
骨髓细胞参与肝癌新生血管的构建
2018-11-02 12:21邵茜雯朱海涛凌芸徐静
东南大学学报(医学版) 2018年5期
邵茜雯,朱海涛,凌芸,徐静
(1.南京医科大学第一附属医院 肿瘤科,江苏 南京 210029;2.江苏省肿瘤医院/南京医科大学附属肿瘤医院/江苏省肿瘤研究所 普外科,江苏 南京 210009;3.南京医科大学第一附属医院 检验学部,江苏 南京 210029)
血管新生是肝细胞肝癌的重要特性,肝癌进展时骨髓动员明显,但骨髓干细胞(bone marrow stem cells, BMSCs)是否参与肝癌血管新生还不明确[1]。我们通过细胞示踪技术,观察BMSCs是否参与肝癌细胞(hepatic cells cancer, HCC)的血管新生。
1 材料与方法
1.1 材料
流式抗体AlexaFluor488- CD133、PE- 血管内皮生长因子受体2(PE- VEGFR2)、PE- CD34购自eBioscience公司,大鼠抗小鼠CD31单抗、PE标记羊抗大鼠二抗购自ABCOM公司,血管内皮生长因子(VEGF)- ELISA试剂盒购自eBioscience公司。C57BL/6小鼠70只,雄性,体重18~22 g;骨髓供体:eGFP转基因C57BL/6小鼠10只,雌性,体重18~22 g。小鼠由南京大学模式动物中心提供。
1.2 方法
1.2.1 小鼠原位肝癌模型制作[2]小鼠称重后,自右下腹腹腔注射盐酸氯胺酮(42 mg·kg-1)麻醉小鼠。麻醉成功后仰卧位固定小鼠,自剑突向下1.0 cm切开,提起腹壁,以无菌棉签固定肝脏,肝内注射2×105个H22细胞(总量0.2 ml)(上海细胞研究所)。对照组:小鼠麻醉后肝内注射等量生理盐水。
1.2.2 ELISA测定血清VEGF浓度 上述原位肝癌建模后14 d尾静脉采血,对照组在同时间点尾静脉采血作为对照,4 ℃静止半小时后4 ℃ 2 000 r·min-1离心5 min,取上清保存后统一测定,具体按ELISA说明书操作。
1.2.3 流式细胞术 原位肝癌建模后14 d,模型组以及对照组小鼠均从尾静脉采血300 μl,加入等量红细胞裂解液,震荡5 min后4 ℃ 2 000 r·min-1离心5 min,小心吸取沉淀细胞并重悬于0.5 ml PBS中。按说明书添加流式抗体AlexaFluor488- CD133、PE- VEGFR2和PE- CD34,室温下避光保存30 min后上机测定CD133+VEGFR2+和CD133+CD34+细胞的百分比。
1.2.4 建立GFP+骨髓的小鼠原位肝癌模型[3]受体小鼠11 Gy照射后2 h内经尾静脉注射2×106个GFP+BMSCs(0.2~0.3 ml),SPF级环境中饲养4周后按上述方法建立肝癌模型。
1.2.5 肝组织病理学检查 肝癌建模7 d及14 d后处死小鼠取肝脏,冷冻切片2 μm,分别进行HE及免疫荧光染色。
1.2.6 RT- PCR 肝癌建模后14 d取肝组织,酚-氯仿法抽提总RNA,ExScriptTM逆转录(TaKaRa日本)合成cDNA。引物由TakaRa公司设计合成。CD133:5′- AACGTGGTCCAGCCGAATG- 3′,5′- TCCCAGGATGGCGCAGATA- 3′;CD34:5′- ACCCACCGAGCCATATGCTTAC- 3′,5′- GATACCCTGGGCCAACCTCA- 3′;β- actin:5′- CATCCGTAAAGACCTCTATGCCAAC- 3′,5′- ATGGAGCCACCGATCCACA- 3′。采用SYBR©GreenⅠ嵌合法计算目的基因相对表达水平。
1.2.7 免疫荧光 肝癌建模后7、14 d取肝组织,快速冰冻后5 μm切片,室温固定,封闭液孵育后加一抗CD31(1∶800)4 ℃过夜。荧光二抗(1∶1 000)室温孵育35 min。二脒基苯基吲哚(DAPI)染核,高倍镜下计数GFP+CD31+细胞在血管内皮中的比率。掺入率=GFP阳性核/所有内皮细胞核×100%。
1.2.8 Western blotting 肝癌建模后14 d取肝癌与癌旁组织并提取蛋白,上样100 μg,常规聚丙烯酰胺电泳、转膜,加一抗(1∶200)4 ℃过夜后加二抗(1∶1 000),X线 胶片显影。
1.3 统计学处理
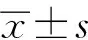
2 结 果
2.1 肝癌组织病理
建模7 d后肝脏出现灰白色结节,镜下见典型的肝癌病理特征,肿瘤新生血管明显。
2.2 肝癌模型小鼠血清VEGF和BMSCs水平
肝癌建模后14 d血清VEGF浓度为(175.13±52.34)pg·ml-1,显著高于对照组的(81.65±32.71)pg·ml-1(P<0.05),同时循环CD133+CD34+和CD133+VEGFR2+细胞明显升高,分别为(0.27±0.13)%和(0.16±0.08)%,显著高于对照组的(0.07±0.04)%和(0.08±0.06) %(P<0.01)。
2.3 建立GFP+骨髓的原位肝癌模型
肝癌小鼠有核细胞GFP阳性率为95%~98%,表明骨髓标记成功。建模后14 d肿瘤中出现大量绿色荧光,直接证明BMSCs动员后富集于肝癌组织中(图1)。同时肝癌组织中BMSCs表型基因CD133、CD34水平显著高于无瘤组织。

A.肝癌组织HE染色结果(×50,2 μm);B.A在荧光显微镜下图像,肿瘤组织中出现GFP(×100)T为肝癌组织,N为正常组织
图1原位肝癌组织快速冰冻切片
2.4 肝癌新生血管中的BMSCs
CD31是血管内皮特异性抗原,镜下肿瘤血管内皮中存在CD31+GFP+细胞,这是内皮细胞来源于BMSCs的直接证据(图2)。随着肿瘤的增殖,BMSCs在肝癌新生血管中的比例从7.6%(建模7 d)逐渐升高到19.3%(建模14 d)(P<0.05)。
2.5 肝癌组织中VEGF表达水平
肝癌组织中VEGF含量高于正常肝组织,见图3。
3 讨 论
研究发现,BMSCs动员后能分化为成熟的血管内皮细胞而参与血管新生[1,4],VEGF是最主要的骨髓动员因子和促血管因子[5]。本实验显示,血清VEGF随着肿瘤生长明显升高,BMSCs大量动员,肝癌组织中VEGF也明显高表达。推测肝癌组织释放VEGF入血动员BMSCs进入循环,受局部微环境的趋化而在局部富集,最终在肿瘤微环境诱导之下分化为成熟的血管内皮而参与肿瘤血管新生。有研究认为除了直接分化为血管内皮外,BMSCs也可在局部分泌促血管生成因子以“旁分泌”方式参与血管新生[6]。
目前对动员后的BMSCs在肿瘤组织中如何分布,观点不尽相同。有人认为肿瘤组织独特的缺氧、低pH、高液压以及高细胞因子水平对BMSCs具有显著的募集作用[7],但也有人认为BMSCs主要在癌旁异常富集[8]。鉴于肿瘤生长有赖于血管的生成,理论上肿瘤组织应该含有更多的BMSCs以促进肿瘤血管新生。实验结果显示动员后BMSCs在肝癌组织中富集,定量PCR结果与其吻合,BMSCs表型基因CD133、CD34在肿瘤中的表达水平显著高于无瘤区。研究结果和临床结果有差异的可能原因:(1) 动物模型没有肝硬化背景,而肝硬化常伴有血管新生[9];(2) 实验肿瘤增殖较血管新生活跃;(3) BMSCs分化丢失干细胞标志后不易为临床检测到,而GFP可以有效示踪BMSCs。某种程度上我们的实验反映了BMSCs在肝癌组织中的自然分布。
有研究发现抗肿瘤治疗导致血清VEGF以及循环BMSCs增高[10]。组织酸中毒、低氧、坏死加剧后更多的细胞因子释放入血,随之更多的BMSCs入血。因此尽管肿瘤血管可以被破坏,但是BMSCs释放入血后可以构建新生血管以补偿肿瘤血供。我们实验发现肝癌建模7 d后BMSCs在血管内皮中的比率约7.6%,随着肿瘤的生长,其比率逐渐达到19.3%。肿瘤初始阶段即需求血管新生为其进一步生长铺垫,直至形成实体瘤。事实上,肝硬化、肝炎、肝癌癌前病变就已经发生了血管新生。当形成肿瘤实体后,血管新生就成为肿瘤明显的特征同时也决定了肿瘤的进展和患者的预后。

肿瘤中可见GFPCD31双阳性细胞(箭头),A.红色,血管内皮CD31;B.绿色,GFP标记的BMSCs;C和D.蓝色,DAPI染色的细胞核 ×300,2 μm,Frozen
图2肝癌组织新生血管中的BMSCs
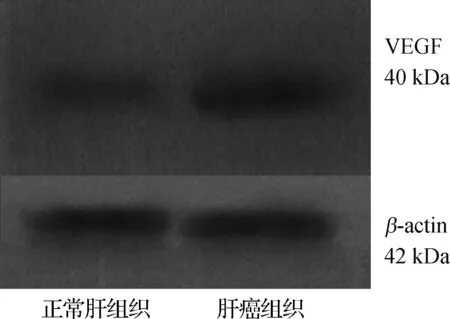
图3肝癌组织与正常肝组织中VEGF蛋白电泳图
我们的研究提示在肝癌生长过程中BMSCs在新生血管构建中起着重要的作用。循环BMSCs或许可以作为肝癌重要的预测指标,阻断骨髓介导的血管新生可能在进一步提高抗肿瘤新生血管治疗效果中起到积极的作用[11]。
猜你喜欢
中国慈善家(2022年1期)2022-02-22
中国临床医学影像杂志(2021年10期)2021-11-22
国际放射医学核医学杂志(2020年3期)2020-07-27
文苑(2018年18期)2018-11-08
海峡姐妹(2017年7期)2017-07-31
现代检验医学杂志(2016年2期)2016-11-14
安徽医科大学学报(2015年9期)2015-12-16
视野(2015年4期)2015-07-26
医学研究杂志(2015年11期)2015-06-10
中国记者(2014年1期)2014-03-01
