物量分配比率在不同强度Pb胁迫处理下无明显变化(见图1B).
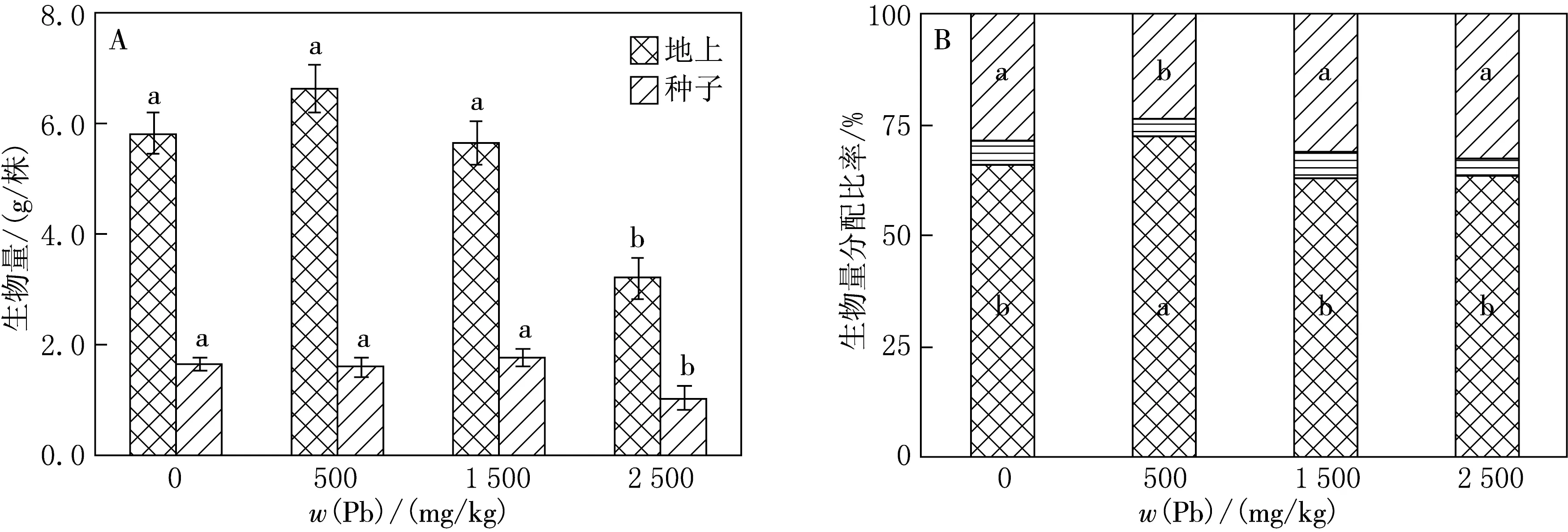
图1 Pb胁迫对水稻地上部分和种子生物量(A),种子、繁殖支撑物及非繁殖部分生物量分配比率(B)的影响
不同质量分数Pb胁迫处理下种子数量与植株高度均存在异速生长关系,种子数量与植株高度异速生长的斜率无显著变化(P>0.05),且斜率均显著大于1(见表2、图2A).
Pb胁迫处理下水稻单粒重与种子数量无异速生长关系,但对照条件下存在异速生长关系(见表3、图2B),单粒重与种子数量对照条件下异速生长关系斜率显著小于-1.

表2 Pb胁迫对水稻种子数量与植株高度间异速生长关系的影响
注:同行内相同字母表示在P=0.05水平无显著差异.
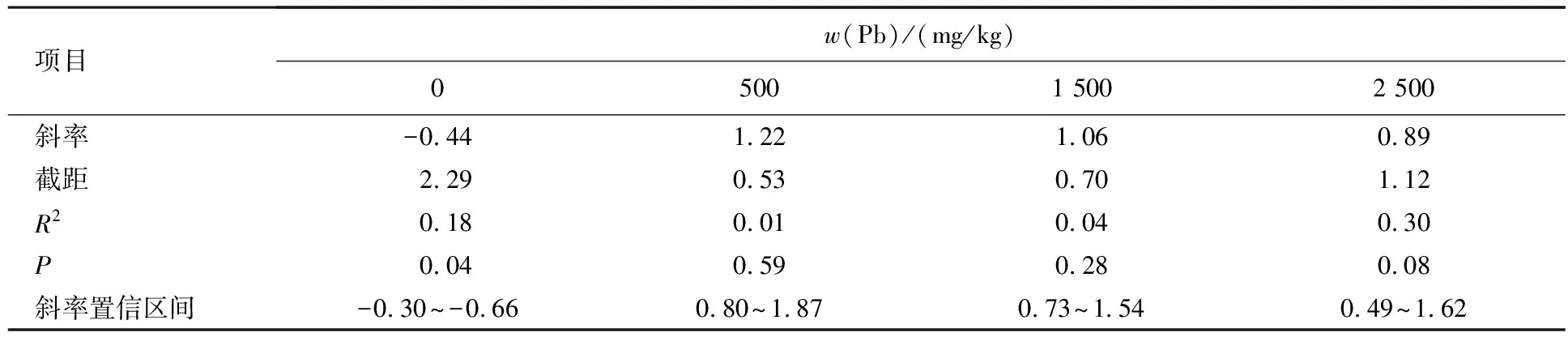
表3 Pb胁迫对水稻种子数量与单粒重间异速生长关系的影响

图图2Pb胁迫对水稻种子数量与植株高度(A)、单粒重(B)异速生长关系的影响
不同质量分数的Pb胁迫显著影响水稻单粒重和种子内的Pb含量(见图3).单粒重在0~1 500 mg/kg Pb胁迫处理条件下无显著变化(P>0.05),在2 500 mg/kg Pb胁迫处理时,单粒重下降到最小值(见图3A);种子内Pb含量随Pb胁迫强度的增加而显著增加(P<0.05),2 500 mg/kg Pb胁迫处理种子内Pb含量达到最大值(见图3B).

图3 Pb胁迫对水稻单粒重(A)和种子内Pb含量(B)的影响
3 讨论
植物为了适应重金属污染的环境条件会采取一定的适应策略,例如,在繁殖过程中调整分蘖数、繁殖分配比例,改变种子特性等.本文的研究表明,Pb胁迫对水稻分蘖数具有显著影响,在高质量分数Pb胁迫下分蘖数显著下降;0~1 500 mg/kg Pb胁迫处理时,水稻总分蘖数及繁殖分蘖数无显著变化,说明在此胁迫范围内水稻表现出一定的抗性,这与前人[10]的研究结果一致.分蘖作为水稻自身的生理遗传特性,其数量是水稻高产栽培的重要环节之一[11],因此,对水稻分蘖数的研究至关重要.
在不同的环境压力下,植物的生长、发育过程会采用不同的繁殖分配格局,这种分配格局的变化对于植物能否对其生长的环境达到最大适合度具有重要作用,因此,研究植物在不同环境下的繁殖分配比率对于理解植物的适应机制是非常必要的[12].本研究中,500 mg/kg Pb胁迫处理下繁殖部分分配比率最小,这可能是因为Pb的含量还没有达到产生毒害的阈值,反而对水稻生长起到了促进作用,生物量积累在茎、叶中的比例增加,导致植物非繁殖部分生物量分配比率增大,即在资源适宜的条件下植物会将更多的资源分配到营养生长以增加植株大小提高潜在繁殖能力的机会[13];相反,对照和高质量分数的Pb胁迫处理则抑制了植物生长,提高了生物量分配到繁殖部分的比率,这种资源分配模式是植物在异质环境中的适应性特征,是为了最大可能地保留遗传多样性、生产相对较多的种子.有研究[14]表明,高浓度重金属胁迫环境对水稻生长将产生不同程度的伤害,主要表现为光合速率下降、同化物形成减少、叶绿体受伤,呼吸速率也会发生变化,有关光合和呼吸的酶失活或变性.水稻体内这种生理、生化变化导致其生长、发育发生变化,进而影响水稻的繁殖分配,分配到繁殖部分的生物量或能量增多.
本研究中,种子数量与植株高度的异速生长关系斜率显著大于1,表明植株越高、产生的种子越多.单粒重与种子数量间的权衡结果显示,在对照条件下单粒重与种子数量间存在异速生长关系,这种关系是植物繁殖投入中的又一个重要权衡.植物种子大小是资源投入权衡的结果,即植物选择生产少而大的种子还是多而小的种子,这种权衡会影响到植物的多个方面,甚至会影响到植物的种间关系和群落结构,是植物对生存环境的一种适应,尤其在资源有限时更容易出现生活史特征的改变[15].
本文的研究结果表明,Pb胁迫显著影响水稻的繁殖策略,特别是高质量分数的Pb胁迫会抑制水稻的生长发育,导致稻米产量和品质显著下降.
猜你喜欢
种子(2023年9期)2023-11-22 13:10:56
麦类作物学报(2021年3期)2021-05-25 07:03:32
今日农业(2020年19期)2020-12-14 14:16:52
北京农学院学报(2020年1期)2020-03-09 07:18:28
农家科技中旬版(2018年2期)2018-05-05 10:09:54
现代农业(2018年7期)2018-02-18 10:58:09
中国科技信息(2015年2期)2015-11-16 08:18:32
新农村(2015年10期)2015-09-28 00:37:29
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
甘肃农业科技(2014年8期)2014-01-16 04:34:24
