
高原花斑裸鲤过氧化氢酶(CAT)基因的克隆鉴定表达及其对Cu2+胁迫的应答※
2018-12-14 08:07王玉晴朱世海栗瑞红卫福磊韩步鹰刘小红千康康简生龙王国杰李长忠
中国高原医学与生物学杂志 2018年2期
王玉晴,吴 君,朱世海,张 霞,栗瑞红,卫福磊,刘 扬,韩步鹰,刘小红,千康康,简生龙,王国杰,李长忠*
(1.青海大学 生态环境工程学院, 青海 西宁 810016;2.青海省渔业环境监测站,青海 西宁 810012)
生物标志物作为敏感的生物效应“早期预警”工具,可以快速、准确地预测生物体在受到重金属污染时的毒性与毒害作用,但如何筛选有效的生物标记物是研究的难点。本研究以高原花斑裸鲤(Gymnocypriseckloni)为研究对象,拟通过对高原花斑裸鲤过氧化氢酶(catalase,CAT)基因的克隆鉴定表达及对其Cu2+胁迫的应答分析,筛选Cu2+胁迫下的有效生物标记物。
2013年发布的中国渔业生态环境状况公报表明,铜污染在重要渔业区中已越来越严重。铜作为超氧化物歧化酶(SOD)等催化酶类的辅助因子,是生物体所必需的微量元素[1],而铜作为必需的微量元素和致毒金属之间的浓度范围是很小的[2-3]。当生物体内铜过量时,机体产生大量的ROS,引起蛋白质、脂类等的氧化以及免疫毒性[4],导致重金属中毒,而SOD-CAT系统作为抗氧化防御的第一道防线,能有效清除ROS,从而使机体有效地抵制氧化反应[5]。有研究表明[6],CAT可以作为一种有效的生物标志物(biomarker)应用于分子生态毒理的研究中。
1 材料与方法
1.1 实验动物
实验前将水族箱用高锰酸钾进行消毒(3~5h),然后用清水洗净。养殖用水经西安国联质量检测技术股份有限公司检测无重金属超标现象,为正常水质。
3龄期成体高原花斑裸鲤取自青海省渔业环境监测站隆务河口养殖场,体重为105±10 g,体长为19±3 cm。运回实验室后,暂养于青海大学生态环境工程学院细胞遗传学实验室冷流水养殖箱(温度17±2℃;光照周期为14h/10h),适应性饲养2周,然后挑取无伤病、规格相近的健壮个体作为实验对象。
1.2 胁迫方法
本研究选择以《中国渔业水质标准》(GB 11607-1989)中Cu2+的标准值(0.05mg/L)为胁迫浓度。试验设置1个空白组和1个处理组,同处理组设置3个平行组。每个平行组投入6尾高原花斑裸鲤。实验期间正常喂食,每天换三分之一的水以保持水体清洁,并增加相应量的Cu2+以维持浓度的平衡。在胁迫后12、24、48 h时取样,每次取样时,随机从每组取1尾鱼,解剖鳃、脑、肌肉、肾脏和肝脏组织,立即用液氮速冻,然后放入-80 ℃冰箱保存。
1.3 试剂
CuSO4和总RNA提取试剂盒(DP419,天根,中国),PrimeScriptTMⅡ1st strand cDNASynthesis Kit(6210A,TaKaRa,Japan),SMARTer RACE 5′/3′Kit(TaKaRa),Premix ExTaq(TaKaRa),pMDTM 19-T Vector Cloning Kit(TaKaRa),PrimeScriptTM RT reagent Kit(Perfect Real Time,RR0370A,TakaRa,Japan),iQTM SYBR Green Supermix(BIO-RAD)
1.4 实验方法
1.4.1CAT基因的克隆
用总RNA提取试剂盒提取高原花斑裸鲤肝组织总RNA,用PrimeScriptTMⅡ1st strand cDNA Synthesis Kit及SMARTer RACE 5′/3′Kit(TaKaRa)试剂盒获得cDNA模板。
根据虹鳟鱼(Oncorhynchusmykiss)、鲤鱼(Cyprinuscarpio)、斑马鱼(Daniorerio)、鲫鱼(Carassiusauratus)等已知物种的CAT基因cDNA保守序列,设计简并引物(表1)扩增特异性片段。用特异性片段设计RACE引物(表1),通过RACE技术获得基因末端片段。将得到的三段序列拼接并再次设计引物(表1),应用RT-PCR法扩增并测序,确定克隆得到的是GeCATcDNA全长序列。所有测序均由生工生物工程(上海)股份有限公司完成。
表1PCR引物相关参数

Table 1 Reference of the Oligo nucleotide primers used for PCR
1.4.2 序列分析与同源性分析
首先利用National Centre for Biotechnology Information blastn(https://blast.ncbi.nlm.nih.gov/Blast.cgi)比对序列,通过DNAMAN V6进行蛋白质翻译,利用NCBI blastp中保守结构域数据库(Conserved Domain Database,CDD)分析保守结构域,利用VectorNTI11、MAGA5对GeCAT进行同源性分析和系统进化树的构建。
1.4.3 各组织中GeCAT的表达分布特征及其在Cu2+胁迫下的应答模式
根据实验克隆得到的GeCATcDNA序列设计Real time PCR引物(表1),用RT-PCR法比较高原花斑裸鲤肝脏、肾脏、脑、鳃及肌肉组织中GeCAT在mRNA水平上的表达分布以及其在Cu2+胁迫下的应答模式,以β-actin为内参照。总RNA提取同前所述,用PrimeScriptTM RT reagent Kit试剂盒合成cDNA。RT-PCR体系为iQTM SYBR Green Supermix 10 μL;Ge-CAT-RTF1及Ge-CAT-RTR1 0.5 μL;ddH2O 8.0 μL;cDNA模板1.0 μL。扩增条件为95 ℃ 30 s ;95 ℃ 15 s;57 ℃ 30 s;72 ℃ 30 s,共39个循环。同一样本重复3次,单次实验重复3次,运用2-△△Ct法计算CAT的相对表达量。
1.5 统计分析方法
利用SPSS 20.0(IBM,USA)和OriginPro 8.0.0(OriginLab,USA)等软件对数据进行分析,差异性检验采用单因素方差分析(Analysis of variance,ANOVA),多重比较采用Student-Newman-Keuls。结果均以P<0.05作为显著性判断的标准。
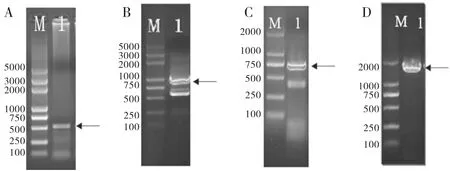
M:DNA分子量标准;PCR扩增结果(箭头所指为目标条带)
M:DNA Maker;PCR fragment(The arrow refers to the target product)
图1高原花斑裸鲤CATPCR产物电泳图
Figure1PCRproductsofGeCAT
2 结果
2.1 CAT全长序列的克隆及序列
根据1.2中的实验方法进行RT-PCR和RACE扩增,获得特异性片段条带(图1A)、5′-UTR条带(图1B)、3′-UTR条带(图1C)及通过Confirm PCR获得全长条带(图1D)。测序比对后得到全长cDNA序列编码区1665 bp、5′-UTR 158 bp和3′-UTR 440 bp,其ORF编码555个氨基酸残基。利用CDD分析GeCAT氨基酸序列保守结构域,其具有1个酶活性中心序列和亚铁血红素结合信号序列、3个催化位点残基、1个3'端加尾信号序列和多个亚铁血红素结合口袋(heme binding pocket)、NADPH结合位点(NADPH binding site)及多肽结合位点(polypeptide binding site)(图2)。

方框分别表示起始密码子ATG和终止密码子TGA;酶活性中心序列“FDRERIPERVVHAKGA”用加粗下划线标出;亚铁血红素结合信号序列“RLFSYPDTH”用方框标出;3个催化位点残基“H”“N”“Y”及其相应的核苷酸密码子用阴影标出;信号序列“AATAAA”用下划线标出;加粗下划线部分为亚铁血红素结合口袋;斜体加粗为NADPH结合位点.
The start codon“ATG”and the stop codon“TAA”are indicated by blocks;Enzyme active site sequences“FDRERIPERVVHAKGA”are indicated by bold and underline;and heme-ligand banding signal sequences“RLFSYPDTH”are box;The three conserved catalytic amino acids“H”,“N”and“Y”and their corresponding nucleotide codon are shadowed;The polyadenylation signal sequence“AATAA”is underlined.The boldface respresent heme binding pocket;The Italics bold respresent the NADPH binding site.
图2GeCATcDNA序列和氨基酸序列图
Figure2ThecompletecDNAsequencesandaminoacidsequencesofGeCAT
利用Vector NTI11对GeCAT分析同源性(表2)。再通过MEGA6.0构建邻接(neighbor-joining,NJ)系统发育树(图3)。经同源序列比对分析高原花斑裸鲤与草鱼(Ctenopharyngodonidella)、大西洋鲑(Salmosalar)、条石鲷(Oplegnathusfasciatus)、人(Homosapiens)、褐家鼠(Rattusnorvegicus)、鲢鱼(Hypophthalmichthysmolitrix)、斑马鱼(Daniorerio)、核盘菌(Sclerotiniasclerotiorum)等的同源性为33.6%至87.6%。系统进化聚类分析表明,GeCAT与草鱼、鲢鱼、斑马鱼的亲缘关系很近,聚为一枝,而与菌类的亲缘关系较远。
表2高原花斑裸鲤与其他物种CAT氨基酸序列同源性分析表

Table 2 Homology analysis of GeCAT amino acid sequences between Gymnocypris eckloni and other species
续表:

物种登录号与CAT同源性大西洋鲑Salmo salarNP_001133774.168.8%草鱼Ctenopharyngodon idellaACL99859.287.1%鲢鱼Hypophthalmichthys molitrixADJ67807.187.6%家蚕Bombyx mori NP_001036912.158.8%棉铃虫Helicoverpa armigeraXP_021187541.159.5%囊舌虫Bombyx moriSaccoglossus kowalevskiiXP_002738841.160.8%盘鲍Haliotis discus discusABQ60044.159.2%虾夷扇贝Mizuhopecten yessoensisAKV63251.160.8%合浦珠母贝Pinctada fucataADW0870061.3%酵母Saccharomyces sp.‘boulardii’KQC44730.140.1%曲霉菌Aspergillus flavusKOC11428.137.3%寄生曲霉Aspergillus parasiticusKJK68729.134.3%核盘菌Sclerotinia sclerotiorumEDN93275.133.6%

所引用的序列均来自GenBank All sequences and the accession numbers are from the GenBank database
图3GeCAT进化树分析图
Figure3ThephylogenetictreeanalysisofGeCAT
2.2 GeCAT的应答模式
2.2.1 各组织中GeCAT的表达分布特征
正常生理状态下,高原花斑裸鲤肝脏、肾脏、鳃、脑及肌肉等组织中GeCAT转录水平的表达表现出差异(图4)。GeCAT在肝脏中表达水平最高,脑、肾脏和肌肉组织次之,鳃中最低。

图4GeCAT在高原花斑裸鲤各组织中的表达分布图
Figure4ExpressionanalysisofGeCATintissuesofG.eckloni
2.2.2 Cu2+胁迫下GeCAT的应答模式
Cu2+胁迫下48 h内,高原花斑裸鲤GeCAT在不同时间段中肝脏、肾脏、鳃及脑组织中的转录水平表达量的变化表现出一定的规律性(图5)。Cu2+胁迫后48 h内,GeCAT最先在脑和鳃中应答,在胁迫12 h时表达量达到高峰;其次是在肾中应答,在胁迫24 h时表达量达到高峰,而在肝脏中胁迫48 h时表达量才有显著升高(P<0.05)。
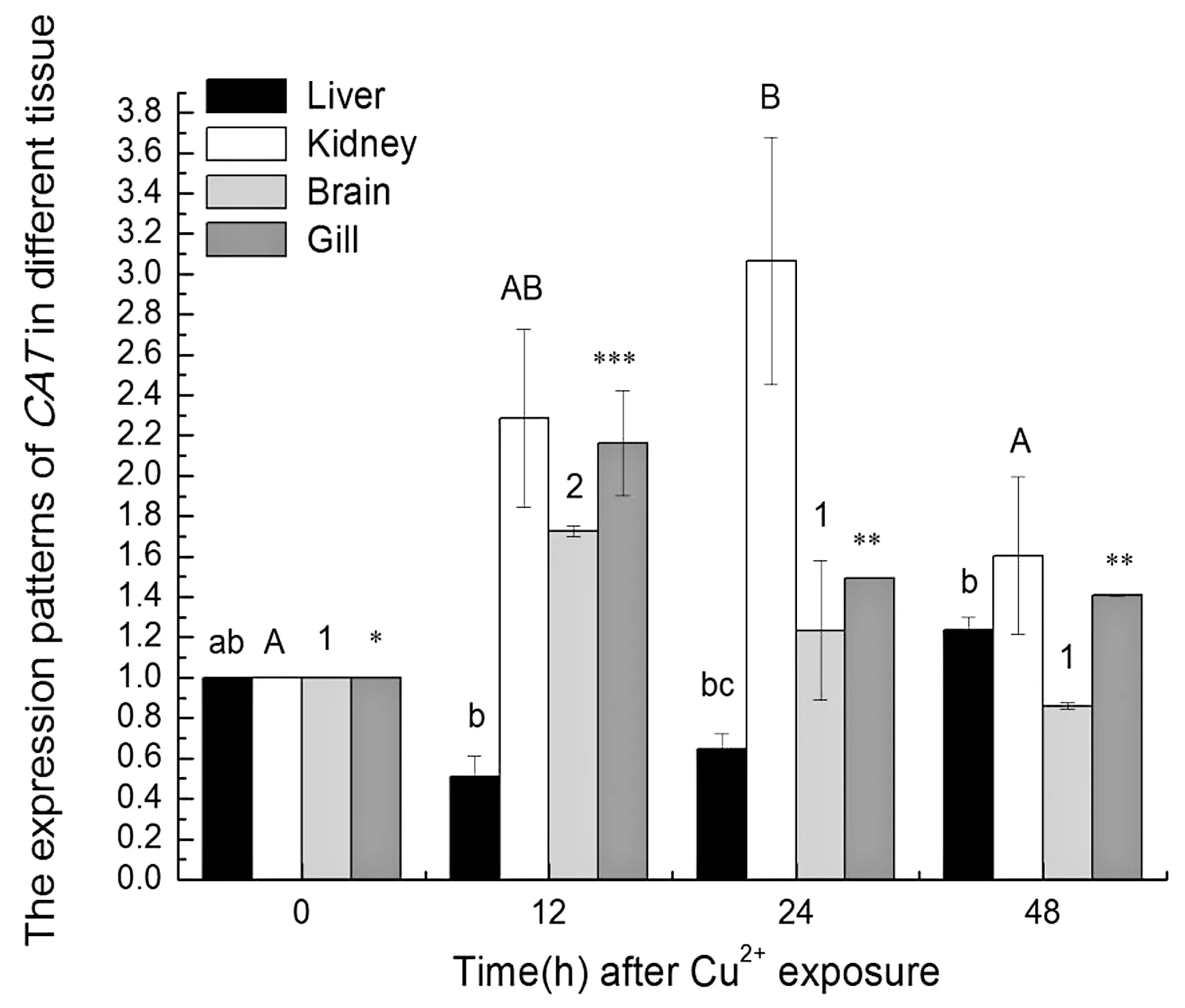
图5Cu2+胁迫下GeCAT表达模式图
Figure5TheexpressionpatternsofGeCATunderCu2+stress
3 讨论
保守序列结构分析表明,GeCAT属于单功能CAT。GeCAT的3′-UTR不含CA重复序列,这与草鱼[7]和斑马鱼[8-9]的CAT3′UTR具有较长CA重复序列不同,这表明在不同的水生动物中,CAT3′-UTR有着不同的序列特征。序列特征分析表明,GeCAT与鲢鱼、草鱼的同源性分别为87.6%、87.1%,花斑裸鲤同硬骨鱼的同源性都较高,在进化关系上表现出高度的保守性。
在Cu2+胁迫下,在不同组织中的GeCATmRNA水平相对表达量的变化趋势呈现出一定的规律性和组织特异性。在脑和鳃中,胁迫12 h时GeCAT表达量达到最高值(P<0.05);在肾脏中胁迫24 h时表达量达到最高值(P<0.05),而在肝脏中,胁迫48 h时表达量才有显著升高(P<0.05)。首先,这一结果表明,在本实验条件下,花斑裸鲤被检测的4个组织中GeCAT在mRNA水平产生了不同程度的应答。其次,不同组织中的GeCAT在mRNA水平产生应答的时间和量上表现出组织特异性。这与Hidalog等人[10]、Gul等人[11]的研究结果基本一致。分析其主要原因,一方面,Cu2+在本实验条件下的胁迫可能使花斑裸鲤体内产生大量的ROS,而ROS可以作为信号分子使核因子E2相关因子2(nuclear factor E2 related factor 2,Nrf2)与Keap1(Kelch-like相关蛋白1)发生泛素化解体,Nrf2发生核易位进入细胞核,先与小MAF蛋白结合,然后结合到抗氧化反应元件(Antioxidant Response Element,ARE),使靶基因(CAT、SOD等)转录激活[12],发挥抗氧化调节作用,保护机体免受氧化应激损伤。另一方面,在Cu2+胁迫下,Cu2+首先会作为必须元素结合到Cu/Zn-SOD上,使Cu/Zn-SOD先发挥作用,所以在Cu2+胁迫之初并没有观察到GeCAT表达量的升高,也可能是我们以国家渔业水质标准规定的浓度进行了胁迫,GeCAT表达量的变化与胁迫浓度相吻合。
GeCAT在肝脏中的表达变化最显著,Franchi等人[13]研究发现,肝脏是氧化反应最强的器官,其抗氧化酶活性最高,氧化反应最激烈。GeCAT在鳃组织中的应答比其他组织更早,可能是由于鳃长期暴露在水中并最先接触Cu2+而导致的。大脑含有大量的不饱和脂质,占用总需氧量的20%,很容易受到氧化损伤的影响[14]。所以,在Cu2+胁迫后,脑中GeCATmRNA表达水平与鳃一样最先进行应答。
综上所述,Cu2+胁迫下GeCAT在不同组织中的应答模式主要由Nrf2-ARE信号通路调控,并表现出明显的组织特异性,而Cu2+胁迫产生的ROS可以作为多条信号通路的信号分子,激活其他信号通路,所以花斑裸鲤在Cu2+胁迫下的应答调控机制以及GeCAT在不同组织中应答模式的调控机理都还需要进一步研究。本研究成功实现花斑裸鲤CAT的克隆鉴定表达,初步分析了花斑裸鲤CAT对Cu2+胁迫的应答模式,更好地揭示了CAT等抗氧化酶类与氧化应激之间的关系,并筛选出了有效生物标记物。
猜你喜欢
环球时报(2022-09-20)2022-09-20
第二课堂(课外活动版)(2021年6期)2021-01-18
今日农业(2020年24期)2020-12-15
家教世界·创新阅读(2020年4期)2020-06-03
当代工人·精品C(2020年1期)2020-05-20
家教世界·创新阅读(2020年1期)2020-05-11
家教世界·创新阅读(2020年2期)2020-04-07
作文周刊·小学二年级版(2019年12期)2019-04-26
小资CHIC!ELEGANCE(2015年14期)2015-09-23
小资CHIC!ELEGANCE(2015年15期)2015-09-01
