
南海水域生物固氮作用研究进展
2018-12-06 03:37李丹阳
厦门大学学报(自然科学版) 2018年6期
李丹阳,张 润
(厦门大学海洋与地球学院,福建 厦门 361102)
氮是海洋生物所必需的营养元素.海水中氮最大的储库是氮气(N2),但海洋中绝大部分的初级生产者不能直接利用海水中的氮气,因此,开阔大洋的生物生产力常受到氮营养盐的限制[1-2].少数固氮生物可以在固氮酶的作用下,将N2还原成氨氮或者有机氮,这一过程被称为生物固氮作用.通过固氮作用,固氮生物既可以满足自身的氮需求,又可将新固定的氮传递给其他生物,缓解其受到的氮限制,支持海洋新生产力[1,3-4].因此,生物固氮作用是重要的新氮来源之一,影响着海洋对大气中二氧化碳的吸收能力,进而对全球气候产生重要的影响[1,3-4].
Dugdale等[5]利用15N2示踪培养法,首次实测了马尾藻海中束毛藻(Trichodesmium)的固氮速率.随后,在一些低纬度寡营养盐海区陆续开展了生物固氮作用的研究,采样区域主要集中于亚热带北太平洋环流区(夏威夷HOTS站)及北大西洋等少数海区[3,6-7].随着认识的不断更新,海洋固氮研究已不再局限于低纬度寡营养盐开阔大洋,而是拓展至更大的空间范围,对于固氮生物组成的多样性、复杂性也有了全新的认识[8-11].了解海洋固氮作用的时空分布,对全球海洋氮收支的评估及海洋氮循环的研究具有重要意义.
南海位于热带-亚热带地区,是世界第二大边缘海.南海总面积约为3.5×106km2,平均水深1 300 m[12].南海大部分海区具有水体温暖、寡营养盐、距离大气尘降源地较近等特点,这些环境特点被认为有利于固氮作用的发生[3].自20世纪末以来,陆续开展了南海固氮生物(主要针对束毛藻)丰度、固氮速率的时空分布、固氮作用的受控因子等方面的研究[2,13-14],极大地拓展了对南海生物固氮作用的认识.
本文中主要回顾近年来南海水域生物固氮作用的研究成果,并结合全球范围内海洋固氮作用研究的最新认识,对南海固氮作用的研究提出展望.
1 南海不同区域的生物固氮作用
南海幅员辽阔,可根据理化条件的差异分为几个亚区.南海北部受黑潮水入侵的影响[2,15-16],而沿岸区域又受珠江、湄公河等淡水径流的影响[17-18].除此之外,在季风的影响下,南海常出现上升流[11,19-20],上升流为南海表层水体输送大量的营养物质,影响南海表层营养盐分布[21].其他海区的研究表明,生物固氮作用的调控机制往往具有很大的时空可变性[7].鉴于此,可将南海生物固氮作用的研究区域分为南海海盆区、南海北部陆架区、沿岸上升流区和半封闭海湾4个部分,近年来生物固氮作用研究站位分布情况如图1所示.
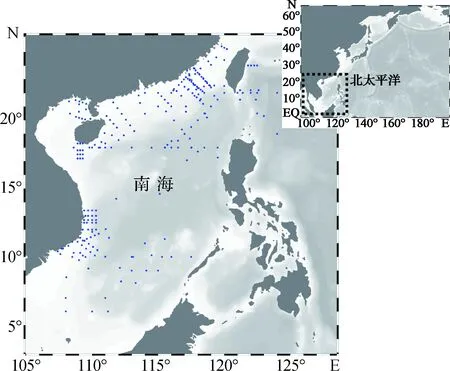
图1 近年来南海生物固氮作用研究站位分布图
1.1 南海海盆区
南海海盆区面积辽阔,约占南海总面积的1/2.目前,南海海盆区固氮作用的研究区域以北部海盆为主,特别集中于东南亚时间序列站(South East Asia time-series study station,SEATS站;位于116° E,18° N)及其邻近海区.
早期,Wong等[22]通过计算SEATS站的氮异常指数,指出SEATS站可能存在较强的生物固氮作用.氮异常指数是指在反硝化作用和固氮作用的影响下,海水中氮营养盐的过剩量,常被用来指征固氮作用的强度[23].近期,Kao等[24]在SEATS站发现了较高的氮异常指数,根据稳定同位素质量平衡模型计算,指出SEATS站的生物固氮作用引入的氮总量约为(20±26) mmol/(m2·a),占新氮总量的5%~10%.SEATS站可观测到束毛藻和胞内植生藻(Richeliaintracellularis)[21],二者在春季的平均固氮量均可达到40 μmol/(m2·d).南海生物可利用的新氮来源主要包括大气沉降、水体输送和生物固氮三方面.南海海盆区固氮生物组成复杂[25-26],若仅考虑束毛藻和胞内植生藻的固氮作用,SEATS站中3种氮来源所提供的氮总量所支持的生产力仅占该区域群落净生产力(4.47 mmol/(m2·d))的55%[27].这一方面揭示了再生氮对SEATS站群落净生产力的支持,另一方面揭示了其他类型固氮生物的重要贡献.若以SEATS站的生物固氮量为参考,将此调查结果外推到整个南海海盆(总面积≈1.8×106km2),南海海盆每年通过生物固氮作用收入的氮总量可达3.6×107mol.
尽管束毛藻长期被认为是全球海洋最重要的固氮生物[3],但在南海很少观察到束毛藻的高丰度.近年来,单细胞固氮生物被发现广泛分布于全球海洋中[9-10,28],在南海也可检测到单细胞固氮生物的高丰度.Zhang等[25]于2011年报道了南海海盆区固氮生物的组成及丰度,指出南海海盆区的固氮生物以变形菌为主,其中α-变形菌和γ-变形菌的丰度最高(约为104~105L-1),相比之下,丝状蓝藻(如束毛藻)的丰度较低.单细胞固氮生物在南海广泛分布,而不仅局限于海盆区[15-16,26],对南海的生物固氮作用可能有重要贡献.
1.2 南海北部陆架区
南海北部广阔的陆架区也是生物固氮作用研究的重要场所.黑潮入侵、陆地径流等因素都可能对此处的固氮作用产生影响.
黑潮水自吕宋海峡进入南海,是影响南海北部陆架区固氮作用的一个重要因素.虽然具体原因不明,但是黑潮上游往往具有高固氮生物丰度[15,29-30].据报道,黑潮水中无论是束毛藻的平均丰度,还是水体实测固氮速率均比邻近南海海区高出近1个数量级[15,31],随着黑潮水的入侵,一定丰度的固氮生物也可能被携带进入南海.近年来的研究结果进一步证明,黑潮上游区的固氮生物群落也具有较高的多样性,单细胞固氮生物是不可忽视的固氮贡献者[15].Wu等[16]在2018年的报道中指出了黑潮及南海中单细胞固氮生物的重要性,报道指出:单细胞固氮生物的固氮速率变化范围为0.63~7.61 nmol/(L·d),均值可达3.67 nmol/(L·d),单细胞固氮生物贡献了采样区域内近75%的固氮通量.
台湾海峡南部也是固氮作用较为活跃的海区,林峰[32]利用15N2示踪法测定了台湾海峡南部海域的生物固氮速率,结果表明夏季台湾海峡南部海域的生物固氮速率介于0.168~1.08 nmol/(L·d)之间,平均为0.537 nmol/(L·d),其中次表层的固氮速率更高.Chen等[15]指出,次表层较强的生物固氮作用是由单细胞固氮生物所贡献.
受陆地径流的影响,珠江口附近海域结合态氮营养盐较为丰富,此处也是生物固氮作用的一个热点区域.刘甲星等[33]报道了2014年秋季珠江口附近海域具有较高的固氮速率,表层水的固氮速率均值高达(0.77±0.54) nmol/(L·d).较高的无机氮营养盐浓度并未抑制固氮作用的发生,这和该区域的固氮生物组成有着较大的关系.Kong等[18]发现珠江口附近海域固氮生物组成较为多样,包括异养型变形菌、光合蓝藻、Cluster Ⅲ类固氮生物等,而束毛藻所占比例不高,盐度可能在一定程度上调控着固氮生物的组成.
1.3 沿岸上升流区
沿岸上升流是南海西侧的一个典型现象.沿岸上升流区的固氮作用在很长时间内被忽视了,这是由于上升流水体具有的典型理化条件(较低的水温、较为充足的结合态氮营养盐等)与固氮作用的传统观点不符.但是,近年来越来越多的实测研究证据表明,沿岸上升流区并非固氮作用的禁区;相反地,热带-亚热带沿岸上升流区(包括赤道上升流区)往往是固氮作用发生的热点,已有研究报道了南海越南沿岸上升流区、琼东上升流区及台湾海峡南部上升流区的固氮作用[11,19-20].
目前,在热带越南沿岸上升流区域开展的生物固氮作用的研究较多.在非季风时期,越南沿岸附近以寡营养盐为特征,而在季风期,受上升流影响,越南沿岸的营养盐含量明显提升,除上升流外,湄公河径流也是越南沿岸海域中重要的营养物质来源[34-36].湄公河河水每年向南海输入大量淡水,使越南沿岸海域出现明显的盐度梯度,同时携带大量陆源的颗粒物质及微量元素进入南海[17,36].在越南沿岸上升流区,西南季风盛行期固氮速率较非季风期明显升高,这表明湄公河羽状流可能在很大程度上调控着该区域固氮速率及固氮生物的空间分布[11,37].目前具体作用机制不明,推测可能的原因包括河流淡水对水体垂向稳定度的提升、痕量元素的输入、硅营养盐输入促进硅藻-固氮藻共生体的生长等方面[11,17-18].
Moisander等[13]检测了越南沿岸流域中固氮生物的组成和丰度,结果表明:调查区域内的固氮生物主要由束毛藻和α-变形菌组成,除此之外,单细胞的固氮蓝藻和共生型蓝藻也广泛存在.Grosse等[17]研究发现单细胞固氮生物多出现在湄公河河口附近,束毛藻仅在盐度较高的(盐度>33.5)区域中出现,而与硅藻共生的固氮生物则是广泛分布于调查区域内.由此可见,湄公河径流影响着越南沿岸流域中固氮生物的分布.Voss等[11]检测了越南沿岸上升流区域内的生物固氮速率,结果表明:当西南季风盛行时,调查区域内的固氮速率变化范围为21.4~190.6 μmol/(m2·d);非季风期时,固氮速率的变化范围为1.9~36.0 μmol/(m2·d).相比之下,季风盛行时,固氮作用要明显强于非季风期,这可能是由于湄公河径流的影响造成的[11,38].来自湄公河的淡水一方面可以加强水体的稳定度,另一方面可携带大量微量元素进入南海,从而促进固氮作用[39].除微量元素外,湄公河向南海输送了大量的硅,这也可能通过促进硅藻的生长而对共生型固氮生物产生间接的影响[11,40].因此,越南沿岸上升流区域内的固氮作用可能受上升流和湄公河径流的共同影响.整体而言,在越南沿岸上升流区,丝状蓝藻和单细胞蓝藻对固氮的贡献量相当,固氮作用对初级生产力的贡献率的变化范围为1%~47%[17],是不容忽视的氮来源.
琼东上升流是亚热带南海北部一个典型的沿岸上升流区.Zhang等[20]报道了2013年夏季海南岛东侧上升流区域的生物固氮作用,结果表明:该区域表层固氮速率的变化范围为0.1~5.6 nmol/(L·d)(均值为1.0 nmol/(L·d)),固氮速率从沿岸区域到开阔海域有增强的趋势,其对初级生产力的贡献率变化范围为0.01%~2.52%.若将琼东上升流区的调查结果外推到南海西北陆架区(总面积≈33×104km2,17° N以北),假设夏季时长为90 d,生物固氮作用引入氮的总量约为1.4 ×109mol[20].该报道提出了一个固氮作用受控的物理-生物耦合机制,即:上升流向表层输送低温、富营养盐且铁含量丰富的深层海水,促进固氮生物增长,而非固氮生物对磷和铁的吸收可能会抑制生物固氮作用,因此,随离岸距离的增加,初级生产力减小,生物固氮作用增强[20],这一机制对于亚热带沿岸上升流区的固氮作用调控可能具有普遍性.Wen等[19]最新的调查研究中报道了台湾海峡附近上升流区域的固氮作用,结果表明:调查区域内固氮作用强烈,最大固氮速率可达7.51 nmol/(L·d),在氮磷摩尔比较低的区域中生物固氮速率更高.
1.4 半封闭海湾
南海存在诸多半封闭海湾,如北部湾、大亚湾等,这些海区往往具有迥异于开阔水体的特点,也是潜在的固氮热点区域.Dong等[41]研究发现铁氏束毛藻(Trichodesmiumthiebautii)是三亚湾内固氮生物的优势种,已成为三亚湾的一个重要氮源.林峰[32]的研究发现,北部湾固氮速率存在南高北低的特点,受温度、无机营养盐等理化因素的影响.固氮作用可支持北部湾近2%初级生产力的氮需求[32].束毛藻曾经是大亚湾的赤潮种[42],但是近年来束毛藻水华未见报道,可能暗示在日益加剧的人类活动压力下,该区域固氮生物组成在过去数十年间可能发生了显著的变化.已有研究显示,大亚湾生态环境正在发生深刻的变化[43],对于其他南海半封闭海湾亦然.
表1给出了部分研究所报道的南海各区域生物固氮速率.南海生物固氮速率的变化范围为0~8.5 nmol/(L·d).相对而言,上升流区域的固氮作用更为活跃,在海盆区也可观测到活跃的固氮作用,而受淡水径流影响的珠江口和三亚湾固氮速率则相对较低.由于调查季节不同,采用方法也有差异,结果之间的可比性较差.除此之外,各区域固氮生物的组成有所差异,且会随季节发生变化.在受黑潮影响的北部陆架区及越南沿岸区域,束毛藻是固氮生物的重要组分[13,15,31],而在珠江口附近海域和开阔南海海盆区,束毛藻的丰度非常小,固氮生物主要由单细胞蓝藻组成,同时存在大量的固氮变形菌[18,25,33].不同类型固氮生物的固氮能力不同,这也是造成各区域生物固氮速率差异的重要原因.整体而言,南海生物固氮速率的报道值分布在北大西洋报道值的低值范围内[1,7],与北太平洋的报道值处于相同量级[3].

表1 南海固氮速率的研究报道
2 南海生物固氮作用的研究方向
2.1 拓展调查范围
目前,在全球范围内,近岸海域中生物固氮的研究正逐渐增加[44-46].沿岸海域理化环境复杂,受人类活动影响很大,固氮作用的调控机制可能迥异于开阔水体.这些研究表明沿岸海域中也可观测到较强的固氮作用.沿岸海域理化环境复杂,既受陆源物质的影响,又可能受人类活动的影响,生物量及生物多样性相对更高,有更大的研究空间.然而,南海中沿岸海域中生物固氮作用的相关报道目前仍较少,有待补充.
在过去20年,已有报道证明,南海中的固氮生物除束毛藻外还包括共生型蓝藻(胞内植生藻)、单细胞蓝藻、固氮变形菌等[13,25],其中单细胞固氮生物的生物量及固氮量甚至可以超过束毛藻等丝状蓝藻[15-16,26].事实上,许多非蓝藻类固氮生物也广泛分布于全球海洋[8-10].一些异养型的固氮生物,由于不受光的限制,具有更广阔的分布范围[9].已有报道指出:在无光层也可以观测到异养固氮生物的固氮活动,引入的氮可占调查海域总固氮量的50%以上[47].目前,南海中生物固氮作用的研究仍多局限在真光层内.后续研究可将南海生物固氮作用的调查范围由真光层拓展到弱光层.
2.2 探索调控机制
生物固氮作用受多种因素共同调控[1,3,48].南海生物固氮作用的研究多数关注无机营养盐和铁对固氮作用的调控[11,15,20,36].事实上,除无机营养盐和铁的含量之外,还有诸多因素可能调控固氮作用的分布.
近年来,有机物与生物固氮作用之间的关系被广泛研究,已有许多报道指出:海水中的溶解有机质可能会刺激生物固氮作用[47-49],甚至自养型的束毛藻也会吸收水体中的溶解态有机质[50].生物固氮是一个极其耗能的过程,有机质可能是固氮生物的一个重要的能量来源.此外,铁是固氮酶合成与表达所必须的元素.海水中有机质的存在可能为生物提供了有效的铁的有机配体,很大程度上提升了铁的生物可利用性,缓解了生物固氮过程受到的铁限制[2,51].南海受湄公河、珠江等淡水的径流影响[17,33],淡水径流携带大量陆源有机质入海,除此之外,沿岸的海湾由于受人类活动的影响,也可能向南海中输入大量的有机物质,因此,未来可在南海深入研究有机质对固氮作用的调控机制.
在气候变化的大背景下,海洋酸化对南海固氮作用的影响可能亦不能小觑[14].固氮作用同全球气候密切相关,因此在这方面的研究需进一步深化.由于生物固氮作用是在多种因素的共同调控之下进行的,今后的研究可更多关注多种因子之间的协同作用.
2.3 分析固氮与固碳过程的耦合
固氮作用与固碳作用之间可能存在复杂的联系.非固氮生物可利用固氮生物所提供的氮来满足自身需求[1,3].同时,在其他营养物质方面,非固氮生物同固氮生物之间存在竞争作用,从而间接地调控着固氮过程[2,20].除了营养物质的竞争外,非固氮生物可能会通过释放固氮生物可利用的溶解态有机质来间接地影响生物固氮过程:海洋中存在大量的溶解态有机质,但其中生物可利用的仅占很小一部分[52],而浮游植物在光合作用后会释放溶解态有机质到水体中[48,53],这可能为固氮过程提供了重要的能量来源[48,50].因此,海洋生物固氮作用与固碳过程之间存在密切的关联.在南海,固氮作用与固碳作用二者的关系如何?是否存在初级生产者对固氮生物的调控?需要进一步研究加以回答.
为进一步探索南海生物固氮过程与固碳过程之间的耦合关系,首先可在实验室进行培养实验,通过结合细胞计数、稳定同位素分析等技术,探索碳和氮在固氮生物和非固氮生物之间的传递,以期获得可应用于实际海洋的结论.
2.4 规范研究方法
生物固氮速率的测定方法主要包括乙炔还原法和15N 示踪法[54].15N示踪法直到20世纪90年代才开始广泛应用[55].15N示踪法通过向密闭体系中加入15N2标记气体,经一段时间的培养后,测定该体系中固氮生物对15N2的吸收情况来测定固氮速率[55].南海中生物固氮速率的研究,在测定生物固氮速率方面使用的方法不一,一些研究中使用的方法可能会使结果偏离真值,且各项研究之间的可比性较差.在利用15N示踪法测定生物固氮速率时,培养时间的不同也会造成测定结果的差异[16,56].今后,有必要对南海中生物固氮作用的调查方法进行规范与统一,一方面可以对南海中各区域生物固氮活动进行对比,另一方面可以真实地评估南海的氮收支情况,这对南海氮循环及碳循环而言具有重要意义.
3 总结与展望
南海是研究生物固氮作用的理想场所.南海固氮生物组成较为复杂,具有较大的时空可变性,主要包括束毛藻、胞内植生藻、单细胞蓝藻和固氮变形菌等,其中单细胞固氮生物对南海生物固氮的贡献不容忽视.南海的固氮速率与其他热带、亚热带海区相近,其中海盆区的年固氮量可达3.6×107mol/a.整体而言,生物固氮作用对南海初级生产力的贡献率较低.南海生物固氮作用受黑潮水入侵、陆地径流的影响,除此之外,上升流为南海表层水体输送大量的营养物质,从而间接调控着生物固氮作用.本文中结合生物固氮作用在全球范围内的研究现状,指出今后需进一步拓展南海中生物固氮作用的调查范围,并对其调控因素以及其与海洋生物固碳过程之间的耦合进行更全面的分析,同时对生物固氮的调查方法进行规范与统一,进而为研究南海中的海洋氮循环提供准确的信息.
猜你喜欢
环境工程技术学报(2022年3期)2022-06-05
小猕猴智力画刊(2022年4期)2022-05-23
农业科技与信息(2021年16期)2021-12-07
生态科学(2021年5期)2021-11-13
黄河之声(2021年10期)2021-09-18
科学与技术(2019年21期)2019-04-27
军营文化天地(2017年1期)2017-03-06
草原(2016年1期)2016-01-31
应用海洋学学报(2015年3期)2015-11-22
应用海洋学学报(2015年1期)2015-11-22
