
南海北部表层沉积物中甘油二烷基甘油四醚随水深的分布特征
2018-12-06 03:37葛黄敏吴伟艳幸继联
厦门大学学报(自然科学版) 2018年6期
葛黄敏,吴伟艳,幸继联
(1.上海海洋大学 海洋科学学院,上海深渊科学工程技术研究中心,上海 201306;2.同济大学海洋地质国家重点实验室,上海 200092)
边缘海沉积物中保存着丰富的气候和海平面变化、洋流活动、沉积演化及生物地球化学过程等信息,是研究海-陆交互作用的很好材料[1].生物标志化合物具有特定的结构,可用来示踪其生物来源,对于了解边缘海的沉积物来源、重建古海洋环境变化和了解过去生物地球化学过程(如碳循环过程)非常重要.甘油二烷基甘油四醚(glycerol dialkyl glycerol tetraethers,GDGTs)是古菌和细菌细胞膜的特征生物标志化合物,广泛存在于陆地(如土壤、泥炭、热泉、湖泊及河流)和海洋环境[2].GDGTs的碳链与甘油分子通过醚键连接,其中古菌GDGTs(isoGDGTs)的碳链是类异戊二烯结构,甘油立体构型为2,3-双-o-烷基-sn-甘油;细菌GDGTs(brGDGTs)的碳链为非类异戊二烯结构,甘油立体构型为1,2-双-o-烷基-sn-甘油[2].isoGDGTs包含多种化合物,其中泉古菌醇(Cren.)通常被认为是海洋奇古菌(thaumarchaeota)的标志物,有一个同分异构体(Cren.′)[2].brGDGTs根据碳链上的甲基数和环数也可分为多类,普遍认为其来源于土壤细菌[2-4].边缘海作为衔接陆地与海洋的纽带,其中检测到的GDGTs通常是土壤、河流和海洋多种来源的叠加,它记录着丰富的环境信号,同时也增加了信号解析的难度.因此,探究边缘海沉积物中GDGTs的来源,是准确利用该标志化合物进行环境重建的基础.
南海是西太平洋热带海区最大的边缘海,海底地貌特殊,受不同海流的影响,其沉积物和有机质的来源与分布具有高度的空间差异性.边缘海的陆源信号主要通过河流输入实现,南海北部受珠江流域和红河流域陆源输入的显著影响[5].针对南海水体和沉积物中GDGTs的研究在过去几年正快速开展[6-16],但其中大部分以GDGTs已有环境指标的应用为重点,对GDGTs分布特征的研究相对较少.除了用基于GDGTs的指标重建环境外,深入探究GDGTs的组成随环境因素(如水深)的变化也十分重要.此外,越南北部湾作为南海的一部分,是典型的受河流输入影响显著的区域,目前还没有关于GDGTs的研究报道.本研究结合已发表的部分南海站点数据[7,13,16](水深52~4 182 m),以及北部湾浅水区(水深5~24 m)的表层沉积物样品,详见图1,分析isoGDGTs和brGDGTs组成随水深的变化规律,并分别探讨其来源.

图1 本研究涉及的南海北部表层沉积物站点分布
1 材料和方法
1.1 试剂和仪器
正己烷、二氯甲烷、甲醇和异丙醇(色谱纯,上海安谱实验科技股份有限公司).配备大气压化学电离(APCI)源的高效液相色谱-串联三重四级杆质谱(Agilent 6460 Triple Quad HPLC-MS),Prevail Cyano色谱柱(2.1 mm×150 mm,3 μm;Alltech,Deerfield,Illinois,USA).
1.2 样品采集与处理
2010年5月在越南海防海洋环境研究所(IMER)与同济大学的合作航次上,利用箱式采样器在越南北部湾7个站点采集了表层沉积物(图1).将沉积物样品采集到干净自封袋后,于-18 ℃下冷冻保存,在实验室将样品冷冻干燥48 h以上,然后在玛瑙研钵中磨成均匀粉末,保存在干净密封袋中,留作分析.
1.3 脂类提取
参照文献[7],采用超声法提取沉积物脂类:称取约10 g冻干研磨均匀后的样品,加入30 μL内标(C46-GDGT)[17],先用甲醇-二氯甲烷(体积比1∶3)超声萃取4次,再用甲醇超声萃取1次.用氮吹仪将萃取液吹干(水浴加热温度低于40 ℃),然后溶于3 mL正己烷-异丙醇(体积比99∶1),取1 mL用0.45 μm PFTE滤膜过滤后,再用氮吹仪吹干,并再次溶于300 μL的正己烷-异丙醇(体积比99∶1),超声溶解后待测.
1.4 仪器分析和指标计算
参考文献[3],采用HPLC-MS测定样品中GDGTs的含量:进样量为20 μL,色谱柱温保持40 ℃,流动相流速保持0.2 mL/min.色谱洗脱程序为:0~5 min内以正己烷-异丙醇(体积比99∶1)洗脱,5.01~45 min内梯度增加异丙醇的体积分数至1.8%,随后用正己烷-异丙醇(体积比99∶1)冲洗色谱柱10 min,再平衡10 min,使仪器回到初始状态,进行下一个样品的测试.采用APCI源进行化合物离子化,MS检测模式为选择性离子扫描(SIM),控制条件如下:雾化压力4.14×105Pa,雾化温度400 ℃;氮气流速6 L/min,温度200 ℃;毛细管电压3.5 kV;电流5 μA(约3.2 kV).目标GDGTs化合物m/z=1 302,1 300,1 298,1 296,1 292,1 050,1 048,1 046,1 036,1 034,1 032,1 022,1 020,1 018(由于大部分样品中m/z=1 048和1 046的信号峰都低于检测标准,所以最后统一放弃采集这两类化合物),标样C46-GDGT的m/z=744.通过各化合物离子峰[M+H]与加入内标的面积比值对GDGTs各组分进行定量,分别得到isoGDGTs和brGDGTs的含量.
进一步计算陆源输入指标(branched vs.isoprenoidal tetraether,BIT)[3]、环化指标(degree of cyclization,DC)[18]和甲烷指标(methane index,MI)[19],计算公式如下:
(1)
(2)

(3)
其中:w为含量,ng/g;Ⅰ、Ⅱ和Ⅲ分别为含有1,2和3个甲基的brGDGTs,Ⅰ a为含有1个甲基和1个碳环的brGDGTs,Ⅰ b为含有1个甲基和2个碳环的brGDGTs,以此类推;GDGT-i(i=1,2,3)表示含有i个碳环的isoGDGTs[7].
2 结果与讨论
2.1 isoGDGTs随水深的分布特征及来源
北部湾表层沉积物中总isoGDGTs的含量分布在20.5~162.3 ng/g之间,随水深增加而增加,isoGDGTs不同组分的相对丰度随水深(0~4 500 m)的分布如图2所示:其主要成分为GDGT-0(不含碳环的isoGDGTs)和Cren.,两者总和占isoGDGTs的77.5%~90.5%,GDGT-0的相对丰度变化范围为10.3%~23.1%(平均值为(18.4±3.0)%,n=50);Cren.的相对丰度整体上比GDGT-0高,变化范围为56.7%~80.2%(平均值为(63.3±5.4)%,n=50);GDGT-1,2,3和Cren.′是次要成分,总和占isoGDGTs的9.5%~22.5%,其中GDGT-3的相对丰度最低.
isoGDGTs各组分相对丰度随水深的变化趋势可分成3个阶段(图3):第1段 (<30 m),GDGT-0 的相对丰度随水深增加而降低,Cren.则随水深增加而增加;次要成分中,GDGT-2和GDGT-3均表现出随水深增加而降低的趋势;GDGT-1和Cren.′无明显的变化趋势,前者分布离散,后者相对稳定.这部分样品表征了北部湾近岸表层沉积物样品中isoGDGTs的分布特征.第2段(30~200 m),GDGT-0和Cren.的相对丰度均随水深增加而缓慢降低;而GDGT-2、GDGT-3和Cren.′则均随水深增加而增加,其中GDGT-2和Cren.′的增幅(61%)比GDGT-3的增幅(38%)大;GDGT-1仍然分布离散.第3段(>200 m),随水深增加GDGT-0和Cren.均未显示出明显的变化趋势,但前者的离散程度比后者大;次要成分中GDGT-1和Cren.′也未随水深出现明显的变化趋势,前者分布离散,后者相对稳定;GDGT-2和GDGT-3则随水深呈微弱下降的趋势.isoGDGTs不同组分含量之间呈现出显著的两两相关,相关系数R2为0.872~0.996(图4).
为了更好地展现isoGDGTs组成随水深的分布特征,计算了w(GDGT-2)/w(GDGT-3)和w(Cren.′)/w(GDGT-3)的值(图5(a)).结果显示,这2个比值将表层沉积物以水深200 m为界线分成了2组.水深≤200 m的表层沉积物中isoGDGTs的分布特征为:w(GDGT-2)/w(GDGT-3)<4,w(Cren.′)/w(GDGT-3)<2.水深>200 m的表层沉积物中isoGDGTs的分布特征为:w(GDGT-2)/w(GDGT-3)>4,w(Cren.′)/w(GDGT-3)>2.为了和Jia等[14]的结果进行对比,以w(GDGT-2)/w(GDGT-3) 为横坐标,以Cren.′的相对丰度为纵坐标作图(图5(b)),结果显示不同深度的样品也以200 m水深为界线分成了2组,其中Cren.′的相对丰度以4%为分界.而图3中0~30 m的样品并未表现出与30~200 m明显不同的比值分布区间(图5).这表明isoGDGTs组成在0~30 m的变化主要是北部湾近岸的区域性分布特征,但基于isoGDGTs的比值在整个0~200 m均反映了类似的微生物类群活动,可用于指示相关环境变化.

图2 isoGDGTs不同组分的相对丰度随水深的变化趋势

图3 isoGDGTs的相对丰度随水深的变化趋势
基于isoGDGTs的指标在古气候和古海洋方面的应用受到isoGDGTs组成变化的影响,详细研究表层沉积物中isoGDGTs的组成和来源对准确有效地运用现有环境指标十分重要.本研究综合分析了南海北部表层沉积物中isoGDGTs的组成分布特征,发现isoGDGTs的组成整体上是相似的,Cren.相对丰度最高,其次是GDGT-0,这和海洋奇古菌的纯培养结果一致[20-22].GDGT-0的来源比较丰富,包括广古菌(Euryarchaeota)中的产甲烷古菌(methanogens)和嗜甲烷古菌(methanotrophic archaea),以及海陆环境中广泛存在的奇古菌,而Cren.则主要来自奇古菌[23].Blaga等[24]提出用w(GDGT-0)/w(Cren.)判断湖泊环境中广古菌对isoGDGTs输入的影响,当样品中的w(GDGT-0)/w(Cren.)>2时,产甲烷古菌的输入影响较大.本研究中南海北部表层沉积物样品中w(GDGT-0)/w(Cren.)的平均值为0.30±0.07(n=50,图6),远低于2.Zhang等[19]提出了评价嗜甲烷古菌对isoGDGTs影响的MI,当MI小于0.3时,isoGDGTs受嗜甲烷古菌的影响不大.本研究结果显示MI的平均值为0.17±0.02(n=50,图6),明显小于0.3.此外,Sinninghe Damsté等[25]也提出当w(GDGT-0)/ (w(GDGT-0)+w(Cren.))>67%和w(GDGT-2)/(w(GDGT-1)+w(GDGT-2)+w(GDGT-3)+w(Cren.′))> 45%时,表明isoGDGTs的来源除奇古菌外还有其他古菌类群的贡献.本研究中,这两个比值分别为(24±4)% (n=50)和(37±2)% (n=50).上述指标结果均表明,南海北部表层沉积物中isoGDGTs主要来自于海洋奇古菌,广古菌中产甲烷古菌或嗜甲烷古菌的贡献均不明显.
isoGDGTs的组成随水深变化的特征没有体现在主要成分Cren.和GDGT-0上,而是体现在次要成分的变化上.以200 m为界线,isoGDGTs明显随水深分成了2组(图5),无论是isoGDGTs单组分的相对丰度,还是isoGDGTs的各项指标,都随水深表现出截然不同的变化特征.已有的基于海洋悬浮颗粒物(SPM)和表层沉积物的isoGDGTs的研究观察到w(GDGT-2)/w(GDGT-3)随水深的增加而增加[25-29],认为深水古菌相对于浮游古菌而言具有更高的w(GDGT-2)/w(GDGT-3)和w(Cren.).在中国南海区域,Wang等[11]报道了珠江口及邻近陆架浅水区表层沉积物中GDGT-2和GDGT-3相对丰度的变化,发现研究海域w(GDGT-2)/w(GDGT-3)偏低,为1~4,而在南海深海区该比值偏高,为4~10.Jia等[14]和郭威等[30]在珠江口和南海不同季节的SPM及表层沉积物中也发现了类似规律,对比水深<154 m和水深>200 m的表层沉积物,后者w(GDGT-2)/w(GDGT-3)明显更高.杨义[15]基于南海63个表层沉积物样品的GDGTs研究,发现水深<200 m时,GDGT-2和Cren.′含量与水深之间呈现很好的线性关系,水深>1 000 m时,则未与水深表现出明显的相关性,但GDGT-3含量随水深表现出逐渐减小的趋势.

GDGT-5和5′分别表示Cren.和Cren.′.
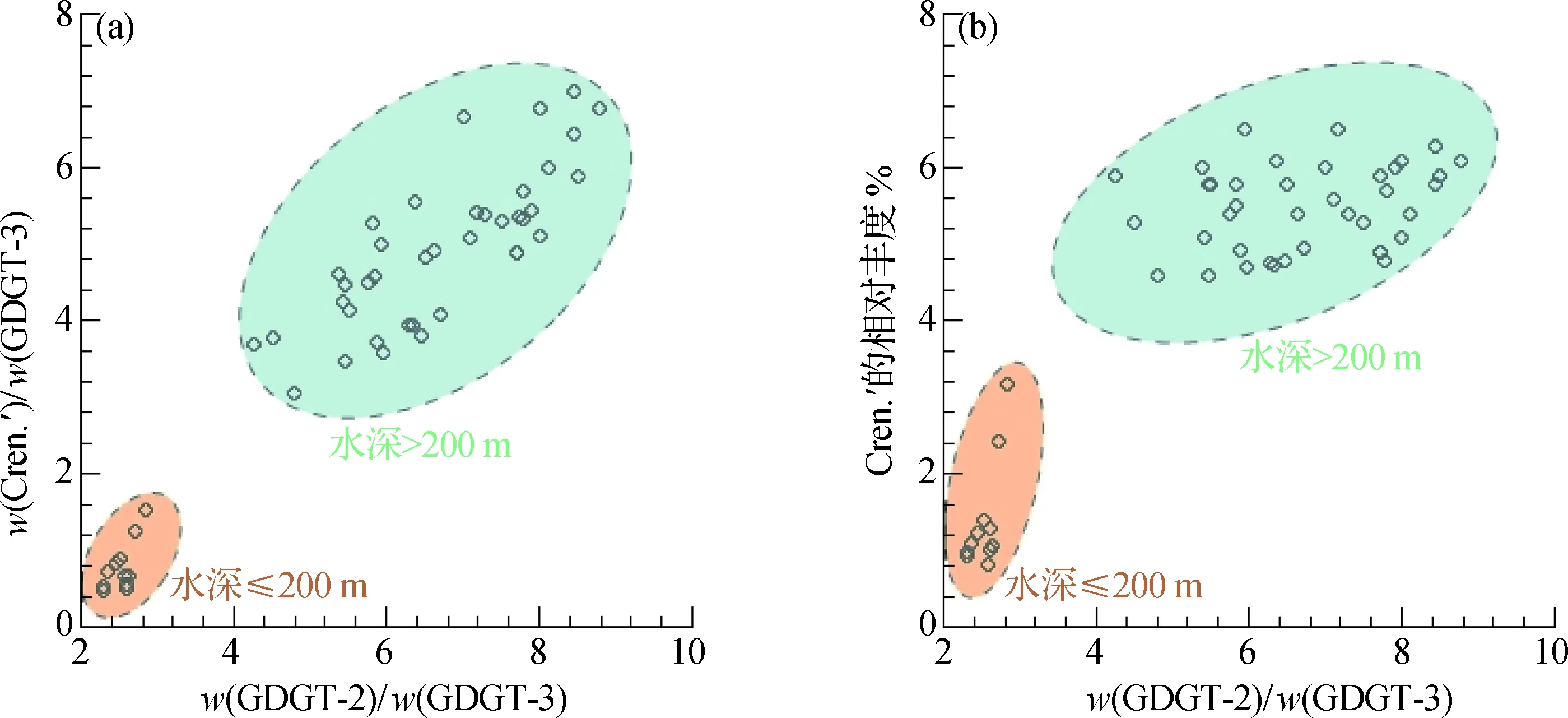
图5 w(GDGT-2)/w(GDGT-3)与w(Cren.′)/w(GDGT-3)(a)以及与Cren.′的相对丰度(b)之间的交叉分析图

图6 w(GDGT-0)/w(Cren.)(橙色)和MI(蓝色)随水深的变化趋势
上述研究结果表明,不同水深对应的古菌类群不同,不同类群之间的最主要差别可能表现在产生GDGT-2和GDGT-3的能力上.不同水深的沉积物中,微生物群落的分布较复杂,很大程度上受到地理位置的影响[31-32],而不同水深微生物群落分布的差别可能引起GDGTs组成的变化.杨义[15]发现在水深1 000 m 上下,会有较多的GDGT-2和相对较少的GDGT-3的贡献,而在碳酸盐补偿深度(3 500 m)以下,古菌群落可能受碳酸盐补偿的影响而发生变化.Kim等[28-29]也提出深海奇古菌会产生相对较多的GDGT-2和相对较少的GDGT-3.尽管上述研究都认为深水区沉积物中w(GDGT-2)/w(GDGT-3)高值是由深水奇古菌贡献的,但目前没有深水奇古菌的纯菌株培养,无法确切了解深水奇古菌与浅水奇古菌isoGDGTs的组成差异.海洋中的浮游古菌主要包括两种类型:奇古菌(marine groupⅠ)和广古菌(marine groupⅡ),其中广古菌是海洋表层水体中最主要的古菌类型[33-34],而奇古菌除了存在于表层水体外,也是深水古菌的主要类型[33].Lincoln 等[34]基于海水中古菌和isoGDGTs的分布特征,推测广古菌也有可能产生Cren..Wang等[12]在珠江口水体中发现广古菌可能产生GDGT-1,2,3,也会对isoGDGTs的组成产生影响.因此,南海不同水深的表层沉积物中,isoGDGTs的组成存在明显差异,极有可能是不同水深的海洋奇古菌贡献所致,但不能排除广古菌的影响.其中,水深200 m(以浅是陆架区,以深是远海深海区)是导致GDGTs相关参数变化的一个关键深度,所以GDGTs环境指标的应用首先要区分所研究区域处于何种海洋环境背景下,在此基础上再探讨相关参数背后的环境意义.如能根据浅水区和深水区isoGDGTs的组成差异建立指标,用于指示水深或海平面的变化,将会是开展古海洋古环境研究的又一有利手段.

图7 brGDGTs不同组分相对丰度随水深的变化趋势
2.2 brGDGTs随水深的分布特征及来源
北部湾表层沉积物中总brGDGTs的含量分布范围在23.1~83.1 ng/g之间,随水深没有明显变化规律,其中不含环的Ⅲ+Ⅱ+Ⅰ占总brGDGTs的相对丰度为68.8%~76.1%,随水深增加略微减少.结合已发表的数据,南海北部表层沉积物中brGDGTs 整体上以Ⅲ、Ⅱ和Ⅰ为主,其相对丰度分别为3.3%~52.3% (平均值为(20.4±11.8)%,n=50),10.8%~27.5%(平均值为(20.4±3.5)%,n=50)和22.0%~55.1%(平均值为(36.1±8.9)%,n=50),其中Ⅲ的波动范围最大.次要成分中Ⅱb、Ⅱc、Ⅰb和Ⅰc的相对丰度分别为0~12.2%(平均值为(7.0±2.8)%,n=50),0~5.3%(平均值为(1.9±1.6)%,n=50),0~15.2%(平均值为(8.9±3.9)%,n=50),0~12.5%(平均值为(5.3±3.6)%,n=50).brGDGTs 随水深的分布特征可分成2段(图7):第1段,水深≤200 m,表层沉积物的Ⅲ、Ⅱb、Ⅱc 和Ⅰc的相对丰度随水深增加而增加,而Ⅰ的相对丰度随水深增加而减少,Ⅱ和Ⅰb 的相对丰度保持稳定.第2段,水深>200 m,随水深增加,Ⅲ的相对丰度先增加后降低,Ⅰc的相对丰度减小;此范围内的brGDGTs组成随水深波动开始较大,在2 200 m之后趋于平稳,体现在Ⅲ、Ⅱb、Ⅱc 和Ⅰc的相对丰度变化上,而Ⅱ和Ⅰb 的相对丰度依然保持相对稳定.brGDGTs不同组分含量之间的相关性(图8)显示:Ⅱ 与Ⅰ和Ⅰb,以及Ⅰ与Ⅰb的正相关性均很好,相关系数R2分别达到0.808,0.879和0.875;此外,Ⅲ与Ⅱb和Ⅱc的相关性也很好,R2分别为0.709和0.806.
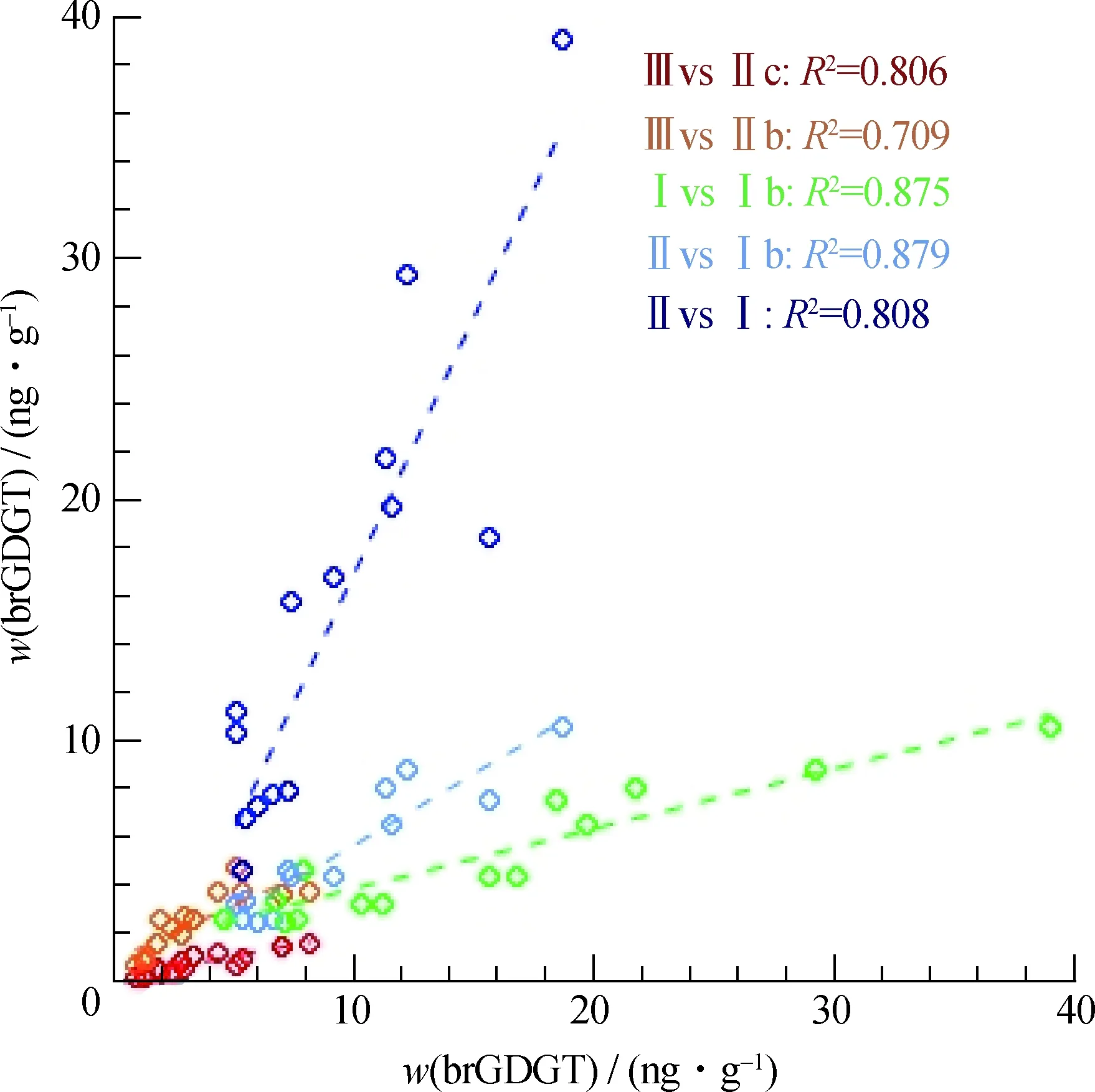
图8 brGDGTs不同组分间的相关性
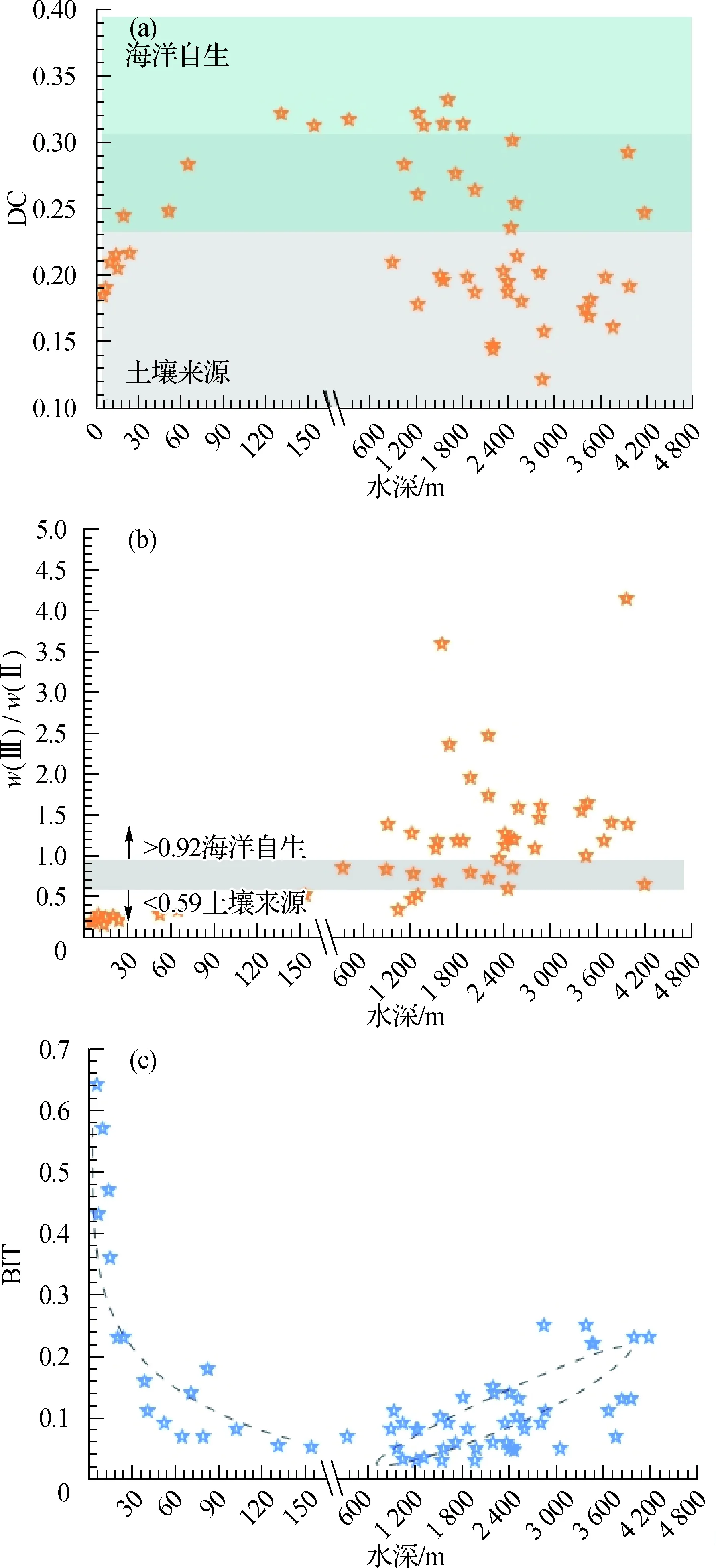
图9 DC(a),w(Ⅲ)/w(Ⅱ)(b)和BIT(c)随水深的变化趋势
总的来说,在浅水区(≤200 m),Ⅰ+Ⅱ+Ⅲ的相对丰度随水深增加而有所减少(从76.1%降至60.6%),可能是因为海洋自生的brGDGTs跟陆源brGDGTs相比,会产生更多含五元环的brGDGTs.Sinninghe Damsté等[18]提出用DC表征brGDGTs中五元环的变化趋势.Weijers等[23]计算了不同类型样品的DC值,其中全球范围内土壤的DC值平均为0.16±0.15,而海洋沉积物的DC值平均为0.40±0.16.Xing等[35]计算了渤海和黄海沉积物的DC值(0.32~0.57,平均值0.43)并由此推证渤海和黄海中检测到的brGDGTs有海洋自生来源的贡献.本研究中,DC值分布范围为0.12~0.33,随水深呈先升高(<200 m,0.18~0.31)后降低(200~3 000 m,0.33~0.16)再升高(>3 000 m,0.16~0.29)的趋势(图9(a)).因为Weijers等[23]的研究结果误差范围较大,其中当DC值为0.24~0.31时,既包含陆地信号,也包含海洋信号,所以仅凭DC值难以判断brGDGTs的来源.Xiao等[36]通过调查全球范围内的土壤和海洋沉积物样品,发现90%以上土壤样品中w(Ⅲ)/w(Ⅱ)<0.59,而90%以上的海洋沉积物样品中w(Ⅲ)/w(Ⅱ)>0.92,由此提出w(Ⅲ)/w(Ⅱ) 可用以指示brGDGTs的来源.本研究中w(Ⅲ)/w(Ⅱ) 分布范围为0.16~4.14,平均值为1.09±0.80(图9(b)),指示明显的海洋自生brGDGTs信号,尤其是水深>200 m的样品中,w(Ⅲ)/w(Ⅱ) 值显著分布在海洋自生区间.此外,用于指示来源于陆地土壤的有机物对水体环境贡献量的BIT[3],在南海北部62个表层沉积物样品中也呈现出随水深变化的分布特征(图9(c)).BIT最高值(0.64)出现在水深最浅的沉积物中,在水深≤200 m时BIT值随水深增加而降低(0.64~0.05),其中在<40 m 范围内快速下降到0.1左右,40~200 m随水深缓慢降低.在水深>200 m的沉积物中,BIT值(0.03~0.25,平均值为0.10±0.06)虽然分布离散,但整体呈现随水深增加而升高的趋势.该水深范围内,Cren.含量保持平缓,而brGDGTs略有上升,应该是造成BIT值呈上升趋势的原因.因此,水深>200 m时BIT值的缓慢上升趋势也暗示了海洋自生brGDGTs的存在.结合之前探究brGDGTs来源的相关研究结果[14,23,37-38],可以认为在浅水环境(≤200 m),海洋沉积物中brGDGTs反映的主要是陆源信号,而在深水环境(>200 m),brGDGTs则显示出陆源输入和海洋自生的双重贡献.关于BIT对陆源输入的指示作用,在浅水区尚能适用;在深水区,尤其是具有明显证据显示海洋自生brGDGTs贡献时,BIT值的升高并不能用于指示陆源输入增加,而关于其在深水区有何指示作用尚待进一步研究.
3 结 论
本研究利用北部湾7个表层沉积物及南海北部已发表的55个表层沉积物的GDGTs数据,通过单个GDGT化合物相对丰度随水深的分布特征、不同GDGTs之间的相关性分析以及基于GDGTs的多个比值和指标的分析,探究了南海北部表层沉积物中GDGTs随水深的分布特征及来源问题.结果显示,以200 m为界,GDGTs不同化合物及相关指标均呈现显著的分段式分布趋势:
1) isoGDGTs各组分的相对丰度随水深的变化趋势可分成3个阶段:<30 m,30~200 m,>200 m.水深≤200 m时w(GDGT-2)/w(GDGT-3)<4,w(Cren.′)/w(GDGT-3)<2,而当水深>200 m时,w(GDGT-2)/w(GDGT-3)>4,w(Cren.′)/w(GDGT-3)>2.这种变化趋势显示不同水深对应的古菌类群可能不同,而类群之间最主要的差别可能表现在产生GDGT-2和GDGT-3的能力上.
2) 在浅水环境(≤200 m),海洋沉积物中brGDGTs反映的主要是陆源信号,而在深水环境(>200 m), 海洋沉积物中brGDGTs则显示出陆源输入和海洋自生的双重贡献.随着水深增加,brGDGTs的环化增强而甲基化减弱,说明海洋自生的brGDGTs相对陆源输入的brGDGTs来说,更易产生环化结构而较少产生甲基化结构,这种差异性可能是微生物群落适应环境的结果.
3) 越来越多的证据表明GDGTs在不同环境中来源不同,因此在应用全球性的GDGTs指标时,应尝试校正区域性因素导致的偏差.
致谢:感谢张洪瑞在图1绘制时提供的帮助,感谢贾国东教授和Harunur Rashid教授对文章校正提出的宝贵意见.
猜你喜欢
河北水利(2022年10期)2022-12-29
海洋通报(2022年4期)2022-10-10
土壤学报(2022年3期)2022-08-26
大自然探索(2022年5期)2022-07-11
知识就是力量(2022年6期)2022-06-16
海洋信息技术与应用(2022年1期)2022-06-05
海洋渔业(2021年4期)2021-08-10
北京大学学报(自然科学版)(2020年3期)2020-06-07
海洋渔业(2018年3期)2018-06-29
农村经济与科技(2018年24期)2018-05-14
