
南海北部天然气水合物的形成分解与微生物的偶联关系
2018-12-06 03:58雷怀彦潘富龙程伟东
厦门大学学报(自然科学版) 2018年6期
孔 媛,雷怀彦,2*,许 江,王 斌,潘富龙,张 劼,陈 勇,程伟东
(1.厦门大学海洋与地球学院,2.近海海洋环境科学国家重点实验室(厦门大学),福建 厦门 361102;3.国家海洋局第三海洋研究所,福建 厦门 361005)
天然气水合物是一种笼形的、其中充填甲烷等小分子烃类气体为主的冰状混合物,理想结构下其甲烷含量是标准状态下的164~184倍[1],由于其分布广、埋藏浅并且资源量巨大,单位体积的天然气水合物燃烧所释放的能量远高于常规天然气,所以是一种重要的新型清洁能源.天然气水合物是甲烷的巨大储库,其形成和分解控制着进入海洋乃至大气的甲烷通量,对海洋生态环境的变化(大洋缺氧、海洋酸化等)甚至气候变化的影响巨大[2-5],也是诱发海底滑坡等海底地质灾害的重要原因[6-9].天然气水合物的形成和分解受多方面因素的影响,其中微生物起着十分重要的作用.在形成方面,微生物降解有机质产生的甲烷为天然气水合物的形成提供了必要的气源条件,据统计,大多数海域的天然气水合物主要以甲烷为主,该甲烷来源主要为微生物成因气[10].而在分解方面,虽然目前没有直接证据表明天然气水合物的分解与微生物有关,但是Lanoil等[11]在天然气水合物内部检测到的细菌与古菌具有生成和氧化甲烷的功能,暗示其与天然气水合物的稳定性、气体组成以及地球化学特征有关;另外,天然气水合物溶解[12]或失稳分解后释放的大部分甲烷会在微生物的作用下发生甲烷厌氧氧化(anaerobic oxidation of methane,AOM)反应而被消耗掉[13].因此,在天然气水合物的形成、溶解或分解情况下,沉积物中将会形成某种或某几种特定微生物聚集的现象,而在一定条件下,大量的单一或有限属种的生物聚集能够产生一种或少数几种具诊断性的生物标志化合物[14].沉积物中特定的微生物群落及其在沉积物中保留的相关生物标志化合物,为利用微生物勘探天然气水合物提供了相应的依据和方向.
我国南海北部具有良好的天然气水合物成藏的地质条件,是天然气水合物调查与研究的重点海域.据《2015中国国土资源公报》显示,在南海北部神狐海域发现天然气水合物,圈定矿藏面积达128 km2,目前已有多位学者在南海北部圈定天然气水合物潜在区,例如台西南盆地、琼东南盆地以及东沙海域的部分区域[15-19].与天然气水合物形成和分解相关的矿物、沉积物和孔隙水地球化学以及微生物等方面的研究也屡见报道,其中关于微生物及其标志化合物的识别与天然气水合物的形成、分布、分解和成岩(如自生碳酸盐岩)等方面的研究不仅为利用微生物勘探天然气水合物提供了有效的依据,还对海洋环境以及气候变化研究具有重大意义.
1 产甲烷微生物与天然气水合物形成的关系
充足的气源是天然气水合物形成的必要条件,天然气水合物的主要气体成分为甲烷,还含有乙烷、丙烷等同系物以及CO2、N2、H2S等其他气体,其中甲烷体积分数超过99%的天然气水合物通常又被称为甲烷水合物.据研究统计,世界大部分天然气水合物的甲烷来源均为微生物成因气[10],微生物成因气的碳氢化合物气体成分比值(R=n(CH4)/[n(C2H6)+n(C3H8)])较高,一般大于1 000,δ13C值一般小于-55×10-3[20].在我国南海天然气水合物钻探区所取得的沉积物以及天然气水合物样品,其气体组成主要为微生物气或者以其为主的混合气[21-22],因此,探讨微生物生成甲烷的机制以及条件对天然气水合物的形成以及赋存环境具有重要意义.
1.1 产甲烷微生物分类及其作用机制
微生物成因气是指在成岩作用或有机质演化的早期阶段,由不同微生物种群协同作用下生成的以甲烷为主的天然气,其形成过程主要分为3个阶段,分别为水解发酵阶段、产氢产乙酸阶段和产甲烷阶段[23],前两个阶段由多种微生物协作完成,而产甲烷阶段主要由产甲烷菌(methanogens)实现.
产甲烷菌属广古菌门[24],是绝对厌氧菌,依赖于其他微生物将有机质降解为简单化合物再加以利用,是有机质彻底分解及海洋沉积物中物质循环的重要组成部分.目前有3种已知的产甲烷途径,即H2/CO2还原、甲基还原和乙酸裂解[24],超过70%的产甲烷菌都能利用H2/CO2产生甲烷[25].大多数产甲烷菌生长pH范围为6.0~9.0,pH过高或者过低都会影响产甲烷菌的生长及其产甲烷效果.不同类型的产甲烷菌有各自不同的最适生长温度,且该最适生长温度范围较大,其中嗜冷产甲烷菌最适生长温度<25 ℃,极端嗜热产甲烷菌最适生长温度>80 ℃.此外产甲烷菌对于环境的氧化还原电位有严苛的要求,其生长的最适氧化还原电位在-320 mV以下,在-160 mV时仅缓慢生长.目前已经分离得到的产甲烷菌已有200多种,形态有规则或不规则的球形、杆状、螺旋状、八叠球状等,还有的能连成长链状,也有一些不规则形状如平盘状[26];目前分为7个目,分别是甲烷杆菌目(Methanobacteriales)、甲烷球菌目(Methanococcales)、甲烷微菌目(Methanomicrobiales)、甲烷八叠球菌目(Methanosarcinales)、甲烷火球菌目(Methanopyrales)[24]、甲烷胞菌目(Methanocellales)[27]以及近来发现的Methanomassiliicoccales[28],其中从海洋沉积物中分离的主要为甲烷球菌目、甲烷微菌目、甲烷八叠球菌目和甲烷火菌目,其代谢途径包括目前已知的3种产甲烷途径,最适生长温度范围较大,详细信息见表1.
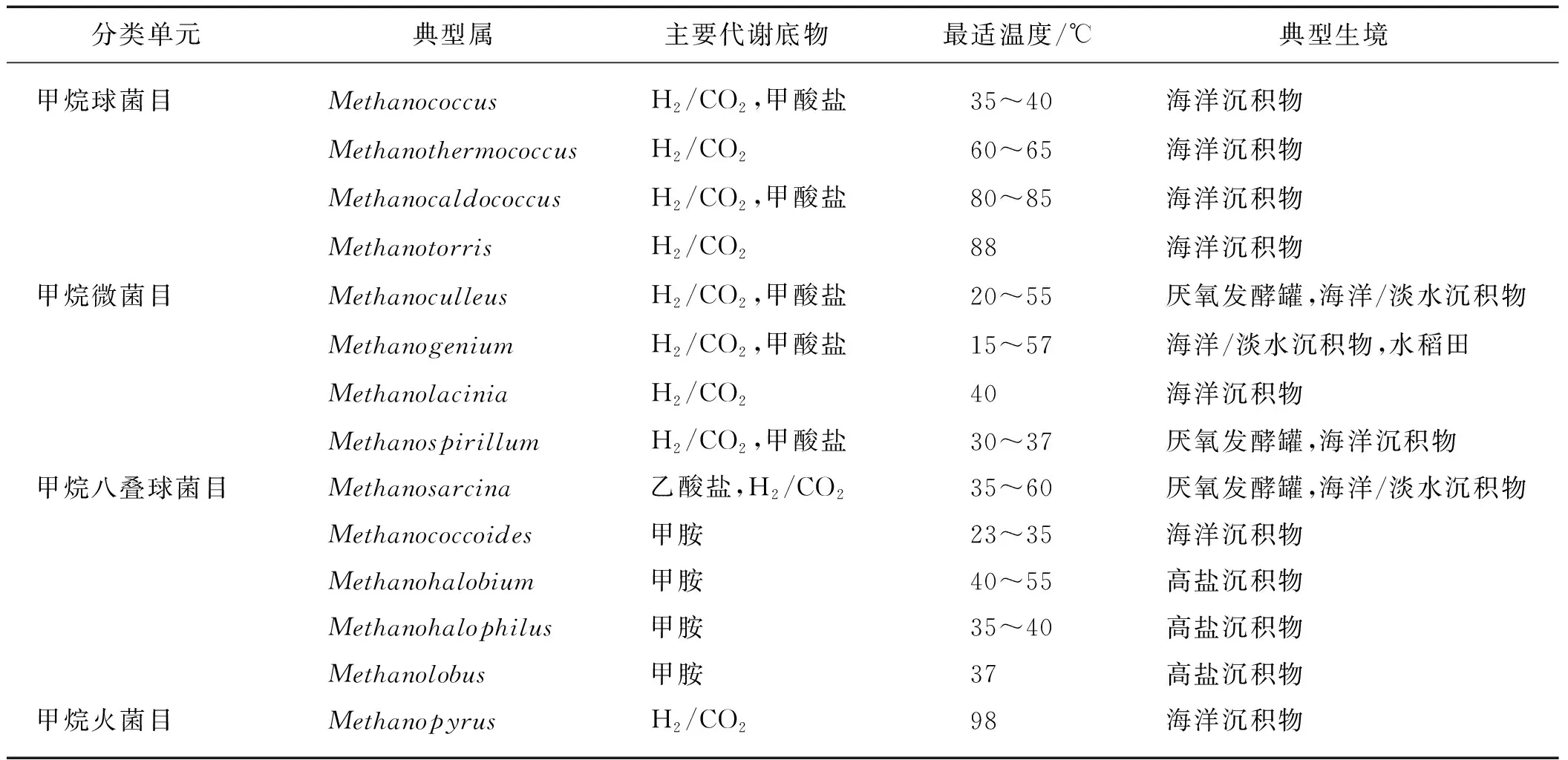
表1 海洋沉积物中产甲烷菌的主要类群及其生理特征[24]
产甲烷菌的细胞壁类脂物具有类异戊二烯结构,目前报道的其特有的生物标志化合物包括2,6,10,15,19-五甲基二十烷烃(Ⅱ)[29]、2,6,10,15,19,23-六甲基二十四烷烃、头-头连接的C28-C40无环类异戊二烯烷烃[30]等.由于微生物对碳同位素具有分馏作用,其代谢作用生成的生物标志化合物具有较低的稳定碳同位素组成(δ13C).另外,产甲烷菌含有独特的催化完成产甲烷最后一步反应的催化酶,即甲基辅酶M还原酶(methyl coenzyme-M reductase,MCR),可以作为检测和区分产甲烷菌的分子标记[31].
在海洋沉积物中,主要在富有机质的厌氧环境中,尤其是硫酸盐穿透深度以下的沉积物中发现微生物产甲烷.这是由于海洋环境中存在大量硫酸盐,有机质在堆积和埋藏过程中首先在硫酸盐还原菌(sulfate-reducing bacteria,SRB)的作用下被氧化分解,当绝大部分硫酸盐被消耗之后,产甲烷作用才开始发生.综上所述,微生物产甲烷首先要有充足的碳源,即有机质;其次要有适宜产甲烷菌的生长环境,即适宜的温度和pH等;另外还需要还原环境以及无SRB的竞争.
1.2 微生物产甲烷形成天然气水合物的条件
沉积物中有机质的来源和特性(活性)是形成天然气水合物的控制因素.有关研究表明,高含量的陆源有机质的降解可促进微生物的产甲烷作用,产生大量甲烷从而形成天然气水合物[32].快速沉积使得沉积物中的陆源有机质得以保存并降解,提供CO2以及乙酸等,有利于微生物产生大量的甲烷;同时沉积速率高的区域易形成欠压实区,可构成良好的流体输导体系并形成有利的储集空间,有利于甲烷的聚集和保存,为天然气水合物成藏提供了充实的气体来源;且沉积速率高使得盆地热流值低,有利于天然气水合物的形成.有研究认为当沉积速率大于3 cm/ka且有机碳含量高于0.5%时,就可满足天然气水合物形成所需的生物成因甲烷[31].
大多数海洋环境都符合天然气水合物稳定带形成的热力学条件,然而目前只有部分海域发现了天然气水合物,其形成主要还是受控于沉积物中可用甲烷的量[33].天然气水合物稳定带内的部分有机质通过微生物作用转化为甲烷,浓度高于水-天然气水合物二相平衡溶解度的溶解甲烷将形成天然气水合物,甲烷的供应则主要包括来自深部流体所携带的溶解甲烷以及微生物作用原位生成的甲烷[34].通常被动大陆边缘的陆源有机质输入高,能生成足以形成天然气水合物的甲烷[34-35],如Cascadia大陆边缘的水合物脊附近,分散型天然气水合物的甲烷即来自于微生物原位产甲烷作用[36].而增生大陆边缘更多为大量向上运移的流体所带来的甲烷[37],微生物原位产甲烷作用对其天然气水合物形成的贡献较低,如美国俄勒冈州外海的水合物脊,实测的原位甲烷生成速率为4×10-6mol/(m3·a)[38].模拟计算结果表明,孔隙水溶解甲烷的对流作用是天然气水合物成藏的主要甲烷供给方式,其次是扩散作用,微生物原位产甲烷作用最小[39].然而,海洋大部分天然气水合物的气体来源于微生物作用产生的甲烷,因此除了微生物原位产甲烷的贡献外,还有微生物产的甲烷通过对流和扩散作用以及一定的气体运移输导通道,运移到有利的圈闭条件下形成的一定规模的天然气水合物.
2 天然气水合物的分解与AOM的关系
海洋沉积物中天然气水合物的稳定主要受外部环境的控制,如水深、海底温度、地温梯度、孔隙水盐度、沉积物的物理和化学性质.一旦天然气水合物稳定存在的条件被破坏,将导致其分解并释放出甲烷,甲烷在沉积物中扩散运移的过程中,如果碰到合适的温压条件可以二次生成天然气水合物,或者在厌氧环境中微生物的作用下发生AOM反应而被消耗掉[40].AOM会改变沉积物中微生物的群落组成特征,形成大量生物标志化合物,因此,识别沉积物中参与AOM的微生物群落特征及其生物标志化合物对于指示下伏地层中是否有天然气水合物藏具有重要意义.
AOM是海洋沉积物中消耗甲烷的主要途径,通常与硫酸盐还原作用(sulfate reduction,SR)同时发生,并在垂向上形成较宽的硫酸盐-甲烷转换带(sulfate-methane transition zone,SMTZ),其位置主要是由深部向上渗漏的甲烷通量决定[41],在SMTZ内SR-AOM速率达到最高.目前已知有3种甲烷厌氧氧化古菌(anaerobic methanotrophic archaea,ANME)参与到AOM反应中,即ANME-1、ANME-2[42]和ANME-3[43].ANME通常与SRB(如Desulfosarcinales和Desulfobulbus[44-45])伴生,但其中ANME-1和部分ANME-2类型被发现可独立存在[42,46].通过研究发现AOM也可使用其他电子受体,与金属(铁、锰)还原[47-49]以及硝酸盐、亚硝酸盐还原[50-52]同时发生.然而到目前为止,ANME和SRB等古菌和细菌均未获得纯培养,人们对其认识还十分有限.随着分子生物技术、同位素示踪、富集培养等研究手段的发展,相关古菌和细菌的分布、多样性、形态以及代谢功能等研究已逐渐丰富.基于16SrRNA基因的检测,目前公布的有大约超过1 800条与ANME相关的16SrRNA基因序列[53],目前已知的ANME都属于广古菌门,ANME-1与Methanosarcinales和Methanomicrobiales有亲缘关系,但在进化树上位置相对疏远;ANME-2与Methanosarcinales古菌近源,可进一步细分为a、b、c和d亚群,根据最近的研究,ANME-2d被分作一个新的类群[52];ANME-3在进化树上与Methanococcoides距离比较近.Knittel等[54]总结了截至2009年通过荧光原位杂交(FISH)以及酶联荧光原位杂交(CARD-FISH)方法所获得的ANME以及ANME和SRB细胞团等的显微照片,目前ANME的主要形态有长方形(单个ANME-1细胞)、球状(单个ANME-2c细胞)、和SRB(如Desulfosarcina)形成的细菌席、和SRB形成的混合状细胞团以及贝壳状混合细胞团等.
特殊的生物标志化合物及其δ13C特征被广泛用于识别自然环境中的ANME.在现代天然气渗漏和水合物发育区,根据参与的微生物所产生的生物标志化合物可分为ANME来源的醚类异戊二烯类和类异戊二烯烃类,以及SRB来源的二烃基甘油二醚(DGDs)和脂肪酸(fattyacids)[55-57].醚类异戊二烯类主要有古醇(archaeol)、sn-2-/sn-3-羟基古醇(sn-2-/sn-3-hydroxyarchaeol)、头-头相连无环和环状的甘油二烷基甘油四醚(iGDGTs);类异戊二烯烃类主要有尾-尾相连、不规则的类异戊二烯烃及其不饱和衍生物,有2,6,10,15-四甲基十六碳烷(2,6,10,15-tetramethylhexadecane)、2,6,10,15,19-五甲基二十烷烯(2,6,10,15,19-pentamethylicosene,PMI)、双植烷(biphytane)、角鲨烷(squalane)、植醇(phytanol) 和 植 烷 酸(phytanic acid)以及不规则的含0~6 个双键的 2,6,10,14,19,23-六甲基二十四碳烯和含有 0~7个双键的2,6,10,14,19,23,27-七甲基二十八碳烯等.DGDs包括DGDs-Ⅰ和DGDs-Ⅱ两个系列,脂肪酸则主要有异构/反异构脂肪酸(iso-/anteiso-C15,C17)、环丙基C17脂肪酸等.可依据这些生物标志化合物的浓度及其δ13C识别自然环境中的ANME,在墨西哥湾和黑海水合物赋存区的海洋沉积物中检测出的古醇类、iGDGTs和二醚类(diethers)等脂肪醇和醚类化合物,其浓度较正常海洋沉积物中的高数倍[58];天然气水合物失稳释放出的甲烷在微生物作用下发生AOM,其生物标志化合物通常具有极低的δ13C,一般低于-50×10-3[59],最低的可低于-130×10-3[60],极低的δ13C说明微生物在代谢过程中利用了来自天然气水合物渗漏的甲烷,在该富集层位存在ANME和SRB活动.另外,不同的微生物类群具有不同的生物标志化合物特征,如与ANME-2和ANME-3相比,ANME-1的sn-2-羟基太古醇和古醇的所占比例较低,并且古醇的δ13C相对较低;同时,古菌种群ANME-2比ANME-1对甲烷的碳同位素的分馏作用更强,使得在以ANME-2/Desulfosarcina为主要微生物种群的冷泉环境中,其代表性生物标志化合物具有更低的δ13C[59].
3 南海北部天然气水合物研究
我国南海平均水深1 000 m以上,陆坡区水深一般在300~3 000 m,海底温度大致在0~5 ℃,地温梯度(钻孔中测得14~76 ℃/km,平均35 ℃/km)适中[61],沉积速率高,具备形成天然气水合物的有利地质条件[62].
目前在南海已圈定多处天然气水合物远景区,并多次钻获实物样品,且多处海域发现活动冷泉及冷泉形成的碳酸盐岩,海底冷泉的发育一般与天然气水合物的分解或海底的石油天然气藏中的流体沿地质薄弱带上升有关[63-64].因此,海底冷泉一般视作海底天然气水合物藏和石油天然气藏在海底的露头,冷泉区碳酸盐岩通常是由流体中的甲烷发生氧化形成的.
3.1 气源与气体类型
2007年在神狐海域钻获的实物样品显示天然气水合物分解气中甲烷体积分数为62.11%~99.91%,平均达98.04%,δ13C为-54.1×10-3~62.2×10-3,n(CH4)/n(C2H6) 值为130~11 995,其气体来源主要是微生物成因气,即以微生物气为主的混合气[22,65].2013年在南海东北部取得的可视天然气水合物样品的甲烷体积分数超过99%[66],甲烷来源为微生物作用还原CO2[67],也有少量的热成因气[68].通过雷怀彦课题组的前期研究发现,我国南海北部坡沉积物有机碳的含量为0.39%~1.26%,一般大于0.5%,同时沉积速率高,平均沉积速率介于0.13~0.73 m/ka[69],可为微生物产甲烷提供充足的有机质及良好的沉积环境.
南海北部气源充足,不仅有大量生物气及亚生物气,还有热成因气甚至幔源气的存在[70],同时由于构造运动,发育泥底劈等运移通道,使得深部热成因裂解气、原位生成的生物成因气,甚至幔源气随流体沿断层、裂隙等运移,为天然气水合物的形成提供了必要条件.
3.2 相关微生物及其标志化合物
天然气水合物区沉积物中的微生物丰度和甲烷的浓度变化相关[71-72].天然气水合物潜在区DS08-2014站位的优势古菌为Bathyarchaeota,甲烷代谢类群约占古菌的7.30%,主要为ANME-1类群并且随着深度的增加其占比逐渐增大[73].雷怀彦课题组在甲烷含量较高的中层检测到甲烷氧化菌(methanotrophs)和SRB 等与甲烷密切相关的细菌[74],该层位对应于SMTZ[75].根据收集的南海北部天然气水合物赋存区及潜在赋存区的微生物研究资料(表2)显示,MBG-B在大多数天然气水合物赋存以及冷泉活动海域均为优势古菌类群,该类群在秘鲁边缘以及喀斯喀特古陆边缘天然气水合物区SMTZ中也为主要的优势类群[80],但是在不含天然气水合物的表层沉积物中该类群也为优势菌群[81].因此,目前尚缺乏可靠的具有指示性的微生物群落特征来指征天然气水合物的存在.

表2 南海北部沉积物中古菌及细菌的菌群特征对比
注:*表示海底以下;MBG:marine benthic group;MCG:miscellaneous crenarchaeotic group;C3类群是泉古菌门喜高温的一类.
通过目前南海北部冷泉区及天然气水合物远景区和赋存区的相关研究(表3)发现,生物标志化合物的种类、含量以及碳同位素特征可以用来指示微生物的存在及种类,还可以说明微生物群落特征的变化[85].在沉积物中,结合环境参数,AOM生物标志化合物在SMTZ相对富集或者仅在甲烷含量高的层位被检测到,说明在检测到特征生物标志化合物的层位发生了AOM[60,82].据雷怀彦课题组在南海北部海域多个站位的研究推测,微生物的AOM作用加速了脂肪酸甲酯化过程,使得脂肪酸甲酯在相应的层位富集[86].而在冷泉碳酸盐岩中,细菌和古菌的生物标志化合物普遍都具有极低的δ13C,说明该碳酸盐岩的形成缘于AOM作用[60];其碳源均来自于微生物产的甲烷,并且极有可能是来自于深部天然气水合物分解所释放的甲烷[83].同时,不同生物标志化合物的组成,包括碳同位素的差异,可以反映嗜甲烷微生物组成的不同,进而反映其甲烷通量的大小[87-88].
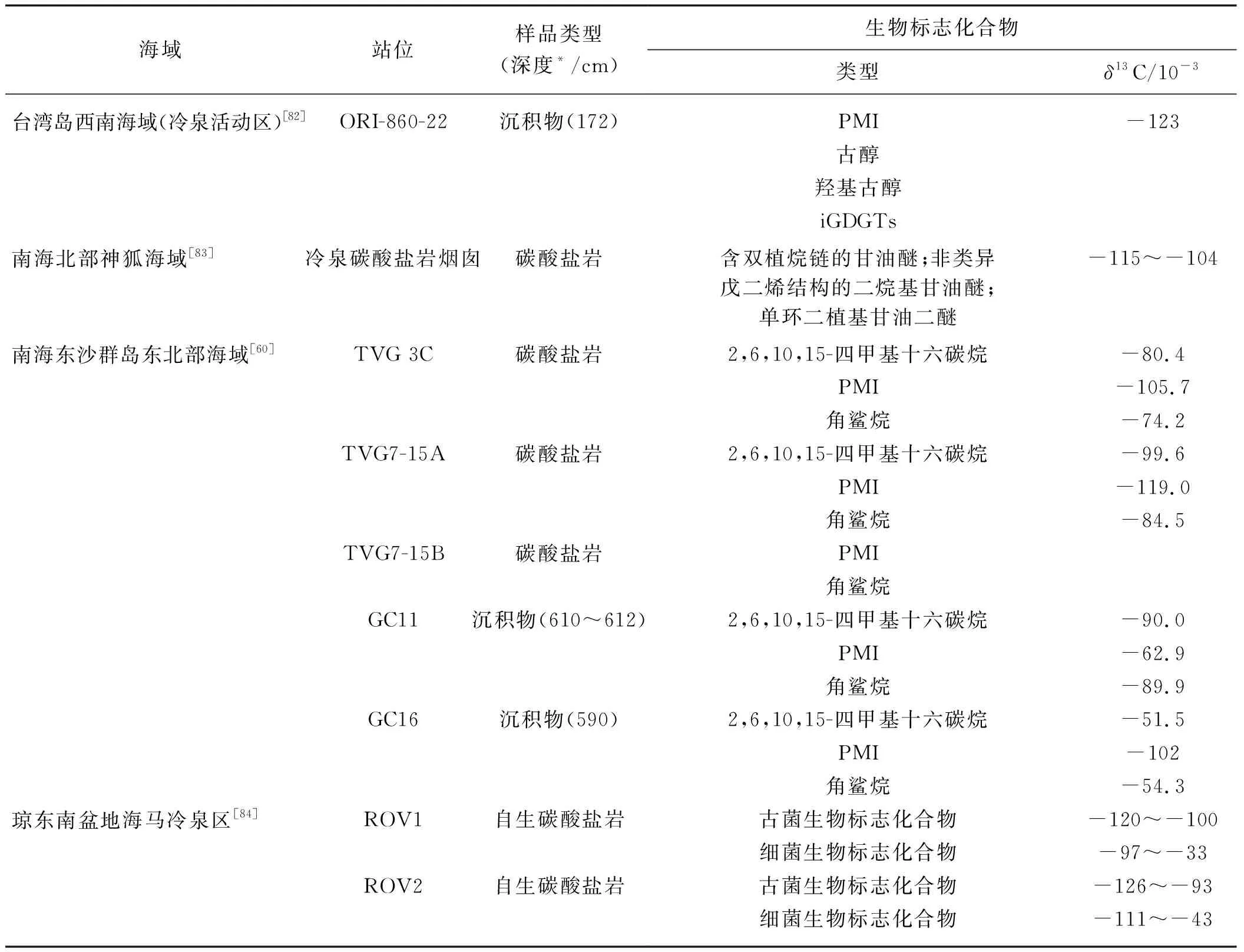
表3 南海北部沉积物中生物标志化合物对比
注:*表示海底以下;古菌生物标志化合物主要包括醚类异戊二烯类和类异戊二烯烃类;细菌生物标志化合物主要指SRB生物标志化合物,包括DGDs和脂肪酸.
4 展 望
微生物勘探技术是通过直接检测油气藏微渗漏的轻烃及烃氧化微生物从而判断油气藏的存在,该方法作为油气藏的钻井勘探前期的辅助手段,已经成功地在东海实施[89],目前通过甲烷氧化菌和丁烷氧化菌结合酸解烃的检测,成功运用于大陆冻土带的天然气水合物勘探中[90-91].在海洋沉积物中,天然气水合物的形成需要大量的生物气,该生物气是由大量的产甲烷菌富集在一定层位产生的.而天然气水合物渗漏的甲烷使得在一定深度的沉积物上检测到的特定ANME及其共生体,以及由于这些微生物聚集而产生特殊的生物标志化合物,均可以作为海洋环境中探测天然气水合物的存在与否的证据,具有成本低、操作简单、灵敏、快捷等优点,可以将其作为辅助手段勘探目标区域是否有天然气水合物的存在.
然而在目前的研究中,由于很多古菌的不可培养性,海洋沉积物中微生物与沉积地质环境的关联尚未建立完善,如Inagaki等[80]的研究中认为绿弯菌通常存在于天然气水合物较少但有机质丰富的沉积物中,含天然气水合物区发现该类群的概率远远低于不含天然气水合物的沉积物;但是在我国南海天然气水合物赋存区的微生物调查中,却发现绿弯菌为细菌文库中的优势类群[72].南海北部发现的多处天然气水合物赋存区及潜在赋存区作为研究相关微生物的天然试验场,目前大部分研究都集中在海洋沉积物的表层[81,92-93],但是稳定存在的天然气水合物的渗漏甲烷一般难以到达表层,很难影响表层微生物的群落特征,在表层沉积物样品中暂时还没有获得与天然气水合物赋存区相同或相似的群落结构特征.因此在接下来的研究中,应进一步推进现代分子生物学技术对天然气水合物赋存区不同深度沉积物的古菌和细菌的多样性调查,结合地球化学数据以及沉积环境,利用产甲烷菌以及ANME等群落及生物标志化合物特征,建立相应的微生物分布模型,以提高古菌群落结构特征对天然气水合物藏的指示性.
猜你喜欢
中国感染控制杂志(2022年11期)2022-11-24
土壤学报(2022年3期)2022-08-26
大自然探索(2022年5期)2022-07-11
知识就是力量(2022年6期)2022-06-16
化工环保(2021年5期)2021-10-19
化学与生物工程(2021年8期)2021-08-26
环境卫生工程(2020年6期)2020-12-30
江苏理工学院学报(2020年2期)2020-10-23
北京大学学报(自然科学版)(2020年3期)2020-06-07
智富时代(2019年5期)2019-07-05
