
茶树挥发性萜类物质研究进展
2018-10-15 02:49王让剑孔祥瑞
茶叶学报 2018年3期
王让剑,杨 军,孔祥瑞
(福建省农业科学院茶叶研究所/国家茶树改良中心福建分中心,福建 福安 355015)
香气是挥发性物质种类和含量的综合效应,是评价茶叶品质的重要因子之一。茶叶香气物质占茶叶干重的比例很小,但种类繁多。在众多挥发性物质中,挥发性萜类物质是构成茶叶花果香的重要来源之一,该类化合物还具有化感作用、防虫抗病等生物学功能,是茶树中一类重要的次生代谢物质。本文对茶树中萜类挥发性物质研究进展进行归纳,并对其发展趋势进行展望,以期为今后相关研究提供参考。
1 茶叶中挥发性萜类的生物学功能
萜类化合物是种类、数量最丰富的一类植物、微生物次生代谢产物。已发现的各种萜类物质超过55000种[1],其中倍半萜是最大的一类,达到7000种[2]。萜类均具有“异戊二烯”的基本结构单元,除以萜烯的形式存在外,还以各种含氧衍生物的形式存在,包括醇、醛、酸、酯、酮等。根据分子中碳原子数目可将萜类分为半萜、单萜、倍半萜、二萜等,其中半萜、单萜和倍半萜属于“低萜”,具有挥发性,二萜及以上的化合物属于“高萜”,一般不具有挥发性(表1)。
茶叶中已分离鉴定的萜类有近百种,主要是挥发性的单萜和倍半萜[3],这些挥发性萜类香味活性高、香气阈值低,常具有宜人的花果香,是构成茶叶香气的重要物质基础[4],常见挥发性萜类香型特征如表2。

表1 萜类化合物的分类

表2 挥发性萜类香型特征
萜类化合物具有化感作用,尤以单萜和倍半萜类活性最强[5]。萜类等具有化感活性的物质向环境中释放,能抑制或促进自身及其邻近植物的生长发育,具体表现为排异作用或自毒作用。加州蒿和银叶鼠尾草的叶中含有丰富的挥发性萜类物质,可抑制其它草本植物种子的萌发和幼苗生长,因而其灌丛植物的周围可出现裸带[6]。华北落叶松枝叶挥发物对自身种子表现出延迟萌发和抑制萌发的作用,20个挥发物中,单萜占5个,倍半萜占12个[7]。倍半萜内酯类化合物可以促进弯管列当的发芽和生长,使红豆下胚轴生长减少40%[8]。
萜类化合物对植食性昆虫具有直接或间接的防御作用。茶树芽梢的挥发物是害虫向茶树定位的信息物质,茶尺蠖、茶蚜、假眼小绿叶蝉对芳樟醇、香叶醇等挥发物具有较强的趋性,但单个化合物的引诱活性不如全组分芽梢挥发物[9]。天然茶树芽梢释放的挥发物对害虫具引诱活性,但对天敌一般无引诱活性,但害虫加害茶树芽梢后诱导形成的新的挥发物(HIPVs)则对天敌具很强的引诱活性,而对害虫一般没有引诱活性[10-12]。α-法尼烯是虫害诱导挥发物的主要成分之一,茶树叶片受茶小绿叶蝉危害后会释放出α-法尼烯,可显著引诱叶蝉寄生性天敌缨小蜂[13,14],α-法尼烯在一定浓度下还可对茶小绿叶蝉表现出排斥效应[15]。此外,研究还表明包括芳樟醇在内的挥发性混合物能引起茶尺蠖绒茧蜂最强的EAG反应[16],在低、高剂量下,门氏食蚜蝇对橙花醇分别有极显著和显著的趋性[17]。
萜类化合物还具有抑菌和杀菌作用。芳樟醇、香叶醇等萜类物质对革兰氏阳性菌、阴性菌及一些真菌具有一定的抑制作用[18]。在受茶饼病菌侵染的云南大叶种茶树叶片中,萜类、含氮含硫化合物、芳香族化合物、绿叶性气体为主要挥发物,萜类中芳樟醇是主要的挥发性物质[19]。茶叶中游离态香气组分及键合态糖苷类香气前体均对茶云纹叶枯病致病菌具有不同程度的抑制作用,以香叶醇抑制作用最强[20]。此外,香叶醇在浓度为0.1 mg·mL-1时对茶炭疽病菌的抑制率达到100%[21]。
2 茶叶中挥发性萜类的生物合成
2.1 挥发性萜类的生物合成
茶鲜叶中挥发性物质的来源主要包括萜类的生物合成路径、莽草酸代谢路径以及脂肪酸代谢路径,萜类挥发性物质来自萜类的生物合成路径(图1)。
萜类的生物合成过程主要包括3个阶段:异戊烯基二磷酸酯(IPP)及其双键异构体二甲基烯丙基二磷酸酯(DMAPP)生成阶段;香叶基二磷酸GPP(香叶基香叶基二磷酸GGPP)、法尼基二磷酸FPP生成阶段;萜类生成及其修饰阶段,包括氧化还原、酰化、糖基化等。第1个阶段中IPP及DMAPP有两个合成途径,即质体中的MEP途径和细胞质中的MVA途径。前两个阶段的合成过程已比较清晰,是所有的萜类化合物所共有的阶段,第3个阶段决定了萜类化合物结构多样性,是植物次生代谢研究的重点领域[22]。
参与萜类生物合成途径的酶也可分成3类。第1类是催化IPP及DMAPP合成的酶,如:3-羟基-3甲基戊二酰辅酶A还原酶(HMGR)、脱氧木酮糖-5-磷酸合成酶(DXS)、脱氧木酮糖-5-磷酸还原异构酶(DXR)等。第2类是催化IPP及DMAPP形成各种中间体或萜类终产物的酶,如各种萜类合成酶等。第3类是对萜类终产物进行结构修饰的酶,如甲基化、羟基化、酰基化、糖基化等。合成途径中的关键酶决定了下游萜类产物合成与积累的量,通过调控这些关键酶基因的表达水平可以调节相关萜类物质的产量[23]。迄今为止,已经克隆了茶树萜类物质合成途径中的3-羟基-3甲基戊二酰辅酶A还原酶(HMGR)、脱氧木酮糖-5-磷酸合成酶(DXS)、脱氧木酮糖-5-磷酸还原异构酶(DXR)、法尼基焦磷酸合成酶(FPS)、香叶基香叶基焦磷酸合成酶(GGPS)、芳樟醇合成酶(LIS)、4-羟基-2-甲基-2-E-丁烯基-4-焦磷酸合酶(HDS)、4-羟基-2-甲基-2-E-丁烯基-4-焦磷酸还原酶(HDR)等基因[24,25]。
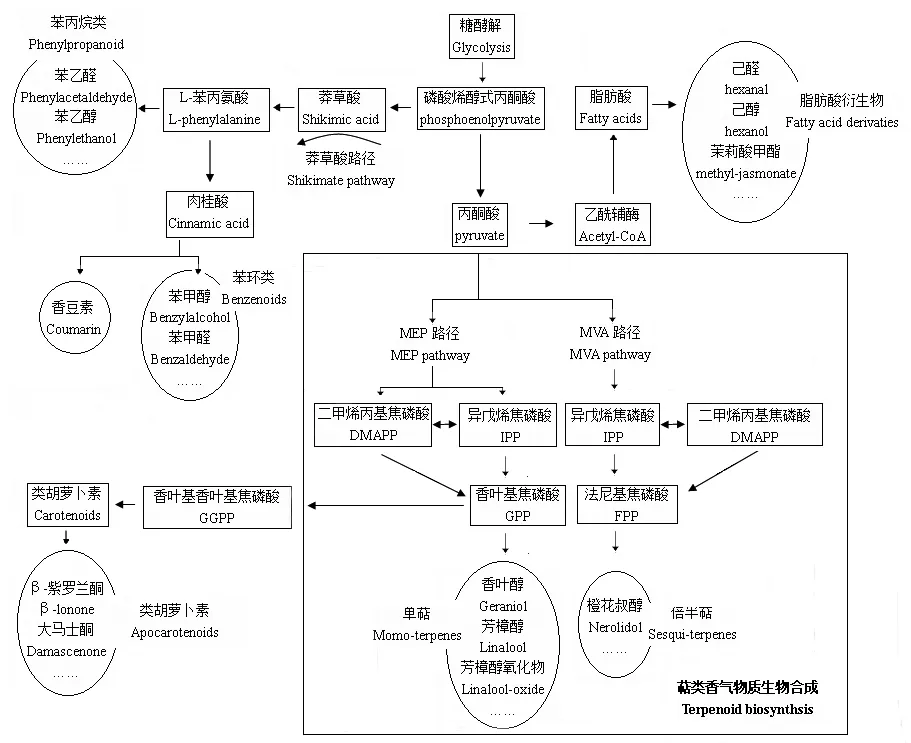
图1 茶树鲜叶中挥发性香气物质生物合成途径
萜类合成酶是萜类化合物合成的关键酶。萜类合成酶结构上的细微改变可以产生新的催化活性,能催化单一底物形成多种产物,是萜类化合物结构多样性的主因[26-28]。研究表明,倍半萜合成酶一般无信号肽,主要定位于细胞质,而单萜和二萜合成酶有一段定位于质体的信号肽,主要定位于质体上[29],也有研究表明单萜和二萜可在细胞质及内质网中生成[30]。目前为止,已在拟南芥、葡萄、烟草等40多种植物中克隆了200多个单萜和倍半萜合成酶基因[26]。茶叶中萜类合成酶研究近年来获得了重要突破,茶树芳樟醇/橙花叔醇合成酶基因(CsLIS/NES)可生成2个转录本:全长CsLIS/NES-1与断头的CsLIS/NES-2,前者含定位于叶绿体的转运肽,后者则丢失该转运肽,两者分别定位于叶绿体和细胞质,离体条件下,二者均可催化生成芳樟醇/橙花叔醇,但在机体内CsLIS/NES-1仅催化芳樟醇的生物合成,而CsLIS/NES-2仅催化橙花叔醇的生物合成[31]。
2.2 挥发性萜类的合成调控
挥发性萜类的生物合成在很大程度上受到昆虫或微生物诱导。研究表明茶小卷叶蛾[32]、茶丽纹象甲[33-34]、茶尺蠖[35]取食茶树后,均能诱导茶树释放芳樟醇、α-法尼烯、橙花叔醇和罗勒烯等萜类挥发物。当茶树遭受病虫害侵害时,叶片中一些萜类合成酶基因的表达明显增强。茶尺蠖取食茶树后,挥发性萜类生物合成途径中共有9个基因表达上调,分别包括MVA路径中2个3-羟基-3甲基戊二酰辅酶A合成酶(HMGS)与2个3-羟基-3甲基戊二酰辅酶A还原酶(HMGR),MEP途径中1个羟甲基丁烯二磷酸合成酶(HDS)及1个羟甲基丁烯二磷酸还原酶(HDR),3个萜类合成酶(TPS)[36]。茶树萜类合成MVA路径中的1个HMGR能被茶尺蠖和小绿叶蝉显著诱导上调表达,进一步证明虫害能诱导萜类生物合成途径中的关键酶基因的上调表达[37]。我们前期在研究茶丽纹象甲取食茶树新梢后其基因表达谱变化时亦发现茶树萜类合成途径中的1个法尼醇脱氢酶基因表达显著上调(未发表)。
紫外照射及激素诱导也能影响萜类物质的生物合成。紫外条件下,茶树HGMR、DXR、GGPS基因表达强度随时间延长逐渐增加,FPS基因则先降后升,DXS表达总体下调,而Lins基因在处理6 h内先升后降[24]。用茉莉酸甲酯、水杨酸、ABA等激素处理茶树后发现HDS与HDR基因均呈现不同程度的上调和下调表达[25]。用茉莉酸甲酯处理茶树芽叶后,芳樟醇、香叶醇及苯乙醇含量明显增加,则直接证明激素诱导能提高茶树叶片中萜类物质的生物合成量[38]。
3 茶叶中的挥发性萜类的存在方式
茶树在自然状态下释放的萜类物质主要来自花,在花蕾绽放初期,芳樟醇、香叶醇以及橙花醇等单萜类的含量明显升高,吸引昆虫授粉[39-40]。茶鲜叶中游离态的挥发性萜类化合物含量远低于键合态的萜类糖苷[41],这也是茶鲜叶一般无香味的主要原因。在茶叶加工过程中,体内积累的糖苷会在内源糖苷酶的作用下水解,释放出挥发性萜类苷元,从而表现出花果香特征。目前已经从茶叶中分离鉴定了多种萜类糖苷,基本为二糖苷和单糖苷,其中二糖苷以β-樱草糖苷为主,单糖苷以β-葡萄糖苷为主,且β-樱草糖苷含量远高于β-葡萄糖苷[42-44]。
茶叶中的糖苷酶是参与糖苷水解的一类重要酶,以β-葡萄糖苷酶与β-樱草糖苷酶的研究居多。β-葡萄糖苷酶水解β-D-糖苷键,释放出β-D-葡萄糖和相应的配基,而β-樱草糖苷酶水解樱草糖苷生成二糖和相应的配基,但保留二糖内部化学键。目前为止,有关茶叶中β-葡萄糖苷酶与β-樱草糖苷酶的蛋白分离纯化[45-46]、基因克隆表达[47-50]等研究已有较多的报道。
糖苷的存在,一方面可能是有利于其在体内运输,另一方面可能是以一种低毒或脱毒的形式避免其苷元对植物自身造成毒害[51]。糖苷的形成与糖基转移酶有着重要的联系,该酶通常催化活化的尿苷二磷酸-D-葡萄糖向糖基受体转移葡萄糖分子形成糖苷,几乎存在于所有生命体中,大量的代谢过程均需要其参与[52]。糖基转移酶是一个复杂的家族,不同植物中糖基转移酶家族成员数目不同,小立碗藓含有17个糖基转移酶家族1成员,而桉树则含有377个家族1成员[53]。近年来随着基因组测序技术的发展,大量的糖基转移酶基因被发现,但多数基因的功能均未进行验证,模式植物拟南芥中进行功能验证的糖基转移酶基因不到其总数的20%[54]。值得一提的是,利用代谢组学方法联合基因表达以及重组蛋白功能鉴定技术验证了第一个单萜糖苷糖基转移酶的生物学功能[55],这为今后开展糖基转移酶功能鉴定提供了参考。茶叶中糖基转移酶相关研究目前尚未见报道。
研究表明β-葡萄糖苷酶可以催化糖苷转移反应,形成新的糖苷化合物[56]。漳平水仙茶加工过程中晾青叶糖苷类香气前体含量较鲜叶有显著增加,摇青叶在做青过程中可能存在着糖苷酶解与合成的动态平衡[57],亦说明β-葡萄糖苷酶可能存在催化糖苷合成的生物学功能。
4 展望
挥发性萜类化合物是茶叶香气的重要来源,且对茶叶病虫害具有抑制作用,是茶叶中一类重要的次生代谢物质,其在茶树芽叶中主要以糖苷形式存在。活体状态下,糖苷类与糖苷水解酶类是隔离的,当茶树叶芽经历病虫害侵袭、损伤以及加工时,细胞破裂导致糖苷与糖苷水解酶充分接触并在其作用下水解,释放出挥发性的萜类化合物,从而表现出香味、抗病虫等生命特征。
虽然萜类的生物合成途径已基本清晰,但茶树芽叶中的萜类多以糖苷的形式存在,其合成机制仍不明确,因此糖苷类在茶树叶芽中是如何合成和累积的将成为今后茶叶香气形成机理及茶树病虫害防治研究的重点之一。随着茶树基因组的破译[58-59]以及功能基因组、转录组、代谢组等技术在茶学研究中的逐步应用,茶树芽叶中萜类糖苷生物合成途径及其累积机理的充分解析必将得以实现。
乌龙茶最显著的特征是具有自然优雅的花果香,研究表明[60]适制乌龙茶品种糖苷类香气前体含量较高,绿茶品种则较低,因此在高香型茶树品种选育过程中宜加强筛选高糖苷含量的茶树种质资源。
猜你喜欢
中草药(2022年21期)2022-11-05
中国农业科学(2021年1期)2021-01-14
湖南中医药大学学报(2020年8期)2020-10-09
核农学报(2020年4期)2020-03-13
应用化工(2019年11期)2019-12-03
中成药(2018年11期)2018-11-24
天然产物研究与开发(2018年7期)2018-08-21
中国酿造(2018年4期)2018-05-10
中文信息(2017年2期)2017-04-13
中国调味品(2017年2期)2017-03-20
