
色季拉山急尖长苞冷杉天然更新影响因素
2018-11-30 06:12王瑞红李江荣
浙江农林大学学报 2018年6期
王瑞红, 李江荣, 潘 刚
(1.西藏农牧学院 高原生态研究所,西藏 林芝860000;2.西藏农牧学院 西藏林芝高山森林生态系统国家野外科学观测研究站,西藏 林芝860000;3.西藏农牧学院西藏高原森林生态教育部重点实验室,西藏 林芝860000;4.西藏农牧学院 资源与环境学院,西藏 林芝860000)
急尖长苞冷杉Abies georgeivar.smithii属于耐荫性植物,特别是在幼苗期,温凉和寒冷的气候区域比较适宜其生长。常在高海拔地区至低海拔的亚高山与高山地带的谷地、阴坡及半阴坡处形成纯林,与喜冷湿的云杉Picea aspereta,落叶松Larix gmelinii,铁杉Tsuga chinensis,松树Pinus及阔叶树形成针叶或针阔混交林。急尖长苞冷杉是西藏东南部阴坡高山林线森林群落的主要建群树种之一,在色季拉山,主要分布在海拔3 300~4 300 m,分布范围比较广泛[1]。但是由于长期过度采伐,林地生态环境恶化,天然更新比较困难,其自然分布区急剧缩小,种群数量逐渐减少[2]。在实地调查中,西藏色季拉山急尖长苞冷杉幼苗很丰富,但是幼苗向幼树过渡期间死亡率比较高,导致其天然更新效果不理想。目前,国内对于冷杉天然更新影响因素方面的研究比较多,主要集中在不同采伐方式[3-5]、生态学特性[6-7]、海拔与坡向[8-9]、土壤种子库[10-11]及地被物[9,12-15]等。对于西藏高寒环境下急尖长苞冷杉天然更新影响因素的研究还比较少,主要为林隙对其天然更新的影响[16-17]。本研究通过海拔、坡向、种子大小、林隙等方面展开色季拉山急尖长苞冷杉天然更新影响因素的初步研究,为进一步认识色季拉山原始暗针叶林的天然更新,特别是如何采取措施促进其更新具有重要的实践意义。
1 研究区概况
研究区位于西藏鲁朗境内藏东南的林芝地区。色季拉山山体海拔为2 100~5 300 m,水平走向大致呈西北向东南,由此形成了阳坡与阴坡。由于海拔差异较大,林下发育的土壤也出现明显的不同。从低海拔到高海拔呈现的土壤类型依次为山地棕壤、山地酸性棕壤、山地漂灰土、亚高山草甸土、高山草甸土、高山寒漠土[18]。研究区域属典型的亚高山寒温带半湿润气候,雅鲁藏布江为其提供水汽通道,降雨比较丰沛,但干湿季节比较分明。西藏林芝高山森林生态定位研究站数据显示:降水多集中在每年4-10月,年降水量为875~1 350 mm,年平均气温为-0.73℃,7月为最暖月,1月气温最低。该区域乔木以原始暗针叶林急尖长苞冷杉为主,还有少量的林芝云杉Picea likiangensisvar.linzhiensis和方枝柏Sabina saltuaria;林下灌木生长繁茂,主要有西南花楸Sorbus rehderiana,红景天Rhodiola rosea,杯萼忍冬Lonicera lanceolata,悬钩子Rubus corchorifolius,蛾眉蔷薇Omeiensis,林芝杜鹃Rhododendron tanastylumvar.lingzhiense等;草本主要有龙胆Gentiana scabra,委陵菜Potentilla chinensis,西南草莓Fragaria moupinensis,卷叶黄精Polygonatum cirrhifolium,野丁香Syringa persica等;地被层主要包括枯枝落叶层和苔藓层,其中苔藓层比较发达,尤其是阴坡,厚度达10 cm以上,盖度约90%。
2 研究方法
2.1 样地设置
采用梯度格局法设置样地,在阴坡与阳坡均在海拔3 700~4 200 m处,每升高100 m设置1块样地,共设置样地12块,样地面积均为30 m×30 m。采用邻接格子法把样地划分成9个10 m×10 m的样方,各样方设置5个2 m×2 m的小样方进行灌木调查,并在2 m×2 m的样方中设置5个1 m×1 m的小样方调查草本、苔藓植物、凋落物厚度及幼苗数量。在样地中或附近的林下、林缘、林隙处分别设置3个1 m×1 m的小样方,记录各小样方内幼苗数量,测量样方内各幼苗高度、周边苔藓层厚度及凋落物厚度。
2.2 测量方法
2.2.1 球果采集 对海拔3 700~4 200 m样地中急尖长苞冷杉每木检尺,各海拔均选取标准木,待种子成熟初期,将树冠分上层、中层、下层共3层,每层用高枝剪取下5个球果,标记装袋带回实验室。
2.2.2 种子特征及质量测量 从球果中手剥出种子,待种子自然风干后,随机选取各海拔阴坡与阳坡各50粒种子,用游标卡尺测量种子带翅长度、去翅长度、种子宽度、种子厚度,用100粒法测量其千粒重。
2.2.3 苔藓、凋落物厚度及幼苗数量的测量 在海拔3 700~4 200m处设置的5个1 m×1 m小样方中记录幼苗数量,四周分别测量苔藓植物厚度及凋落物厚度,另在林下、林缘、林隙处设置的小样方中记录每个小样方内幼苗的数量,用直尺测量样方四周苔藓层厚度及凋落物厚度。
2.3 数据分析
数据统计分析使用SPSS 19.0,利用Excel 2010制作图表。
3 结果与分析
3.1 海拔与坡向对急尖长苞冷杉天然更新的影响
3.1.1 不同海拔与坡向种子特征差异性分析 海拔与坡向作为重要的环境因子,其对温度、湿度、光照等诸多环境因子产生直接的影响。这些因子对急尖长苞冷杉种子去翅长度、带翅长度、宽度及厚度均产生影响。对色季拉山种子特征研究表明:在阴坡,种子带翅长度、种子去翅长度、种子宽度、种子厚度在海拔3 800或3 900 m处达到最大值,最大值分别为16.87,10.54,3.85,2.31 mm,最小值分别为8.03,4.27,1.69,0.85 mm;在阳坡,种子带翅长度、种子去翅长度、种子宽度、种子厚度亦是在海拔3 800或3 900 m达最大,最大值分别为 17.44,11.52,4.18,2.93 mm,最小值分别为8.79,4.85,1.81,0.72 mm;阳坡种子带翅长度、种子去翅长度、种子宽度、种子厚度要略好于阴坡。由表1可以得出:在阴坡,急尖长苞冷杉种子去翅长度、宽度、厚度均表现为差异不显著,但是种子带翅长度表现为差异显著(P<0.05,F=2.467);在阳坡,种子去翅长度、种子宽度、种子厚度表现为差异性不显著(P>0.05),而种子带翅长度表现为差异显著(P<0.05,F=2.527)。由表2可以得出:种子特征均表现为差异不显著。可见,色季拉山急尖长苞冷杉种子形状相对稳定,同一坡向不同海拔对种子形状影响较小,同一海拔不同坡向对种子形状影响不明显。
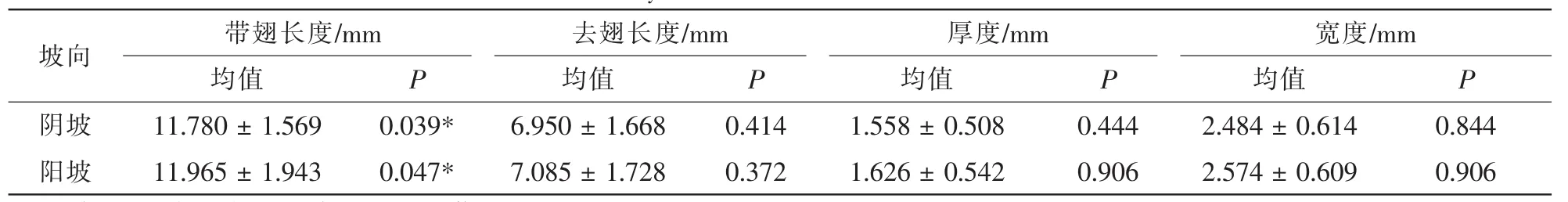
表1 不同海拔种子特征差异性分析Table 1 Difference analysis of seeds characteristic on different elevations

表2 不同坡向种子特征差异性分析Table 2 Difference analysis of seeds characteristic on different slopes
3.1.2 种子质量对天然更新的影响 种子是植物生活史中的一个重要环节,关系着种群未来的命运。种子千粒重在一定程度上可以预知种子的饱满度,同时也是衡量种子品质的重要指标,对种子萌发、休眠、出土、幼苗发育及幼苗生长产生影响,从而影响急尖长苞冷杉林的天然更新。急尖长苞冷杉种子带翅且比较小,主要靠重力扩散及借助风力,扩散能力比较强,扩散距离比较远,可以为急尖长苞冷杉提供更多的生长空间。武高林等[19]研究表明:较小的种子扩散能力强,有利于种群建植和天然更新,能减少动物捕食的数量,可形成持久土壤种子库。在色季拉山,阴坡种子千粒重分布范围为3.32~12.86 g,海拔3 900 m处种子千粒重达到最优值,平均为10.46 g;阳坡种子千粒重分布范围为3.72~14.68 g,在海拔3 800 m处最佳,平均为12.37 g;种子千粒重在阴坡与阳坡随着海拔的升高均呈先升高后下降的趋势;总体来看,阳坡种子质量要优于阴坡。由表3可以得出:阴坡千粒重(P=0.008,F=3.500)和阳坡千粒重(P=0.003,F=4.217)均表现为差异显著;由表4可以得出: 在海拔 3 800 m(P=0.038,F=5.043)和3 900 m(P=0.044,F=4.671)处种子千粒重表现为差异显著;3 700,4 000,4 100,4 200 m处均表现为差异不显著。

表3 不同坡向种子质量差异性分析Table 3 Difference analysis of seeds weight on different slopes
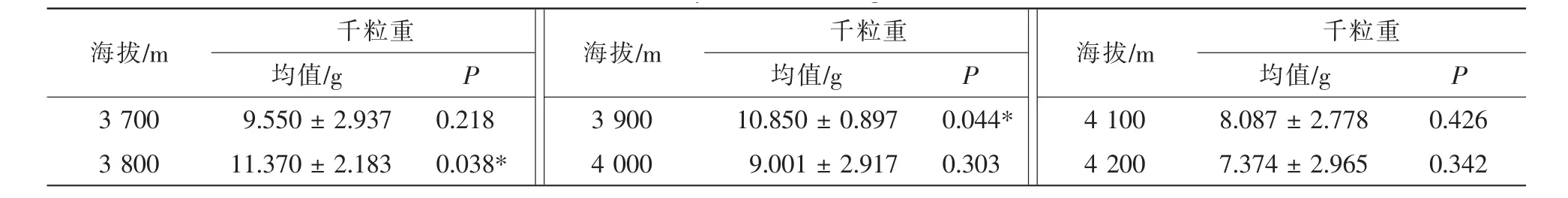
表4 不同海拔种子质量差异性分析Table 4 Difference analysis of seeds weight on different elevations
3.1.3 不同海拔与坡向天然更新幼苗数量 由图1可以得出:随着海拔的升高急尖长苞冷杉幼苗数量总体上呈现先增加后减少的趋势,在海拔3 900 m处达到最大值,阴坡为15.2株·m-2,阳坡为14.2株·m-2;在海拔4 000 m处,由于调查样地处于一乱石滩,水土流失严重,保水能力较差,因此,此处幼苗数量急剧下降;高海拔处幼苗数量递减,主要是由于海拔升高气温会逐渐降低,过低的温度致使土壤温度常处于冻结状态,幼苗根系很难从土壤中吸收水分,其根系的生长受到抑制,再加上光照强度增大,紫外线增强,水分散失比较多,易造成幼苗生理性干旱而死亡;在研究海拔范围内阴坡单位面积内幼苗数量均高于阳坡,主要原因为阳坡光照强烈、温度较高、土壤含水量较低,对耐荫性树种急尖长苞冷杉幼苗的发育不利。

图1 单位面积内不同海拔不同坡向幼苗数量Figure 1 Number of seedlings in per unit area on different elevations and slopes

图2 海拔3 700~4 200 m不同位置幼苗数量Figure 2 Number of seedlings at different positions of 3 700~4 200 m
3.2 林隙对天然更新的影响
林隙是指森林群落中1株以上林冠层树木死亡形成的将会由新个体占有与更新的空间[20]。罗大庆等[17]认为在亚高山急尖长苞冷杉林中,干基折断是形成林隙的主要方式,其次为干中折断与拔根风倒,枯立的较少,且风是形成林隙的主要外力。在色季拉山,急尖长苞冷杉林中林隙广泛分布且面积较大,四周可达10 m,是其天然更新的起点和自然演替的重要机制之一。由图2得出:不同海拔单位面积幼苗数量均表现为林隙>林缘>林下,其中在海拔3 900 m处表现的最为突出,林隙、林缘、林下幼苗数量分别为12.7,8.0,7.5株·m-2;在林隙、林缘及林下均表现为随着海拔的升高急尖长苞冷杉幼苗数量呈现先增加后减少的趋势。
3.3 苔藓、凋落物层对天然更新的影响
苔藓层、凋落物层位于森林植被层与土壤层之间,是森林生态系统的重要组成部分,同时也是林地水汽交换的重要界面[21-24]。由表5可以得出:苔藓层厚度、凋落物厚度均大于死亡幼苗根长,因此导致较多的幼苗死亡,达3.000株·m-2;不同海拔死亡幼苗根长差异性不显著,苔藓厚度、幼苗数量差异性显著,凋落物厚度、死亡幼苗数量差异性极显著。主要有以下原因:苔藓与凋落物具有遮光,减少光照的作用,有利于急尖长苞冷杉种子萌发、幼苗生长;苔藓、凋落物覆盖林地,可以减少动物对种子的觅食,且凋落物层的分解会增加林地养分,促进急尖长苞冷杉幼苗的生长;另外,苔藓层、凋落物层具有明显的持水功能,特别是在干旱胁迫条件下,苔藓层、凋落物层的水分可以暂时补给给幼苗;但是由于苔藓与凋落物的机械阻碍作用,使萌发种子根系在较短的时间很难扎进土壤,随着急尖长苞冷杉幼苗的生长,对水分、光照及其他营养元素的需求增多,使冷杉幼苗缺失水分及其他营养元素而死亡。

表5 苔藓厚度、凋落物厚度、死亡幼苗根长、死亡幼苗数量差异性分析Table 5 Difference analysis of thickness of moss layer,thickness of litter,length of death seeding and number of death seeding
3.4 相关性分析
从相关性分析中可以得出(表6):幼苗数量与林分郁闭度、带翅长度、千粒重、凋落物厚度均呈现显著正相关,与苔藓厚度呈现极显著正相关。这说明色季拉山急尖长苞冷杉林分郁闭度、带翅长度、千粒重、苔藓厚度、凋落物厚度可能有利于急尖长苞冷杉天然更新,苔藓厚度对其天然更新的影响最大。

表6 急尖长苞冷杉天然更新影响因子与幼苗数量之间的相关系数Table 6 Correlation coefficients among factors influencing natural regeneration between each influencing factor and the number of seedlings of Abies georgei var.smithii
4 讨论与结论
由于海拔与坡向的不同,急尖长苞冷杉种子带翅长度、种子去翅长度、种子宽度、种子厚度均存在差异性,尤其是阴坡与阳坡种子带翅长度均表现为显著差异。色季拉山急尖长苞冷杉种子形状相对稳定,同一坡向不同海拔对种子形状影响较小,同一海拔不同坡向对种子形状影响不明显。急尖长苞冷杉冷杉种子较小,扩散距离比较远,可以为急尖长苞冷杉提供更多的生长空间。阴坡种子千粒重分布范围为3.32~12.86 g,海拔3 900 m处种子质量达到最优值,平均为10.46 g;阳坡种子千粒重分布范围为3.72~14.68 g,在海拔3 800 m处最佳,平均为12.37 g;种子千粒重在阴坡与阳坡随着海拔的升高均呈现先升高后下降的趋势;总体来看,阳坡种子质量要优于阴坡。急尖长苞冷杉种子在海拔3 800,3 900 m处千粒重表现为显著差异;在阴坡与阳坡,种子千粒重均表现为显著差异。在海拔3 700~4 200 m,急尖长苞冷杉天然更新幼苗数量均表现为林隙>林缘>林下。幼苗数量与林分郁闭度、带翅长度、千粒重、凋落物厚度均呈显著正相关,苔藓厚度呈极显著正相关,但是苔藓层厚度、凋落物厚度均大于死亡幼苗根长,这样会使急尖长苞冷杉幼苗缺失水分及其他营养元素而死亡,可见苔藓层、凋落物层对急尖长苞冷杉幼苗定居和生长发育具有双重影响。林隙处天然更新幼苗数量较好于林下与林缘,主要是由于林隙内气温、土壤温度及湿度等小气候因子产生影响,改变了急尖长苞冷杉生长的物理环境和生物环境,对维持色季拉山急尖长苞冷杉种群的稳定性有重要作用。急尖长苞冷杉林天然更新的影响因素还有很多,比如土壤种子库、自身的生长基质、伴生物种间的竞争、苔藓及枯枝落叶层的化感作用、自身的化感作用等,因此,对于急尖长苞冷杉天然更新的影响因素还需进一步的研究。
猜你喜欢
种子科技(2021年17期)2021-10-29
思维与智慧·下半月(2021年7期)2021-08-13
林业与生态(2019年9期)2019-10-07
绿色科技(2019年5期)2019-04-12
花火A(2019年1期)2019-03-22
绿色科技(2018年17期)2018-11-05
现代养生·上半月(2017年7期)2017-07-07
发明与创新·大科技(2017年6期)2017-06-20
课程教育研究(2017年11期)2017-04-17
