
植物内生细菌修复重金属污染土壤作用机制研究进展
2018-11-30 03:08张萌萌曹立东王仁卿孙瑞莲
生物技术通报 2018年11期
张萌萌 曹立东 王仁卿 孙瑞莲
(1. 山东大学生命科学学院,青岛 266237;2. 山东大学环境研究院,青岛 266237)
近年来,植物内生细菌在土壤重金属修复领域的研究受到广泛关注,尤其是内生细菌和植物相互作用方面。植物内生细菌作为植物微生态系统中的天然组成成分,展现出高代谢能力和代谢活性特异性,对加强植物对环境的适应性,改善植物生长,提高植物的逆境耐受性和环境可持续性具有重要意义。内生细菌具有溶磷、产生植物激素、合成铁载体、诱导植物产生抗性等功能,能提高植物修复重金属污染土壤的效果。内生细菌联合植物修复策略包括内生细菌如何加强植物对重金属的抗性、促进植物生长以及植物对重金属的富集等方面,具有重要的理论和实际研究意义。目前,有关内生细菌联合植物修复重金属污染土壤方面已有一些研究,本文对近几年来国内外内生细菌协同植物修复重金属污染土壤的作用机理进行了总结,以期推动内生细菌在重金属污染土壤修复中的应用。
1 植物内生细菌强化植物修复重金属污染土壤的机制研究
全面了解耐重金属内生细菌的特性,是提高重金属污染土壤植物修复效率的重要因素[1]。了解内生细菌的特性,通常进行革兰氏染色、氢氧化钾(KOH)试验、吲哚试验、甲基红试验、氧化酶试验、柠檬酸盐试验、淀粉水解试验、蛋白酶生产试验、明胶液化试验和嗜铁细胞生产等试验。内生细菌作为植株整个微生态系统的重要组成部分,具有强化植物修复重金属污染土壤的功能(表1),主要表现在以下几个方面:(1)通过产生植物生长调节激素、产生1-氨基环丙烷-1-羧酸(1-Aminocyclopropanecarboxylic acid,ACC)脱氨酶等,促进宿主植物种子萌芽、分蘖成长、幼苗存活和生物量累积等,进一步促使植物吸收和累积重金属;(2)通过改变重金属的生物有效性/毒性,增强植物向地上部转运重金属的能力,或者某些具有生物吸附/富集能力的内生细菌,降低重金属的生物有效性,使植物免受重金属毒害;(3)通过与植物形成联合修复体,提高植物抗重金属毒性的能力。
1.1 植物内生细菌的促生作用
植物激素有助于植物组织的生长和分化,调节植物的生长发育,过多或过少都会导致植物生长发育异常。植物内生细菌通过自身合成或者促进植物合成生长素吲哚乙酸(Indole-3-acetic acid,IAA)、赤霉素(Gibberellin,GA)和细胞分裂素(Cytokinin,CTKs)等植物激素,促进植物体的生长发育,其中IAA是最活跃的成分[10]。接种荧光假单胞菌(Pseudomonas fluorescens)的植物对铅具有抗性,其原因在于荧光假单胞菌产生吲哚乙酸并作用于植物体[11]。王璐等[12]从Cu耐性富集植物苏丹草根中分离得到了根瘤菌属K1-6和肠杆菌属K3-9,均具有产生IAA的能力,产量为21.8-54.9 mg/L,在添加Cu2+的条件下,接种菌株K1-6处理的苏丹草根部和地上部干重分别比对照增加27%和13%,接种菌株K3-9处理的苏丹草根部和地上部干重分别比对照增加41%和37%。关于内生细菌产生GA的研究很少,更多的产生GA的内生菌属于内生真菌,它们与植物的生长关系密切,能减轻环境胁迫[13]。
重金属胁迫会诱导植物产生大量乙烯,导致根的生长及根毛的形成受到抑制,阻碍植物生长,甚至导致植物死亡。许多内生细菌能分泌ACC脱氨酶,该酶是一种将ACC(乙烯的前体)水解成α-酮丁酸和氨的酶,能有效降低金属胁迫条件下植物乙烯合成量,从而减少过量乙烯对植物生长造成的不利影响,打破乙烯的抑制作用,提高植物抗逆性,此外,氨还可作为氮源,被植物吸收利用[14]。ACC脱氨酶是内生细菌操控宿主植物的重要手段,Sun等[15]发现,从超富集植物油菜体内分离得到的ACC脱氨酶活性较高的Cu耐受型内生细菌巨大芽孢杆菌JL35和鞘氨醇单胞菌YM22,能显著提高植物的生物量和Cu吸收量。Zhang等[16]报道,在Pb存在的情况下,接种具有较高ACC脱氨酶活性的不动杆菌Q2BJ2和芽孢杆菌Q2BG1,增加了油菜地上部分(15%)和根部(23%)的干重。
内生细菌是植物病害生物防治的天然资源菌,在植物体内有稳定的生存空间,并且能产生与宿主植物代谢相同或相似的生理活性物质,能够有效地提高植物的抗病性。几丁质酶能促进真菌细胞壁几丁质的水解,抑制真菌的生长增殖,提高植物抗真菌能力,从而促进植物生长[17]。内生细菌Serratia marcescensMOSEL-w2和Paenibacillussp. MOSEL-w13能分泌几丁质酶,具有抗病原真菌能力,其中,类芽孢杆菌属MOSEL-w13显示出较大的抗真菌活性,能够阻碍超富集植物欧洲油菜(Brassica napus)和大麻(Cannabis sativa)病原真菌黑曲霉和尖孢镰刀菌的生长[18]。植物内生细菌通过合成胞外酶如几丁质酶,降解植物病原真菌的细胞壁,抑制植物病害,有效促进了植物生长发育。
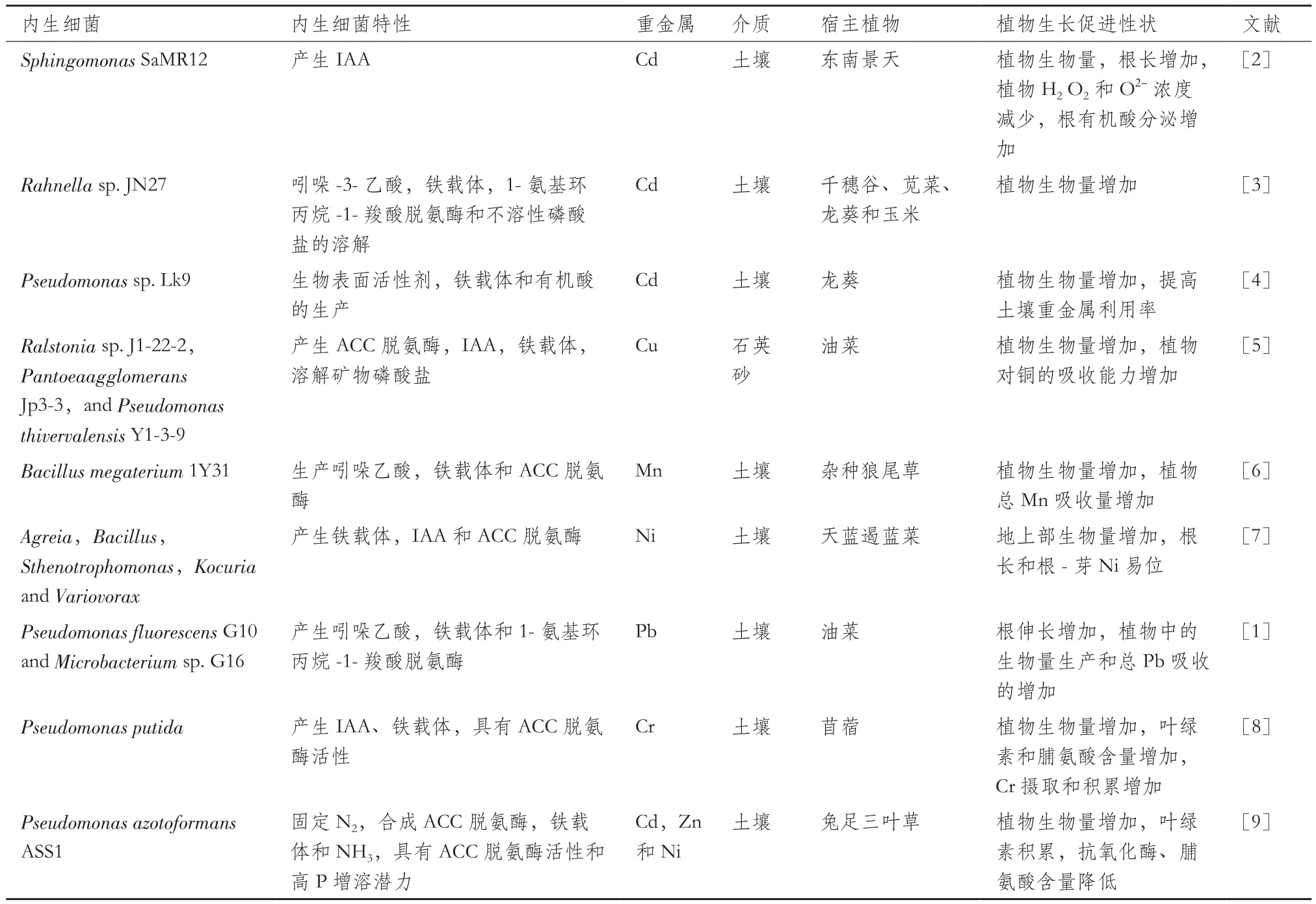
表1 金属胁迫下内生细菌的特性及对植物修复重金属污染土壤的影响
有些内生细菌同时具备多种促生功能,可以在植物不同生长阶段或者不同生长状态下,促进植物的生长发育,调控植物的生理过程。Zhang等[5]从超富集植物美洲商陆(Phytolacca Americana)的根、茎和叶中分离出了耐铜内生细菌,其中,Ralstoniasp. J1-22-2、Pantoea agglomeransJp3-3 和Pseudomonas thivervalensisY1-3-9具有多种植物生长促进特征,表现较高的ACC脱氨酶活性,能产生铁载体和吲哚乙酸,并可以溶解无机磷酸盐,其接种显著促进欧洲油菜(Brassica napus)生长。耐镉内生细菌Rahnellasp. JN27分离自玉米(Zea mays)组织,能分泌吲哚乙酸和铁载体,具有ACC脱氨酶活性和溶磷能力,其接种使千穗谷(Amaranthus hypochondriacus)、 苋 菜(Amaranthus mangostanus)、龙葵(Solanum nigrum)和玉米地上和地下部分的生物量都有显著增加[3]。
1.2 内生细菌改变重金属的生物有效性/毒性
许多内生细菌具有分泌铁载体、有机酸等物质的能力,这些物质可以与重金属形成络合物,或者活化土壤中的重金属,从而改变重金属的生物有效性,减轻重金属对植物体的毒害作用。铁载体是一种对Fe3+具有超强络合力的低分子量有机化合物,不仅能够与铁结合,还可以与Al、Zn、Cu、Pb和Cd等金属形成稳定的复合物,改变重金属的生物有效性[19]。在内生细菌产生的铁载体作用下,许多植物会消耗土壤中的微量可利用铁,获得生长所需的铁元素[20]。内生细菌能产生有机酸,造成环境酸化,使土壤中的重金属得到活化,在一定程度上,提高了土壤重金属的生物有效性。邓平香等[21]研究表明,荧光假单胞菌R1在生长代谢过程中,能分泌苹果酸、琥珀酸、乙酸、柠檬酸、草酸等有机酸,导致ZnO和CdO溶解,提高了重金属的生物有效性,在一定程度上促进东南景天的生长,增加其对Zn和Cd吸收。从Cd超富集植物龙葵中分离得到的Pseudomonassp. Lk9,能分泌较多的乙酸和柠檬酸等有机酸,促使土壤中有效Cd、Zn和Cu的释放,其接种显著提高了Cd的生物富集因子(28.9%)和其他金属的植物提取率(Cd、Zn和Cu分别为17.4%、48.6%和104.6%),其中,土壤重金属水溶性组分的增加主要归因于酸萃取组分的流动性[4]。
某些内生细菌具有生物吸附/富集能力,能通过释放螯合剂等方式,降低重金属的生物有效性,使植物免受重金属的毒害。从球葵属植物Sphaeralcea angustifolia和牧豆树(Prosopis laevigata)根中分离出内生细菌,约70%的内生细菌可产生用于螯合As5+的化合物,而44%的内生细菌可合成用于螯合 As3+的化合物[22]。Shin 等[23]研究表明,耐铅内生芽孢杆菌MN3-4能通过细胞外螯合和细胞内积累铅,减轻重金属对赤杨皮的危害。赵希俊等[24]报道洋葱伯克霍尔德菌G3具有良好的铝离子吸附能力,并且吸附效果主要受吸附时间与pH的影响,受温度变化的影响较小。在单金属或多金属系统中,内生细菌能够选择性地吸附结合重金属。从龙葵(Solanum nigrum)中分离得到内生细菌沙雷氏菌LRE07,在72 h内,对Cd2+和Zn2+的吸附量分别可达65%和36%,而对Cu、Pb和Cr的吸附较少[25]。
内生细菌通过直接氧化或还原作用,改变重金属的形态,降低重金属的毒性,从而达到解毒的效果。众多重金属在土壤中以多种形态存在,而不同形态其毒性也不一样。例如,砷(As)在土壤和水中的主要形式是无机亚砷酸盐[As(III)]和砷酸盐[As(V)],含氧条件下,As(V)占主导地位;相反,在还原条件下,砷主要以As(III)形式存在,其比As(V)更具流动性和毒性[26-28]。从牧豆树(Prosopis laevigata)和球葵属植物Sphaeralcea angustifolia根中分离出的内生细菌,具有As5+还原和As3+氧化的双重能力,可改变As的存在形态。其中,在有氧条件下,Micrococcus luteusNE2E1可减少 94% 的 As5+,Pseudomonas zhaodongensisNM2E7可氧化 46% 的 As3+[22]。Wang等[29]从超富集植物蜈蚣草(Pteris vittata)分离得到的内生细菌Serratia marcescensPRE01,具有较高的V、Cr抗性,PRE01在72 h内对Cd(II)、Cr(VI)和V(V)的去除率分别为97.6%、21.7%和6.6%,大多数V(V)和Cr(VI)通过菌株还原成较低的价态来缓解金属毒性。
1.3 内生细菌能提高植物重金属抗性
内生细菌与植物协同进化,在重金属胁迫下,内生细菌往往可以耐受多种重金属污染,在一定程度上,提高了植物的重金属抗性。大量研究表明,内生细菌可帮助宿主植物适应成长环境,提高植物修复效率[30-31]。内生细菌能调节宿主植物的抗氧化酶活性,植物在受到金属胁迫后,造成活性氧(Reactive oxygen species,ROS)大量累积,如过氧化氢(H2O2)等,这给植物造成很大危害,包括伤害细胞膜的完整性、体内各种酶的活性、光合作用相关的细胞器等[32-34]。植物体内的超氧化物歧化酶(Superoxide dismutase,SOD)、过氧化氢酶(Catalase,CAT)、过氧化物酶(Peroxidase,POD)、抗坏血酸过氧化物酶(Ascorbate peroxidase,APX)、谷胱甘肽还原酶(Glutathione reductase,GR)和谷胱甘肽过氧化物酶(Glutathione peroxidase,GPX)等抗氧化酶能有效减轻对植物的这些危害[35]。接种内生细菌能使宿主植物各种抗氧化酶活性增强,H2O2等降低,提高了植株对有害金属的耐受性。Wan等[36]研究表明,在 10 μmol/L 和 50 μmol/L Cd 条件下,接种粘质沙雷氏菌LRE07的龙葵植物叶片中的SOD、APX和GPX活性显著高于非接菌组,有效抵御了重金属引起的氧化胁迫,减轻了重金属对植物的毒害程度。
1.4 影响内生细菌修复重金属污染土壤的因素
内生细菌具有优异的适应能力和良好的修复效果,有助于减少植物重金属毒性效应,增加重金属的吸收和去除速率,许多因素会影响内生细菌在重金属植物修复中的作用。首先,重金属浓度是一个重要因素,在重金属浓度较低的情况下,内生细菌产生的有利影响更为明显[11]。
内生细菌的来源影响其在土壤重金属修复中的效果。内生细菌在许多植物物种均有发现。其中,超积累植物在土壤重金属修复中具有广阔的应用价值,从超积累植物中分离得到的内生细菌,多具有重金属抗性的特点。例如,从天蓝遏蓝菜(Noccaea caerulescens)中分离得到的可培养内生细菌,在培养基中表现出对Ni不同程度的耐受性[7];从东南景天分离得到的内生细菌SphingomonasSaMR12能增加植物体内Cd的积累量,并且提高植物吸收Cd的能力[2];从蜈蚣草中分离得到的内生细菌Serratia marcescensPRE01具有较高的V、Cr和Cd抗性,能改变重金属形态,进一步加速内生细菌和植物之间的长期相互作用,促进宿主对金属的吸收和运输[30]。从伏毛蓼(Polygonumpubescens)分离出来的Rahnellasp. JN6对重金属(特别是Cd、Pb和Zn)表现出非常高的耐受性,这可能与伏毛蓼在多种金属污染土壤中生长有关[37]。
内生细菌的活性对植物生长和重金属 吸收也会产生影响。例如,与油菜植物接种死菌相比,当接种G10和G16活菌株时,根和地上部干重显著增加,进一步促进重金属污染土壤修复[9]。另外,接种单菌和多菌效果不同,刘莉华等[38]报道,抗性菌株能显著促进龙葵在镉污染土壤上的生长,强化龙葵吸收土壤中镉的能力,不同镉抗性菌株及菌株组合对龙葵各部位吸收镉的影响不同,而且,混菌组合效果要明显优于接种单菌。
此外,环境胁迫也能影响内生细菌在土壤重金属修复中的效果。各种生物因素和非生物因素会影响内生细菌的定殖及其在土壤重金属修复中的效果,如干旱和土壤微生物等因素。在干旱胁迫条件下,与未接种相比,接种产氮假单胞菌Pseudomonas azotoformansASS1显著提高了金属(Cu、Zn和Ni)的积累量、总去除量、生物富集因子和生物积累系数,同时也降低了Cu的易位因子,有助于兔足三叶草(Trifolium arvense)在半干旱环境中繁殖,并提高重金属污染土壤的修复过程[39]。邓平香等[21]在研究产酸内生菌荧光假单胞菌R1对东南景天生长和吸收重金属的影响中发现,土壤灭菌处理更有利于东南景天的生长,其根系和地上部的生物量显著增加,而且土壤中可提取态Zn、Cd含量也显著增加。
2 超富集植物内生细菌多样性
植物内生细菌是从表面消毒的植物组织中分离得到或从植物内部获得的,能够定殖在健康植物的各种组织和器官内,并未使植物的表型特征和功能发生改变的细菌[40]。第一株内生细菌是Pasteur等[41]于1876年从无菌葡萄果汁中提取出的。内生细菌在植物体内广泛存在,绝大多数高等植物体内都存在着数量庞大的内生细菌。据不完全统计,从多种野生植物、农作物及经济作物中分离到的内生细菌种类已超过了745种,鉴定为54个属,其中假单胞菌属(Pseudomonas)数量最多,居优势地位,约占42%;其次为芽孢杆菌属(Bacillus)和黄单胞菌属(Xanthomonas),其他还有伯克霍尔德氏菌属(Burkholderia)等[42]。在目前的植物-内生细菌联合修复研究中,植物多采用超富集植物或富集植物,了解超富集植物内生细菌多样性,对于明晰内生细菌群落结构组成、发现功能菌株等具有重要意义。
目前,研究植物内生细菌多样性的方法主要有培养法和免培养法。相当多的内生细菌处于存活但却不可培养状态,培养法难以模拟自然界中内生细菌生长的真实环境。随着分子生物学技术的发展,基于16S rRNA基因的指纹图谱技术如变性梯度凝胶电泳(Denatured gradient gel electrophoresis,DGGE)、温度梯度凝胶电泳(Temperature gradient gel electrophoresis,TGGE)、单链构象多态性检测(Single strand conformation polymorphism,SSCP)、核糖体DNA扩增片段限制性内切酶分析(Ampli fi ed ribosomal DNA restriction analysis,ARDRA)和末端限制性片段长度多态性指纹技术(Terminal-restriction fragment length polymorphism,T-RFLP)等被广泛用于微生物群落的研究中[43],克服了传统培养法的不足,尤其是近年来发展迅速的高通量测序技术,为检测植物内生细菌的多样性提供了更为有效的手段。Sun等[15]基于16S rDNA序列的系统发育分析表明,从耐铜植株海州香薷(Elsholtziasplendens)和鸭趾草(Commelinacommunis)中获得的耐铜菌株属于三大系统发育类群:厚壁菌、放线菌和变形杆菌。邓平香等[44]利用Illumina Miseq高通量测序技术,分析了超积累生态型东南景天茎、叶内生细菌的群落结构,在属分类水平上,植物叶片和茎的第一优势菌属分别为聚球藻属(Synechococcus)和邻单胞菌属(Plesiomonas)。
此外,内生细菌的数量、种类和群落结构受多种因素的影响。目前,已从大量植物体中分离获得内生细菌[45-48],不同植物体内含有的内生细菌种类和数量有明显差异,同一植物体内不同器官之间含有的内生细菌种类和数量也显著不同。根据形态学特征,从Cd超富集植物龙葵组织中共分离得到30株耐重金属的可培养内生细菌,内生细菌在根中最多,并且从茎到叶逐渐减少[49]。植物内部组织为内生细菌提供了相对独立的生存环境,但外在环境因素会影响内生细菌群落结构。其中,重金属胁迫是影响内生细菌数量、群落结构和活性的重要因素之一[50]。Zhang等[6]发现,从植株根、茎和叶分离得到的内生细菌具有产IAA的特性,可诱导杂种狼尾草(Pennisetum americanum×P. purpureum)对重金属解毒并产生抗性,并且锰胁迫会提高产IAA内生细菌的比例。在重金属胁迫下,从耐重金属植物体内分离到的内生细菌通常具有对重金属吸收、耐受和解毒的特性。从超富集植物东南景天(Sedum alfredii)中分离得到伯克氏菌属SaZR4、SaMR10,鞘氨醇单胞菌属SaMR12和贪噬菌属SaNR1,其中,伯克氏菌属SaZR4显著促进植物对Zn的提取,鞘氨醇单胞菌属SaMR12和贪噬菌属SaNR1显著促进植物对Zn和Cd的提取[51]。
3 展望
内生细菌作为一种天然资源,是辅助植物修复重金属污染土壤的重要工具,具有广阔的理论研究价值和开发应用前景。内生细菌-植物联合修复重金属污染土壤的相关研究已经取得了重大进展,随着研究的深入,有望探讨更深层次的机理,并使大规模的实际应用成为可能,但目前的研究还主要局限在植物生理代谢水平上。为了更好地了解内生细菌强化植物修复重金属污染土壤的作用机制,推动内生细菌大规模应用于植物修复重金属污染土壤,今后的研究重点应考虑:(1)内生细菌自身存活原因以及如何耐受重金属的机制研究;(2)分子生物学角度研究植物内生细菌的行为动力学和代谢;(3)在逆境条件下,接种外源内生细菌的生态学效应研究,包括植物代谢和微生物的群落结构和代谢;(4)内生细菌、植物以及土壤之间的生态互作效应研究。以上这些问题的解决有助于加快内生细菌在重金属植物修复中的应用,具有重要的理论和实践意义。
猜你喜欢
小哥白尼(趣味科学)(2021年10期)2022-01-17
学生天地(2020年10期)2020-08-25
湖北农机化(2020年4期)2020-07-24
矿产综合利用(2020年1期)2020-07-24
世界农药(2019年4期)2019-12-30
今日农业(2019年11期)2019-08-15
乡村地理(2018年2期)2018-09-19
中成药(2018年8期)2018-08-29
科技知识动漫(2017年12期)2018-03-07
当代化工研究(2016年6期)2016-03-20
