
TALEN介导的foxo3a敲除小鼠的表型分析
2018-10-23 01:53赵鲜红刘福航陈忠毅高亚可崔奎青石德顺
中国畜牧杂志 2018年10期
赵鲜红,刘福航,陈忠毅,高亚可,崔奎青*,石德顺*
(1.广西大学动物科学技术学院,亚热带生物资源保护利用国家重点实验室,广西南宁 530004;2.武汉黄浦中西医结合妇产医院,湖北武汉 430000)
foxo3a是叉头框蛋白O亚家族的核心,foxo3a位于第6号染色体上,其cDNA全长2.8 kb,有2个内含子,3个外显子,一共编码673个氨基酸[1],其结构上包含第32位苏氨酸、第253位和第315位丝氨酸3个重要的磷酸化位点。foxo转录因子参与很多不同的生物进程,包括新陈代谢、DNA修复、细胞凋亡、抵抗氧化应激、长寿、生殖等[2-6]。foxo3a–/–雌鼠卵巢出现一些独特的表型—大范围卵泡激活,导致卵母细胞凋亡、功能型卵巢卵泡的早期损耗及继发性不孕。foxo3a在最早期卵泡生长发育阶段作为卵泡发育的抑制因子,另外,foxo3a为研究卵泡生长调控提供了一个分子入口。各位学者对foxo3a的研究为治疗妇女卵巢功能早衰、不孕不育提供了可能[7]。据文献报道,foxo3a还能够保护神经免受氧化应激的损伤并与神经修复有关[8-9];foxo3a是经过实验证实的抑癌基因[10],其磷酸化及亚细胞定位和肿瘤侵袭密切相关[11];另外,foxo3a一直被认为是长寿基因,具有抗衰老、延长寿命的功能[12]。foxo3a在参与这些生物学进程时,主要是通过PI3K/AKT通路,具体过程是磷酸化的PI3K激活AKT,AKT进而激活磷酸化的foxo3a使其与胞核内结构蛋白14-3-3特异性结合,从胞核转移到胞浆,经过泛素化降解,丧失正常的转录活性,从而降低细胞凋亡[13];然而当抑制AKT途径时,去磷酸化foxo3a大部分进人胞核,最终加速细胞凋亡。foxo3a通过激活促凋亡蛋白Bim和FasL诱导细胞凋亡。活化的foxo3a可以升高p27kip1的mRNA以及蛋白水平,抑制周期素依赖性激酶D1,人细胞周期蛋白依赖性激酶2(cyclinE-CDK2)复合物,导致细胞周期停留于G0/G1期,诱导细胞凋亡。
foxo3a敲除早已在猪、牛、小鼠等动物上进行研究,但都主要集中在早期卵泡募集上,对foxo3a敲除动物繁殖力表型研究的报道很少。鉴于此,本研究主要统计不同foxo3a敲除小鼠产仔数并分析原因,为制备foxo3a敲除水牛、黄牛等大型家畜以提高其繁殖力奠定基础。
1 材料与方法
1.1 实验动物 TALEN介导的foxo3a敲除小鼠和WT小鼠,均来自广西大学亚热带生物资源保护利用国家重点实验室石德顺课题组。
1.2 载体和菌种 pMD18-T和感受态宿主E.coli DH5α均购自TaKaRa公司。
1.3 实验试剂 DNA凝胶回收试剂盒、质粒提取试剂盒、DNA Markers、DEPC等均购自天根生化科技有限公司;TRIzol裂解液购自Invitrogen公司;Fast Start Universal SYBR GreenMaster(ROX) 购 自 Roche公司;Oligod(T)18T购自北京全式金生物技术有限公司;PSTI酶、Taq酶、LA Taq酶、GC Buffer、RNase inhibitor购自大连宝生物工程有限公司(TaKaRa);氨苄青霉素钠购自Solarbio公司;LB肉汤、LB营养琼脂购自北京奥博星;普通琼脂糖(BIOWEST AGAROSE)为进口分装产品。
1.4 实验方法
1.4.1 引物设计 根据NCBI网站上公布的小鼠序列,利用Oligo6.71软件分别设计了TALEN介导的foxo3a敲除小鼠后代阳性鉴定所用引物mFtln1和小鼠QRT-PCR所用引物DL-foxo3a。mFtln1-F:5'-AGCACAGATGGG TACTGCTT-3';mFtln1-R:5'-ACCTGTCCTATGCCGAC C-3';扩增片段长度为653 bp,退火温度55℃。DL-foxo3a-F:5'-TACGGCAACCAGACACTCC-3';DL-foxo3a-R:5'-TTCTGATTGACCAAACTCCC-3';扩增片段长度为203 bp,退火温度58℃。引物序列由生工生物工程(上海)股份有限公司合成。
1.4.2 小鼠基因组提取 基因组的提取参照《分子克隆实验指南·第三版》[14]。
1.4.3 小鼠阳性鉴定 小鼠基因组普通PCR产物在华大生物公司进行测序,并对测序结果用DNASTAR-seqMan分析。
1.4.4 总RNA的提取及cDNA的合成 采用TRIZol法提取小鼠卵巢和睾丸的RNA,随后用分光光度计及1%的琼脂糖凝胶电泳检测其纯度和完整性,并置于-80℃保存,备用。采用北京全式金生物技术有限公司的反转录试剂盒进行反转录。反转录体系:总RNA 2 μg,2XTS Reaction mix 10 μL,Trans script®;RT/RT EnZyme Mix 1 μL,gDNA Remover 1 μL,Oligod(T)18T 1 μL,RNase-free Water补充至 20 μL。反应条件:42℃ 30 min,85℃ 5 min,4℃ 终止。
1.4.5 QRT-PCR反应 QRT-PCR反应体系(20 μL):cDNA 产物 100~300 ng,SYBR Premix Ex Taq Ⅱ 10 μL,上、下游引物(10 mol/L)各0.3 μL,用dd H2O补充至20 μL。反应条件:95℃ 5 min;95℃ 30 s;55℃ 30 s;72℃ 30 s并在72℃时收集荧光,共40个循环,实验对每个样品进行3次重复,运用2-ΔΔCt的方法计算目的基因的相对表达量,用内参GAPDH对其进行标准化,将WT小鼠卵巢和睾丸组织作为对照品,其他为实验组样品。
1.5 统计分析 采用SPSS 17.0软件对数据进行One-Way ANOVA方差分析和LSD多重比较,所有计算结果均用平均值±标准误表示。
2 结果与分析
2.1 小鼠后代阳性鉴定与阳性率统计 foxo3a敲除小鼠经过传代培养用于本研究。TALEN介导的foxo3a敲除技术得到很多种foxo3a敲除小鼠模型,但是经过繁殖仅剩下51 bp片段敲除的模型。foxo3a序列设计了1个TALEN位点(表1)。
因为foxo3a-/+小鼠PCR扩增的DNA双链中,其中1条链缺失了51 bp,所以琼脂糖凝胶电泳会显示2条相差51 bp的条带,而WT小鼠仅有扩增出来的653 bp片段(图1);对WT小鼠基因组扩增出来的653 bp用DNAMAN分析酶切位点后,发现在第270位、第285位、第543位有PSTI酶切位点,其中第270位、第285位酶切位点在发现敲除的51 bp目标序列上,故WT小鼠的DNA序列仅能酶切为110 bp和260 bp左右的2条带;foxo3a-/+小鼠的DNA序列能酶切为543、110、260 bp左右的3条带;foxo3a-/-小鼠DNA序列仅能酶切为543 bp和110 bp 2条带(图2);进一步验证阳性小鼠,并对测序结果用DNASTAR-seqMan分析,foxo3a-/-小鼠和foxo3a-/+小鼠测序结果均会出现51 bp碱基的缺失(图3)。

表1 foxo3a-TALEN位点
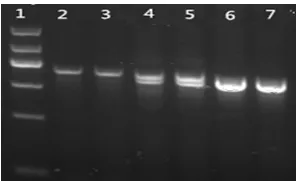
图1 小鼠阳性鉴定PCR

图2 PSTI酶切图
foxo3a-/++ WT后代单敲阳性率为50.00%,阳性率基本符合孟德尔遗传定率(表2);foxo3a-/++foxo3a-/+后代单敲阳性率为55.74%,,双敲阳性率为19.67%,总阳性率为75.41%,阳性率符合孟德尔遗传定率(表3)。
2.2 不同基因型小鼠产仔数统计 由表4可见,相对WT小鼠,foxo3a-/-小鼠和foxo3a-/+小鼠产仔数显著下降(P<0.05);但是foxo3a-/-小鼠和foxo3a-/+小鼠产仔数无显著差异(P>0.05)。

图3 测序结果 DNASTAR-seqMan分析图
2.3 不同基因型小鼠卵巢及睾丸内foxo3a基因mRNA表达量 由图4A知,foxo3a基因 mRNA在foxo3a-/+小鼠卵巢内的表达量下降;由图4B知,foxo3a基因mRNA在foxo3a-/+小鼠和WT小鼠睾丸内的表达量无显著差异。

图4 foxo3a-/+小鼠、WT小鼠卵巢、睾丸内foxo3a基因mRNA的表达量
由图5A知,foxo3a基因 mRNA在foxo3a-/-小鼠睾丸内的表达量升高;由图5B知,foxo3a基因 mRNA在foxo3a-/-小鼠卵巢内的表达量下降。
2.3 不同基因型小鼠卵巢切片HE染色结果 HE染色结果显示,出生后17周的WT小鼠卵巢内还有大量的原始卵泡和初级卵泡;而出生后17周的foxo3a-/+小鼠卵巢内几乎没有原始卵泡和初级卵泡,仅有少量的成熟卵;但是两者的成熟卵泡结构无明显差异;foxo3a-/-小鼠相比foxo3a-/+小鼠,卵巢内卵泡更少(图6)。

表2 foxo3a-/++WT小鼠后代阳性率

表3 foxo3a-/++foxo3a-/+小鼠后代阳性率

表4 不同类型小鼠产仔数

图5 foxo3a-/-小鼠、WT小鼠睾丸、卵巢内foxo3a基因mRNA的表达量

图6 出生后17周WT小鼠、foxo3a-/+小鼠、foxo3a-/-小鼠卵巢切片(40×)
3 讨 论
foxo3a参与细胞周期的调控、细胞凋亡与分化、DNA修复、细胞抗逆性及活性氧(ROS)解毒等生理过程。研究认为,foxO超家族成员中foxo1、foxo3a、foxo4、foxo6是高度保守的PI3K-AKT信号通路中非常重要的下游基因,在造血系统中,PI3K信号通路能调节白血病和自身免疫缺陷病的发病机制,破坏PI3K信号通路会造成免疫缺陷,但是敲除任何一个foxO家族成员,不会出现造血表型异常敲除foxo1、foxo3a、foxo4,造血干细胞会出现明显的缺陷。foxo1敲除动物在胚胎发育的10.5 d会出现胚胎死亡由于血管生成异常[15];foxo4敲除动物是可育的而且没有明显的表型[16];foxo3a敲除动物在出生后15周不育是因为卵巢内原始卵泡大范围募集随后卵母细胞耗尽[17-18],而且foxo3a敲除动物模型会出现淋巴细胞增生和炎症[19]。因此,如何调控foxo3a的表达,促进动物发情周期多排卵进而提高水牛等大型经济动物的繁殖力是值得研究的课题。
本研究利用实验室现有的foxo3a 51 bp敲除小鼠为模型,对其后代进行基因型鉴定并统计后代的阳性率,发现foxo3a-/+小鼠和WT小鼠交配,后代阳性率约为50.00%;foxo3a-/+小鼠和foxo3a-/+小鼠交配,后代阳性率大概为76.74%,符合孟德尔遗传定律,同时也表明小鼠阳性鉴定的正确性。对不同基因型小鼠进行产仔数统计分析,发现foxo3a-/-小鼠和foxo3a-/+小鼠产仔数相比野生型来说,均明显下降。而且foxo3a-/-小鼠和foxo3a-/+小鼠产仔数却没有显著差异。18周以后小鼠处于老龄化阶段,统计了出生后7周至25.5周的foxo3a-/+雌鼠产仔数,结果foxo3a-/+雌鼠没有出现随年龄增长产仔数下降的趋势,也没有随年龄增长而出现产仔数提高的现象。QRT-PCR结果表明,foxo3a mRNA在WT小鼠的心脏、肝脏、肺、肌肉、骨骼、肾脏、卵巢和睾丸均有表达,在肺和卵巢内高表达,表明foxo3a分布广泛,在小鼠的肺和卵巢组织起着重要作用。foxo3a mRNA在foxo3a-/+小鼠卵巢内表达量显著下降,在睾丸内的表达量和WT小鼠相比无明显差异;foxo3a mRNA在foxo3a-/-小鼠卵巢内表达量显著性下降,在foxo3a-/-小鼠睾丸内表达量却显著增加。这可以从定量角度说明小鼠产仔数下降和小鼠卵巢及睾丸内的foxo3a mRNA表达量下降有直接关系,可能是因为敲除的51 bp片段编码的氨基酸正好包括foxo3a结构域中非常重要的第32位苏氨酸位点,造成PI3K-AKT-FOX通路受阻,foxo3a不能和细胞核内的14-3-3蛋白结合,不能转移到细胞质中只能停留在细胞核,调节下游的细胞凋亡因子BIM、FASL、TRAIL、SOD2以及细胞周期调节因子CyclinG2、P27、CyclinD促进卵母细胞凋亡并阻滞细胞周期,造成单敲和foxo3a-/-小鼠卵巢内卵母细胞数量少,排卵率低从而造成单敲和foxo3a-/-小鼠产仔数下降。
一般情况下,小鼠卵巢卵泡发生规律如下:出生后7 d,小鼠的始基卵泡池形成,并且初级卵泡开始形成;出生后3周,小鼠断奶;出生后5周至7周,小鼠的性成熟阶段;出生后9周至12周,小鼠的最佳繁殖期。由于饲养和管理条件不同,小鼠性成熟日龄也有所差异,一般雌性为35~50日龄,雄性为45~60日龄。一般在出生后2~3个月,小鼠呈现体成熟,此时小鼠体重增长缓慢,是小鼠的最佳配种年龄。本实验采集17周龄的WT雌鼠、foxo3a-/+雌鼠及foxo3a-/-雌鼠的卵巢用于卵巢切片,并对切片进行HE染色,观察不同基因型小鼠卵巢内卵泡的发育情况。HE染色显示,出生后17周的WT小鼠卵巢内还有大量的原始卵泡和初级卵泡,而foxo3a-/+小鼠卵巢内几乎没有原始卵泡和初级卵泡,但是两者的成熟卵泡结构无明显差异,这也解释了foxo3a-/+小鼠产仔数下降是由于体内foxo3a基因减少进一步造成卵巢内卵泡减少、小鼠排卵率下降造成的。
本研究中,foxo3a-/+雄性小鼠的睾丸中foxo3a相对表达量和WT小鼠睾丸中相对表达量差异不明显,而雌性却差异明显;foxo3a-/-小鼠雌性卵巢内的foxo3a相对表达量比WT小鼠少,而foxo3a-/-小鼠雄性睾丸内foxo3a表达量却是WT小鼠的30倍左右,这可能是雌性和雄性体内的激素表达不同及foxo3a参与的各种信号通过共同影响的结果。
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23
中国临床医学影像杂志(2022年5期)2022-07-26
中国畜牧杂志(2020年8期)2020-08-16
做人与处世(2020年10期)2020-06-29
中国生殖健康(2019年7期)2019-01-06
科技创新与品牌(2018年5期)2018-07-24
新农村(2018年35期)2018-04-02
猪业科学(2017年3期)2017-04-06
中国猪业(2013年7期)2013-01-23
现代家庭(1999年7期)1999-06-14
