
晚霜冻胁迫后冬小麦株高降低及其与籽粒产量关系
2018-10-11 02:16武永峰胡新任德超史萍游松财
中国农业科学 2018年18期
武永峰,胡新,任德超,史萍,游松财
晚霜冻胁迫后冬小麦株高降低及其与籽粒产量关系
武永峰1,胡新2,任德超2,史萍1,游松财1
(1中国农业科学院农业环境与可持续发展研究所/农业部农业环境重点实验室, 北京 100081;2商丘市农林科学院小麦研究所,河南商丘 476000)
【目的】探究晚霜冻胁迫对冬小麦株高及其构成因素的影响,阐明株高降低特性及其与节间长、穗长和籽粒产量的内在关系,为晚霜冻害评估指标的建立提供依据。【方法】基于低温室和田间可移动式霜箱2种模拟霜冻手段,分别以冬小麦幼穗发育阶段(小花原基分化、雌雄蕊原基分化、药隔形成、四分体形成和抽穗期)和零下处理温度(-1℃、-3℃、-5℃、-7℃、-9℃和-11℃)为梯度,共开展6期盆栽试验和3期小区试验;在考察植株茎部冻害、测定株高与其构成因素、统计籽粒产量要素的基础上,运用方差分析、回归函数拟合、以及突变检验等方法研究晚霜冻胁迫下株高降低特性,分析各构成因素对株高的贡献、以及株高与籽粒产量的回归关系。【结果】(1)在雌雄蕊原基分化至药隔形成后期,株高随处理温度降低而呈突变性降低特征,处理温度低于-5 ℃左右时突变开始,且不同植株个体、品种间有一定差异;在同一处理温度下,株高最大降幅出现在药隔形成后期。(2)在雌雄蕊原基分化期、药隔形成前期和药隔形成后期,对株高贡献排前两位的节间分别为倒四节间和倒三节间、倒三节间和倒二节间、倒二节间和倒四节间,其长度均因冻害胁迫而显著缩短,且与株高呈极强显著相关性(<0.001),此时株高亦呈显著降低趋势。(3)株高与穗粒数、千粒重、单株产量之间的回归模型符合幂函数曲线特征,其中,单株产量的降幅随株高降低而呈现先快后慢的变化态势;当单株产量因冻害胁迫降低至1.5 g以下时,其随株高继续降低而不再明显减少,此时穗粒数变化也已不大。【结论】在模拟晚霜冻胁迫条件下,正在伸长或待伸长的冬小麦节间长度与穗长显著缩短;当缩短节间与对株高起主要贡献的节间相一致时,株高显著降低。利用节间缩短特性、以及单株产量和株高降低之间的幂函数关系模型,可为冬小麦生长后期植株倒伏以及产量损失风险评估方法提供新的研究途径。
冬小麦;晚霜冻胁迫;发育进程;株高;节间和穗长;籽粒产量
0 引言
【研究意义】株高作为冬小麦的一个重要农艺性状,其变化与抗倒伏性、收获指数、产量和生物量等密切关联[1]。冬小麦在分蘖[2]、越冬[3]或拔节期[4]受到霜冻胁迫后,株高均可能出现降低现象。拔节后霜冻通常称为晚霜冻,在我国频繁发生于黄淮麦区,此时冬小麦抗冻性相比前期明显变差[5],因而产量损失风险更大。但由于冻害发生日期距小麦成熟仍有一段时间,其自身会通过后期生长恢复、以及提高穗数(新增分蘖)和粒重等补偿方式,在一定程度上弥补产量损失,从而对晚霜冻害影响评估及预测造成困难[4, 6-7]。相比之下,株高是一个稳定的农艺性状指标,不仅易于在全生育期进行原位观察和测定,而且有特定的节间伸长规律,并在开花后定长[8],这为晚霜冻害评估指标和方法研究开辟了一条可行的途径。【前人研究进展】晚霜冻发生在冬小麦拔节至成熟的各个阶段。Woodruff等[9]认为抽穗前冠层气温低于-6℃即可使幼穗和茎发生明显伤害症状。此时,幼穗因其不同部位冻害而形成各种残穗,致使穗粒数和产量降低[4, 10-11]。茎的冻害症状则往往出现在接近于节的位置,其组织首先变水渍状,随后皱缩和变干,甚至直接因破裂而发生倒伏[12],并最终枯死。对于没有出现明显冻害症状的茎而言,仍能继续生长并为籽粒形成输送营养物质[13]。尽管如此,继续生长的茎仍可能受到一定程度的隐性伤害,主要表现在两个方面:一是营养传导能力下降,导致穗部发育受限,并影响粒重[14];二是节间缩短,株高相应降低[4, 15]。Whaley等[4]通过分析大田自然霜冻条件下小麦冻害与节间关系,认为雌雄蕊原基分化至药隔形成期冻害阻碍了小麦穗下节间、倒二节间和倒三节间的伸长,因此使其长度缩短。对于大田降温环境而言,直接导致茎在短时内破裂倒伏的现象较少出现,后期群体株高降低特征明显。【本研究切入点】株高降低与其构成因素(即节间和穗的长度)的贡献密不可分,并对产量变化具有重要指示意义。针对黄淮麦区晚霜冻害主要发生在拔节至抽穗期这一特点[16-17],前人研究并未系统考虑不同发育进程下、各种降温程度引起的株高及其构成因素降低规律,也较少涉及冻害胁迫下株高与籽粒产量的定量分析,因而对其内在关系仍不甚清楚。【拟解决的关键问题】本研究利用低温室和田间可移动式霜箱2种霜冻模拟手段,以发育进程和胁迫温度为2个梯度来设计控制试验,研究株高降低特性及与其构成因素、籽粒产量的关系,以期为晚霜冻害动态监测与评估提供思路和依据。
1 材料与方法
1.1 试验设计
基于低温室模拟霜冻的试验地点位于商丘市双巴境内小麦试验基地,基于移动霜箱模拟霜冻的试验地点位于商丘市农林科学院南侧小麦试验田。两者管理方式均为大田常规管理,土壤类型为潮土,土壤肥力水平可确保冬小麦正常生长获得所需的营养。相关要素如表1所示。
1.1.1 基于低温室的晚霜冻模拟试验 供试小麦品种为周麦22(ZM22),半冬性,中熟。试验用盆为直径25 cm、高度35 cm、两头开口的圆柱型空心管,播种前先按序排列埋于大田之中,使其顶部与地表基本持平,这样做的目的是尽量保证盆栽冬小麦生长环境等同于大田。土壤过筛后自然风干,施肥后搅拌均匀,再分别装盆、浇水。于2015年10月5日播种,每盆均匀定株11粒。
每次降温处理前,随机拔取3株盆栽小麦,在电子显微镜下观察幼穗所处分化阶段,并记录。将参与试验的小麦盆搬至低温室内,做好冻前准备工作。低温室的长、宽和高分别为7 m、5 m和2.6 m,在其顶部安装2条尼龙布形成的通风管道,分别与低温室外的两台制冷压缩机相连。在管道两侧壁钻出直径为5 cm且均匀分布的若干圆形通风口,冷风通过圆形通风口在1.9 m以上的水平方向上进行流动并形成制冷空间,冷空气均匀下沉,从而形成强辐射降温[18]。降温处理设计如表2所示。一个降温过程持续6 h,采用温控系统来模拟自然降温过程,以确保冻害影响机制类似于大田环境[18]。降温处理完成之后,再将小麦盆搬回大田原处,恢复原来生长环境。试验1至试验4的每个降温处理有3盆重复,1盆对照,试验5和试验6的每个降温处理有4盆重复,1盆对照。
1.1.2 基于田间可移动式霜箱的晚霜冻模拟试验 2011—2012年度供试小麦品种为豫麦18(YM18)、周麦18(ZM18)和偃展4110(YZ4110),其中豫麦18和偃展4110为弱春性早熟品种,周麦18为半冬性中熟品种;于2011年10月8日种植5行,最外行(第1和第5行)作为保护行,第2、第3和第4行分别对应为豫麦18、周麦18和偃展4110。2013—2014年度供试小麦品种为周麦22(ZM22),于2013年10月5日种植5行。所有小麦均以手工点播,行宽14 cm,株距4 cm。

表1 晚霜冻模拟试验相关要素
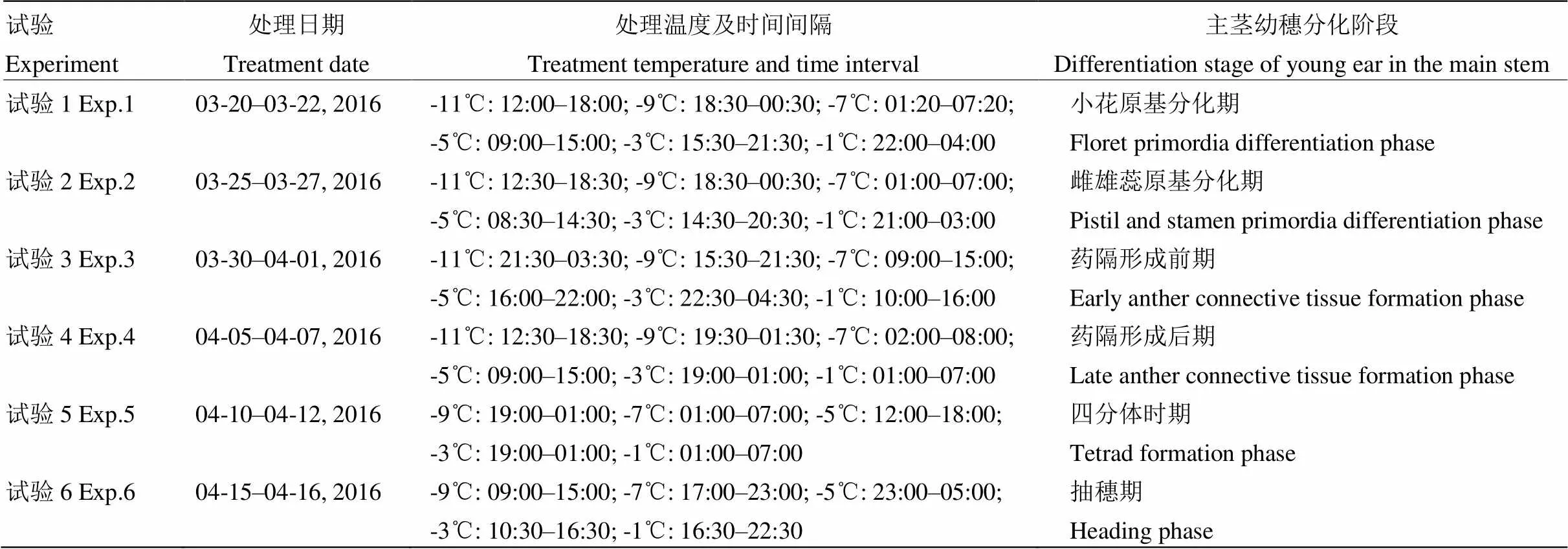
表2 基于低温室的降温处理设计
每次降温处理前,随机拔取3株小麦,在电子显微镜下观察幼穗所处分化阶段,并记录。各试验均需去掉最外行,并沿种植行方向划分出边长不超过60 cm且间隔1米的6个样区。其中,5个样区进行降温处理,第6个样区不作任何处理(对照,CK)。田间可移动式霜箱内径为60 cm×60 cm、水平温差小于0.5℃。将该霜箱完全覆盖样区,四周作密封处理,降温处理过程依据温控系统设定的降温曲线来完成[6, 18]。降温处理设计如表3所示。试验7和试验9的每次降温过程持续6 h,3 h内降至最低设定温度并保持2 h,之后1 h内升至周围环境温度;试验8的每次降温过程持续8 h,4 h内降至最低设定温度并保持3 h,之后1 h内升至周围环境温度。一次降温过程结束之后,移开田间可移动式霜箱,恢复大田生长环境条件。

表3 基于田间可移动式霜箱的降温处理设计
1.2 植株茎部冻害考察与株高、产量测定
在降温处理结束后一周内考察叶片和茎节冻害症状,若出现因变色、水渍或破裂等导致的枯死现象,则进行记录。从试验结束直至小麦成熟,植株未再受各类自然灾害胁迫影响。针对田间样区的每个小麦行,均随机选取10株,分别测定最高茎秆的高度作为株高(Plant height,PH),接着测其穗长(ear length,EL)、穗下节间长(peduncle length,PL)和倒二节间长(penultimate internode length,PIL);针对所有盆栽小麦,处理盆均选取8株,对照盆均选取11株,采取与样区小麦相同测定方法,分别测定株高、穗长、穗下节间长、倒二节间长、倒三节间长(antepenultimate internode length,AIL)和倒四节间长(fourth internode length from the top,FIL)。针对田间样区和盆栽小麦的各个处理,分别统计所有茎蘖数、粒数和粒重,进一步计算单株穗数(ear number per plant,EN)、穗粒数(kernel number per ear,KN)、千粒重(1 000-kernel weight,KW)和单株产量(yield per plant,YP)。
1.3 数据处理及分析方法
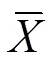
1.3.2 统计分析 (1)利用一元线性函数对株高及其构成因素随处理温度降低的变化趋势进行线性拟合,若拟合精度达到0.05、0.01或0.001的显著性水平,则认为呈显著、强显著或极强显著变化。(2)利用方差分析方法(ANOVA)检测各处理温度间株高及其构成因素差异的显著性。一旦方差分析通过显著性检验(<0.05),则采用Tukey方法(方差齐时)和Dunnett’s T3方法(方差不齐时)进行多重比较。(3)计算单期试验中每个处理的株高及其构成因素的变异系数(coefficient of variation,CV),以比较各处理间离散程度的大小。变异系数的优点在于可以消除株高与其构成因素值相差过大所造成的影响,其计算公式为原始数据标准差与平均值的比值。(4)利用突变检验方法检测株高降低是否存在突变现象,利用逐步回归分析方法建立低温室试验中各构成因素(自变量)与株高(因变量)之间的多元线性回归模型,并依据直接通径系数来比较各因素对株高降低的贡献。(5)利用线性、二次多项式、指数和幂函数等对株高与产量要素间的定量关系进行拟合,并通过比较拟合精度的显著性值大小来确定最佳拟合模型。
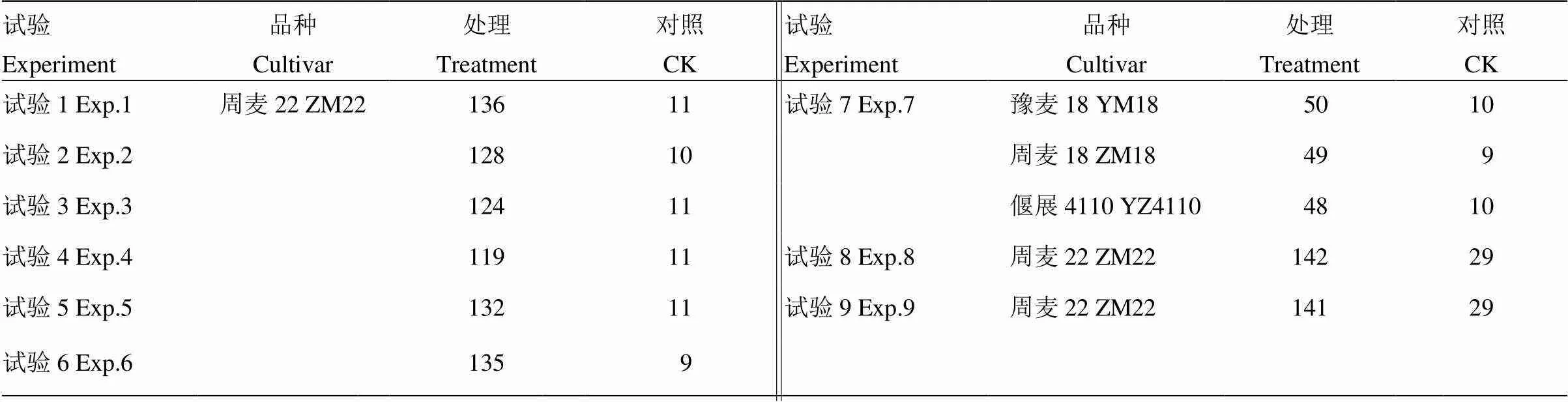
表4 株高及其构成因素数据的样本量
1.3.3 突变检验 在确定株高随处理温度呈显著线性降低的前提下,进一步对其是否存在突变进行检验。在此,引入气候突变研究中常用的Mann-Kendall(M-K)检验法。该方法优点在于不需要样本遵从一定的分布,也不受个别异常值的干扰。其设计思路在于检测时间序列上参量的突变性,每个时间点对应一个样本,即“一对一”的关系。本研究若以处理温度作为参考序列,则在同一处理温度下包含多个株高样本,即“一对多”的关系。因此,仅利用M-K检验法来检测株高随处理温度变化的突变点是不可行的。为解决这一问题,我们采用M-K检验和高阶函数拟合相结合的办法。具体计算步骤如下:
首先,对于具有个总样本量的处理温度序列,构造一秩序列:
(1)
(2)
式中,为株高,S表示第个样本x>x的累计值,[S]为均值,var[S]为方差。
其次,计算统计量:
(3)
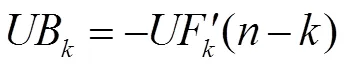
先利用式(3)计算原始处理温度序列下所有样本值的统计量UF,接着将该序列的样本值逆序排列,并按照式(2,3)计算得到逆序列的统计量UF′,再根据式(4)计算UB。给定显著性水平= 0.05,当统计量的绝对值大于1.96时,表示通过了显著性检验。
最后,高阶函数拟合与突变检验:
构建所有样本的UF、UB与处理温度间的散点关系图,利用高阶函数分别对其进行曲线拟合。经对拟合精度进行0.05显著性水平的检验后,最终得到两条统计量曲线。如果UF和UB曲线出现一个交点且在统计量为±1.96的两条临界直线之间(信度区间),则该交点处表示突变开始。如果交点超出信度区间,则采用Yamamoto方法来计算信噪比(),以判断交点前后2个子序列的均值是否有显著差异[20]。
(5)
2 结果
2.1 株高变化
以各期试验开展时冬小麦所处发育进程为序,统计各低温处理下的平均株高,结果如图1和图2所示。可以看出,所有试验的平均株高均在最低处理温度(-11℃或-9℃)下达到最小值,且与对照间表现为显著差异(仅图1-E除外)。其中,试验2(图1-B)、试验3(图1-C)、试验4(图1-D)、试验7(图2-H)和试验8(图2-G)随着处理温度的降低,株高均呈极强显著线性降低趋势(<0.001)。在试验2、试验3和试验4中,对照株高为73.4、70.7和66.9 cm,-9℃处理下的株高为72.5、68.2和62.8 cm,株高降幅分别为1.3%、3.6%和6.1%;在-11℃处理下的株高为70.2、65.8和56.1 cm,株高降幅分别达4.4%、7.0%和16.2%。在试验8和试验7中,株高在-9℃处理下的平均降幅分别达7.5%和27.9%。就不同品种来看,试验7中豫麦18、周麦18和偃展4110的对照株高为65.7、68.6和69.0 cm,-9℃处理下株高为48.2、51.3和47.0 cm,株高降幅分别达26.6%、25.2%和31.9%。多重比较结果表明(图1—2),试验2、试验3、试验4、试验7和试验8中株高的降低特性是在处理温度降至某一临界温度以下时,才显著开始的。从各项试验结果来看,临界处理温度多集中在-5℃(图1-C、图2-G、图2-H-2和图2-H-3)或-7℃(图1-B和图1-D),品种豫麦18甚至在低于-3℃时开始显著降低(图2-H-1)。与之相对应,株高在-9℃至临界处理温度区间的变异系数明显大于临界处理温度至-1℃区间的变异系数,二者之差在试验2、试验3和试验4中依次为0.026、0.028和0.079,在试验8及试验7的3个品种中依次为0.018、0.080、0.022和0.104。总之,不论是基于低温室还是移动霜箱的霜冻模拟试验,株高在幼穗发育进入雌雄蕊原基分化至药隔形成时期,均呈现出显著降低特性;在不同发育进程、且同一处理温度下,株高降幅随发育进程推移而明显增大,个体差异也越发明显;在相同发育进程、且同一处理温度下,基于移动霜箱的模拟冻害比低温室造成了更大的株高降幅;随着处理温度降低,株高会出现非渐变式(即突变性)降低特征,且不同植株个体、品种间的株高降低差异更加明显。

A―F表示试验1—试验6。误差线表示50倍的变异系数。短划线表示株高随处理温度降低的线性拟合直线,小写字母表示株高在各处理温度间的差异(α=0.05),图2、图5、图6同
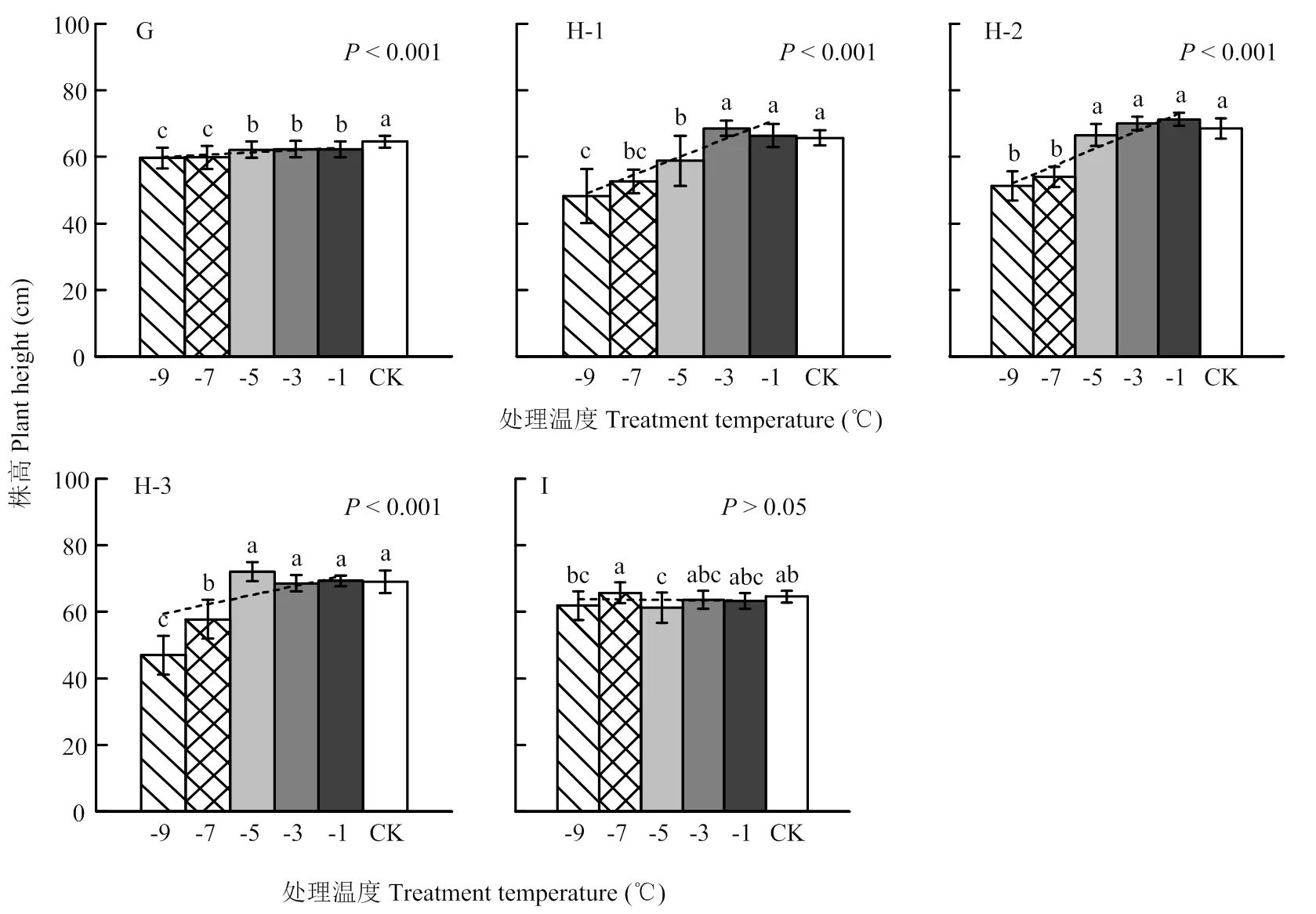
G、H和I分别表示试验8、试验7和试验9,H-1,H-2和H-3分别表示冬小麦品种豫麦18、周麦18和偃展4110。误差线表示50倍的变异系数
G, H and I represent Exp.8, Exp.7 and Exp.9, and H-1, H-2 and H-3 represent cultivars Yumai18, Zhoumai18 and Yanzhan4110, respectively. Error bar represents the 50-time coefficient of variation
图2 基于移动霜箱模拟的晚霜冻胁迫对株高的影响
Fig. 2 Effect of late frost stress on plant height based on the Field Movable Climate Chamber simulation
为进一步检验株高降低是否存在突变特性,在此利用M-K突变检验方法,分别计算试验2、试验3、试验4、试验7和试验8中株高的M-K统计量,并对统计量进行不同阶次的函数拟合。经比较,发现低温室试验和移动霜箱试验在函数阶次分别达到5阶和4阶时,其精度不仅达到0.001显著性水平,而且决定系数与显著性值不再随阶次的升高而发生明显变化。据此,绘制M-K统计量随处理温度变化的拟合曲线,如图3—4所示。可以看出,UF和UB统计量的两条拟合曲线在各试验中均出现一个交点。其中,试验2(图3-B)、试验3(图3-C)、试验8(图4-G)、试验7豫麦18(图4-H-1)和偃展4110(图4-H-3)中的交点均处于信度区间,表明株高在该点处开始突变。试验4(图3-D)和试验7周麦18(图4-H-2)中的交点均位于信度区间之外,其值分别为0.5(<1.0)和2.1(>2.0),据此认为试验4中株高没有发生突变现象,而试验7周麦18的株高则发生了强突变。如将两条统计量拟合曲线交点处的温度称作开始突变温度,则除试验7偃展4110为-7.1℃外,其他试验中株高的开始突变温度均处于-4.6℃—-5.4℃区间。比较移动霜箱试验中的3个小麦品种(图4-H),发现偃展4110的开始突变温度(-7.1℃)明显低于豫麦18(-5.4℃)和周麦18(-5.2℃),这可能表明各品种的抗冻性间存在一定差异。总体来看,不管是基于低温室还是移动霜箱的霜冻模拟试验,冬小麦株高的降低趋势均存在着突变特性。
2.2 株高构成因素变化
针对冬小麦不同发育进程开展的低温室霜冻模拟试验表明(图5),株高构成因素随处理温度降低而变化的趋势与其伸长规律联系密切。在小花原基分化期(图5-A),正值倒五节间(本研究未记录其长度)初伸长、倒四节间待伸长阶段,此时倒四节间长度因霜冻胁迫而强显著缩短,其他因素变化均不显著(>0.05)。在雌雄蕊原基分化期(图5-B),为倒四节间初伸长、倒三节间待伸长阶段,此时倒三节间和穗长因霜冻胁迫而呈极强显著缩短趋势,倒二节间显著缩短,倒四节间和穗下节间长度变化不明显。进入药隔形成期(图5-C,图5-D),倒三节间和倒二节间先后初伸长,至药隔期末时倒四节间伸长结束,该时期倒四节间、倒三节间、倒二节间长和穗长均因霜冻胁迫而呈缩短趋势,其中除倒三节间长在药隔形成前期(图5-C)不显著外,其他均至少达到强显著水平(<0.01);穗下节间长度变化不明显。在四分体形成期(图5-E),处于穗下节间初伸长阶段,至四分体期末时倒三节间和穗伸长结束,该时期仅倒三节间长在霜冻胁迫下显著缩短,穗下节间长则表现出强显著延长趋势。冬小麦抽穗时(图5-F),正值穗下节间快速伸长阶段,至开花前倒二节间伸长结束,此时霜冻胁迫仅造成穗下节间显著缩短,倒四节间和麦穗则出现延长趋势。

B、C和D分别表示试验2、试验3和试验4 B, C and D represent Exp.2, Exp.3 and Exp.4, respectively

G和H分别表示试验8和试验7,H-1,H-2和H-3分别表示冬小麦品种豫麦18、周麦18和偃展4110
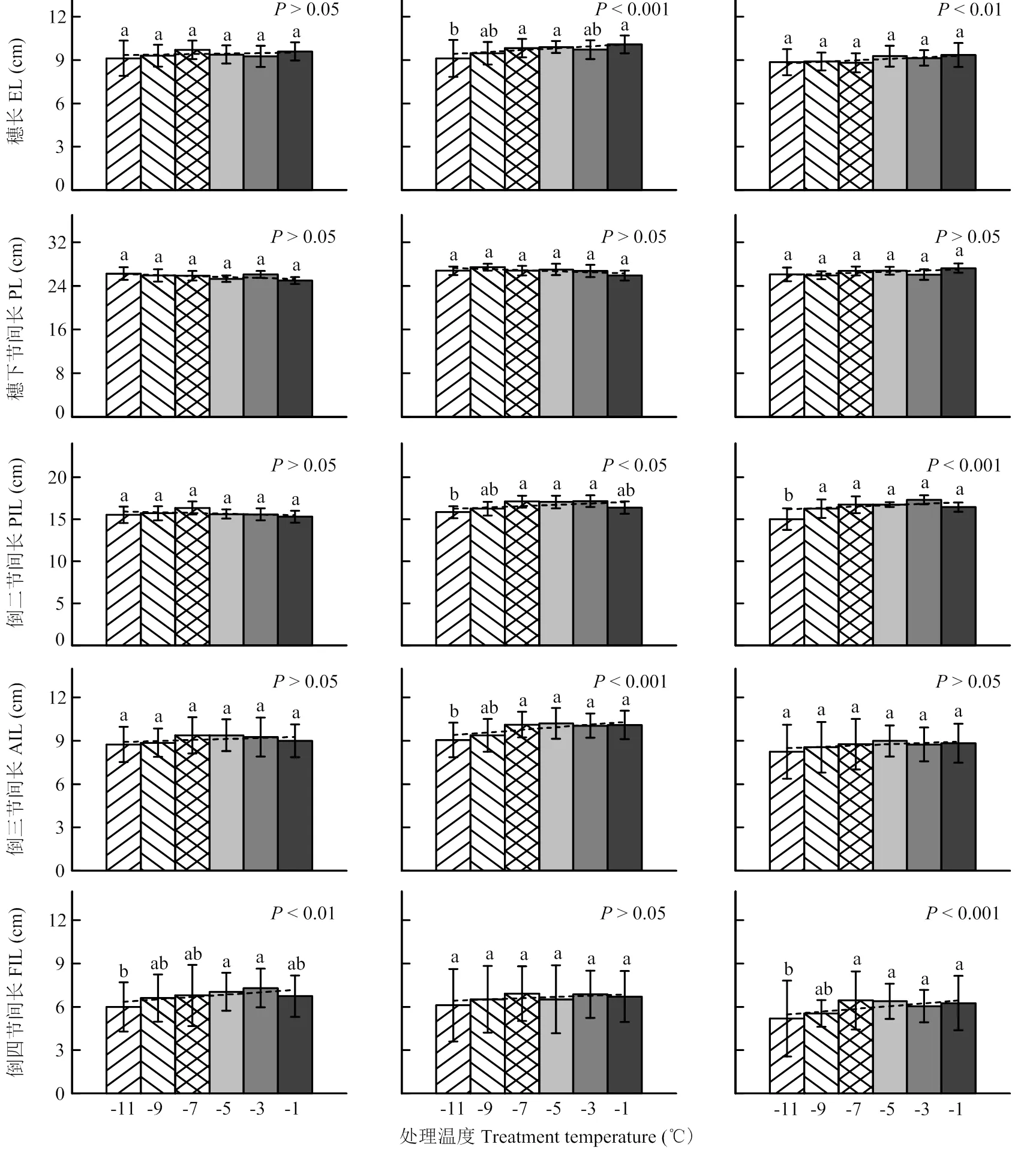
移动霜箱试验仅考察了穗长、穗下节间长和倒二节间长,其随处理温度降低的变化趋势如图6所示。在雌雄蕊原基分化期(图6-G),穗长和倒二节间长因霜冻胁迫而强显著缩短,穗下节间长则极强显著延长。在药隔形成后期(图6-H),穗长、穗下节间长、以及其中2个品种(豫麦18除外)的倒二节间长均因霜冻胁迫而呈极强显著缩短趋势。在四分体时期(图6-I),除穗长强显著缩短外,穗下节间和倒二节间变化均不明显。总体来看,不论是低温室试验还是移动霜箱试验,植株节间长(穗下节间除外)、穗长在各发育进程霜冻胁迫下大体保持了一致的缩短规律,即显著缩短趋势仅发生在其伸长或者待伸长阶段,伸长结束后则无此现象。
比较试验1—试验6中株高构成因素与处理温度的相关系数(表5),可以看出倒四节间长与处理温度间的正相关性在试验1和试验4中最为显著,在试验3中的正相关性仅次于倒二节间长;倒三节间长和处理温度间的正相关性则在试验2和试验5中最显著。这表明冬小麦抽穗前晚霜冻胁迫对倒四节间长和倒三节间长的总体影响是最大的。
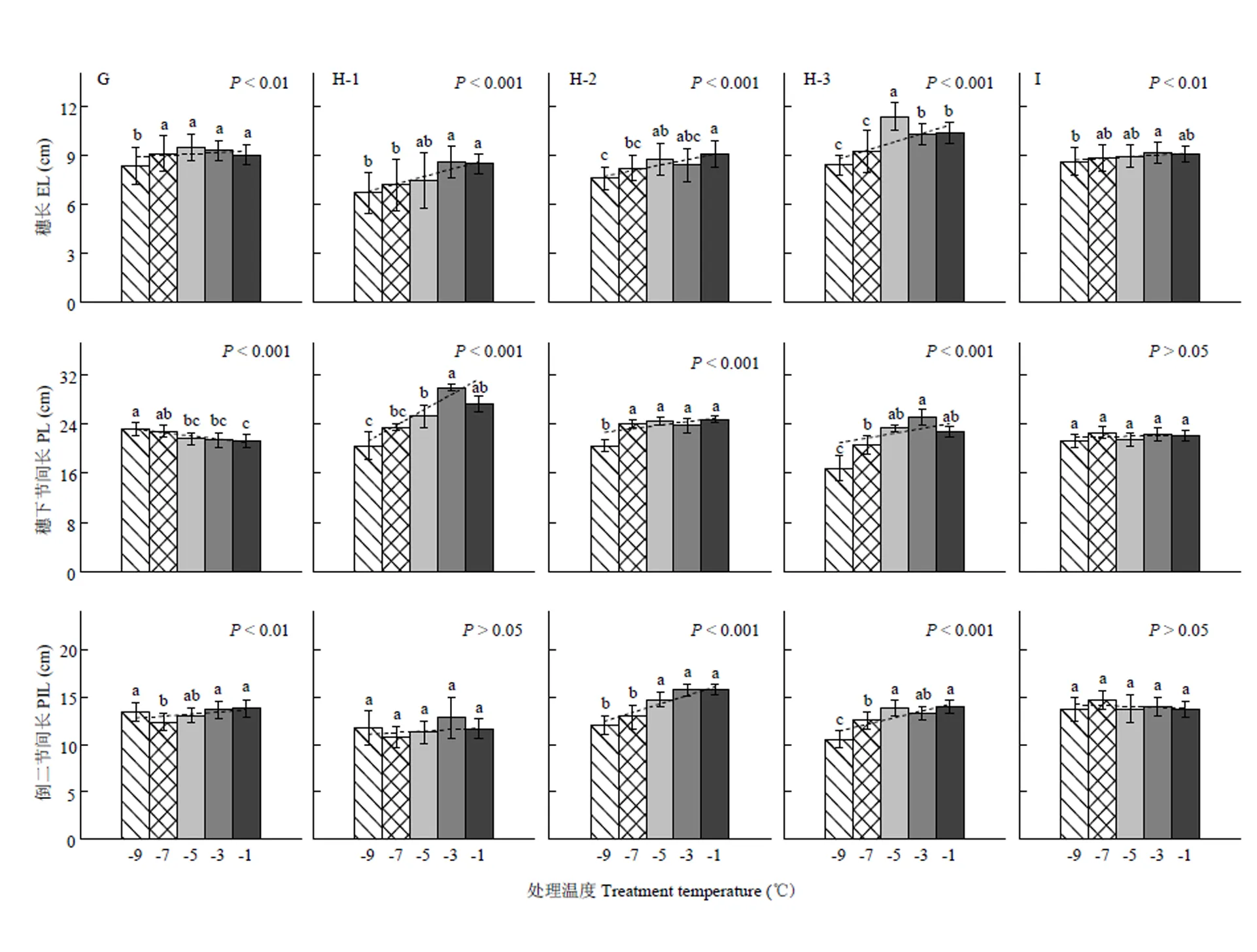
G、H和I分别表示试验8、试验7和试验9,H-1,H-2和H-3分别表示冬小麦品种豫麦18、周麦18和偃展4110。误差线表示10倍的变异系数

表5 株高构成因素与处理温度的相关系数
*,**和***分别表示在0.05、0.01和0.001显著性水平下的相关性,表6同。各试验中相关性的样本量如表4所示
*, ** and *** indicate the correlations at significance levels of 0.05, 0.01 and 0.001 respectively, the same as Table 6. Sample size in each experiment has been shown in Table 4
2.3 株高与其构成因素的关系
为探究各构成因素在不同低温处理和发育进程下的缩短特性与株高是否存在密切联系,对其相关性大小、方向及显著性水平进行了统计,如表6所示。可以看出,倒四节间、倒三节间、倒二节间长度以及穗长与株高的相关系数在各试验中均至少达到显著正相关(<0.05);除试验4和试验7(这两个试验均是在药隔形成后期开展的)外,其他试验中穗长与株高的相关系数均明显低于各节间和株高的相关系数;穗下节间长和株高的相关系数除在试验4、试验6、试验7和试验9达到显著正相关外,在其她试验中均无显著正相关性。可见,不论株高在晚霜冻后是否出现显著降低特性,穗下节间除外的其他各构成因素均与株高保持了显著正相关关系。

表6 晚霜冻胁迫下株高与其构成因素的相关系数
进一步基于低温室试验数据,利用逐步回归建模方法将株高构成因素(穗长、穗下节间长、倒二节间长、倒三节间长和倒四节间长)作为自变量,以构建其与株高间的逐步回归模型,所有模型均通过0.001显著性精度检验。接着计算所有参量因子的直接通径系数,结果如表7所示。为保持各期低温室试验中处理温度范围的一致性,均使用-1℃—-9℃处理的数据。结果表明,除试验1仅有倒二节间长和倒三节间长入选回归模型外,其他试验中各构成因素均入选模型;所有入选因子对株高的影响均呈正效应。如果在试验1中加入-11℃处理,则全部构成因素均能选入模型,这也表明该时期植株的抗冻性明显高于其他各期试验。即使处理温度降低至-11℃,模型中直接通径系数若按大小排序,排前两位的仍为倒二节间长(0.440)和倒三节间长(0.348)。可见,该时期这两个节间长度对株高的影响最大。在试验2—试验4中,直接通径系数最大的依次为倒四节间长(0.529)、倒三节间长(0.488)和倒二节间长(0.426),对株高影响最大的节间刚好与其初伸长时间相对应。试验5—试验6中直接通径系数最大的均为倒二节间长(分别达0.534和0.571),接下来依次是倒四节间长、穗下节间长、倒三节间长和穗长。除了试验4中穗长对株高的影响超过穗下节间长外,在其他试验中穗长的影响均为最小。进一步分析可知,试验1中仅倒四节间显著缩短,此时对株高起影响作用的主要是倒三节间和倒二节间,而这两个节间长度的变化趋势均不显著,因此该时期株高变化不显著;在试验2—试验4中,倒四节间、倒三节间和倒二节间的长度均表现为不同程度的显著缩短趋势,这3个阶段对株高的影响排前两位的分别为倒四节间和倒三节间、倒三节间和倒二节间、倒二节间和倒四节间,可见该时期对株高贡献最大的节间同时也是长度显著缩短的节间,因此株高呈显著降低特性;在试验5和试验6中,仅倒三节间和穗下节间分别表现为显著缩短趋势,这两个节间对株高的影响均明显低于倒二节间和倒四节间,因此该时期株高变化不明显。
2.4 株高与籽粒产量的关系
为揭示晚霜冻胁迫下株高降低特性对产量变化的指示意义,本研究统计了试验2、试验3、试验4、试验7和试验8中株高与穗粒数、千粒重、单株产量的相关系数,并且发现株高与穗粒数、千粒重、单株产量间的相关性在所有试验中均至少达到0.01显著水平;相比较而言,株高与单株穗数间的相关性较差,其中试验7中豫麦18和周麦18、试验8中周麦22这3个品种均未达到显著相关(>0.05)。
分别合并低温室模拟(试验2、试验3和试验4)和移动霜箱模拟(试验7和试验8)的霜冻试验数据,构建株高与穗粒数、千粒重、单株产量间的散点关系,并利用线性、二次多项式、指数和幂函数等对其散点关系进行拟合,发现幂函数拟合精度的置信水平都最高,因此作为最终优选函数,其拟合曲线如图7所示。在低温室霜冻模拟条件下,株高与穗粒数、单株产量、千粒重间的幂函数拟合精度均达到0.001显著性(图7-A);在移动霜箱霜冻模拟条件下,株高与穗粒数(图7-B1)、单株产量(图7-B2)的幂函数拟合精度达到0.001显著性,株高与千粒重(图7-B3)的拟合精度达到0.01显著性。对基于低温室模拟和移动霜箱模拟的两类霜冻试验进行比较(图7-A和图7-B),发现其既有相同之处,又有不同之处。相同之处在于:(1)株高与千粒重间的拟合精度均明显低于株高与穗粒数、单株产量间的拟合精度;(2)随着株高降低,穗粒数、单株产量和千粒重均呈降低趋势,其中单株产量的降幅由大变小,直至变化不甚明显;(3)与药隔形成期相比,雌雄蕊原基分化期霜冻胁迫后的株高、穗粒数、单株产量和千粒重的值相对更高,分布也更为集中(如图7-A中绿色正方形符号和图7-B中粉色倒三角形符号所示)。不同之处在于:(1)在移动霜箱霜冻模拟试验中,株高与穗粒数、单株产量、千粒重间的幂函数拟合精度比低温室霜冻模拟试验相对较低;(2)在低温室霜冻模拟试验中,穗粒数降幅随株高降低亦呈现由大变小的态势,但在移动霜箱霜冻模拟试验中无此规律。
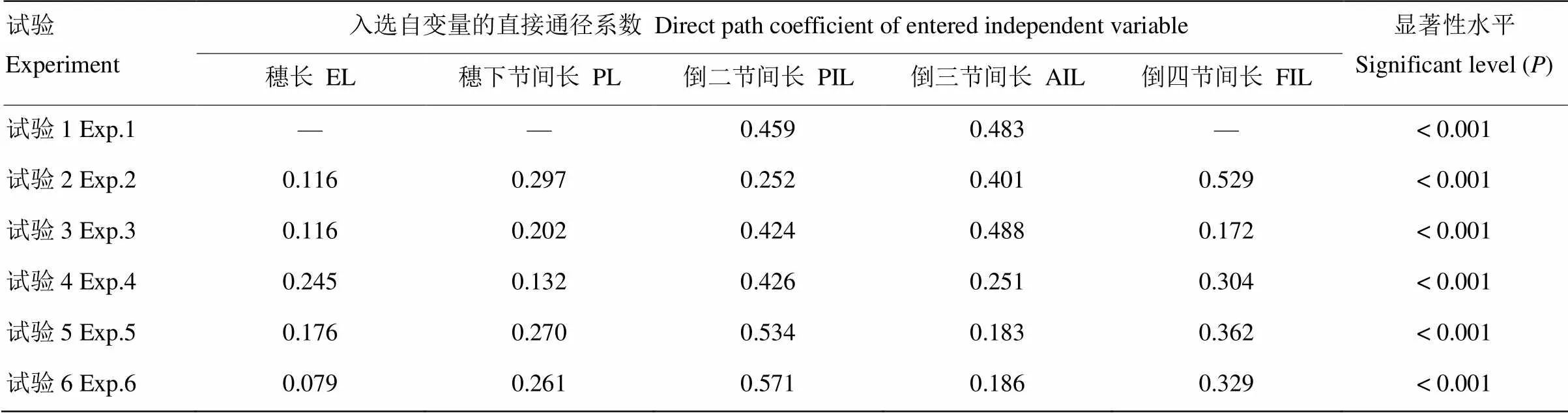
表7 逐步回归模型中各株高构成因素的直接通径系数

黑色实线为基于幂函数的拟合曲线,n为样本量 Black solid line presents a fitting curve based on the power function, and n is the sample size
3 讨论
3.1 株高降低特性及其限定条件
关于晚霜冻胁迫下冬小麦株高的降低特性,在已有研究中取得了一定共识。这些研究从发育进程、降温强度、品种与熟性等限定条件入手,认为不同小麦品种的株高在设定低温处理(如-2℃或-3℃)下,随着胁迫时间(如24、48或72 h)延长而显著降低,且多以药隔期之后研究为主[21-22]。针对大田自然晚霜冻的考察结果也表明,株高随冻害程度加深而明显降低[4, 23]。尽管模拟霜冻研究与自然霜冻研究结论相一致,但是利用超过20 h的低温胁迫时长作为冻害程度控制指标,显然不符合单次自然霜冻过程中的实际降温时长规律。为此,本研究从遵循我国黄淮麦区晚霜冻实际发生时间、降温幅度与时长以及当地主栽品种等的现实性出发,在开展霜冻模拟试验时充分考虑了5点设计原则。一是将试验地点放在晚霜冻频发的河南省商丘地区,以确保冬小麦整个生育期处在当地农田生长环境中。二是试验日期涵盖了拔节后幼穗分化的各个阶段。其中,基于低温室模拟的霜冻试验共开展了6期(试验1—试验6),从小花原基分化开始直至抽穗时结束,历时28 d;基于移动霜箱模拟的霜冻试验共开展了3期(试验7—试验9),从雌雄蕊原基分化至四分体形成时结束。此处必须说明的是,尽管试验7和试验9的时间仅相差1 d,但试验7开展时幼穗分化正处于药隔形成后期,而试验9时已进入四分体形成期,这可能是由于试验9冬小麦播种时间更早,且2014年3月至4月间平均气温比2012年同期高出1.5℃,其发育进程更快。三是设定包括-1℃、-3℃、-5℃、-7℃、-9℃和-11℃在内的处理温度序列,并利用温控系统模拟自然降温过程(如前述1.1节所示),使得每次试验过程均能造成植株由未见受冻到明显死亡的各种程度冻害。四是从当地主栽品种中挑选了2个半冬性中熟品种和2个弱春性早熟品种,以期探究品种间差异。五是利用田间可移动式霜箱和低温室2种手段,分别在农田原位(小区种植)和室内(盆栽种植)进行晚霜冻模拟试验,试验结果用于相互佐证株高的降低特性。由于移动霜箱试验的小麦田块位于城区边缘,热岛效应明显,而低温室试验的小麦田块位于野外,因此移动霜箱试验小麦的发育进程要早于低温室试验小麦。此外,在同一处理温度下,移动霜箱会比低温室造成更大的小麦冻害损伤,这可能与其降温空间狭小有关。
基于以上限定条件开展晚霜冻模拟试验的结果表明,在雌雄蕊原基分化至药隔形成期,两类品种的株高均随处理温度降低而呈显著降低趋势,并且在低于-4.6℃—-5.4℃区间时出现明显突变现象。突变现象表明冬小麦植株的抗冻能力存在一定阈值区间。AL-ISSAWI等[24]和Fuller等[25]研究认为在二棱期/小花原基分化之后,小麦植株可以抵抗-5℃以上冠层低温的侵袭而不受冻,Woodruff等[9]和Livingston等[26]则认为冠层温度低于-6℃即可使幼穗和茎秆发生明显伤害症状。不论是-5℃还是-6℃低温,都恰恰与本研究中株高降低的突变温度区间相接近。本研究对株高显著降低的试验2、试验3、试验4、试验7和试验8中植株茎部冻害进行考察的结果也表明,在-1℃—-5℃处理下,植株叶片和茎节均未见明显外在冻害症状;在-7℃处理下,茎、叶冻害症状出现;在-9℃—-11℃处理下,茎、叶冻害比重明显增加。
3.2 节间缩短对株高降低特性的贡献
株高构成因素在冬小麦发育的不同时期伸长,共同影响株高性状[27],进而关系到小麦产量变化[28]。因此,揭示晚霜冻胁迫下株高降低特性与规律,离不开对其构成因素变化的分析。刘立伟等[22]认为药隔前期低温胁迫下倒四节间、倒五节间和穗的长度缩短明显,倒三节间,倒二节间和穗下节间长度缩短相对较不明显;Whaley等[4]的研究结论则表明,药隔后期冻害主要阻碍了穗下节间和倒二节间的伸长,其次是倒三节间。可见,晚霜冻害主要对伸长期或者待伸长期的节间与穗的长度构成影响,并导致其显著缩短,本研究结论也清楚地表明了这一点。但是从试验1—试验9结果来看,节间因冻害缩短现象并非总与株高降低保持同步。试验1、试验5和试验6开展时对应伸长的倒四节间、倒三节间和穗下节间均出现缩短现象,但是株高并未显著降低。因此,株高降低与否还需考虑各构成因素对其综合影响。正如图5和表7结果所示,只有因冻害缩短的节间与对株高起决定性贡献的节间保持一致时,株高才表现为显著降低特性。另外,经比较发现,低温室试验和移动霜箱试验中穗下节间长的变化差异较大。在低温室试验中,穗下节间长在药隔期变化不明显(图5-F),与刘立伟等[22]研究结论一致;在移动霜箱试验中,穗下节间长在药隔期表现为极强显著缩短趋势(图6-H),与Whaley等[4]研究结论一致。在两类试验中,穗下节间长在四分体时期变化均不显著,而该时期正是穗下节间初伸长阶段,似乎应该出现缩短现象。究其原因,就目前试验结果还无法作出进一步合理解释,因此有必要开展更多类似试验以完善相关研究结论。
3.3 节间缩短与幼穗冻害对产量的共同影响
株高与穗粒数、千粒重、单株产量之间的显著相关性表明,利用株高降低幅度作为评判指标,可提早预测雌雄蕊原基分化至药隔形成期霜冻胁迫下的冬小麦产量损失风险。从株高与穗粒数、千粒重、单株产量间的幂函数拟合曲线(图7)来看,最大亮点在于单株产量降幅随株高降低而呈现出先大后小态势。当单株产量降低至约1.5 g以下时,不会再随株高继续降低而明显降低,主要原因可能在于此时的穗粒数变化已不大。总体上看,基于低温室试验数据拟合的精度要明显高于移动霜箱试验数据,这可能是由于低温室试验仅使用了1个品种,而移动霜箱试验中涉及4个品种,其株高范围和抗冻能力有所不同。
若要探明节间缩短与幼穗冻害对产量的影响机制,需先对晚霜冻胁迫下冬小麦茎、叶和幼穗冻害规律有一个清晰的认识。我们在前面探讨了雌雄蕊原基分化至药隔形成期株高降低特性的突变温度区间,冠层气温低于该温度区间时茎和幼穗均因结冰而发生显著冻害。LIVINGSTON Ⅲ等[29]研究认为植株结冰总是先从茎的基部开始,并逐渐向上扩展至叶片,进而至幼穗。发生冻害的植株会迅速死亡,以致无法形成结实穗。但并非冠层气温低于该温度区间时植株就会结冰并死亡,此时仍有相当一部分植株会因处于过冷却状态而恢复生长,并最终形成不完全结实的穗。Frederiks等[14]则认为,即使冠层气温高于该温度区间(此时茎部看上去完好无损),包在叶鞘内部的幼穗也仍有可能发生冻害,并会形成不完全结实的穗。可见,在高于突变温度区间的晚霜冻胁迫下,茎节通常不会受冻,因而株高降低不明显(图1,图2),此时籽粒产量损失原因主要表现为穗粒数的下降;在低于突变温度区间的晚霜冻胁迫下,不仅穗粒数下降,群体成穗数也会因部分植株的死亡而显著降低[4, 30],这从试验2、试验3和试验4的结果也可以看出来,与对照(总株数均为48)相比,茎蘖死亡率在-7℃处理下分别为0、39.6%和41.7%,在-9℃处理下分别为27.1%、35.9%和43.8%,在-11℃处理下分别为31.3%、43.8%和45.8%。过冷却的植株恢复生长后,株高显著降低,原因在于对其起关键影响作用的节间发生了显著缩短现象。节间缩短的同时也会造成籽粒发育受损,进而影响产量。这主要表现在两个方面:一是茎的营养传导功能受限[12],导致籽粒皱缩、粒重降低[14];二是受冻茎秆相比正常而言变得更为纤弱,因而容易在生长后期发生倒伏[31]。研究表明[32-33],小麦倒伏一般发生在茎的基部第二节间(倒四节间)和第三节间(倒三节间)。这一结论恰好与表5所示结果相对应,即受晚霜冻影响最大的节间亦是最易发生倒伏的节间。可见,利用倒四节间长或倒三节间长的缩短规律来评估/预测晚霜冻胁迫下冬小麦倒伏风险与产量损失,具有潜在应用价值。
4 结论
在模拟晚霜冻条件下,正在伸长或待伸长的冬小麦节间长度与穗长随处理温度降低而显著缩短;当缩短节间与影响株高的关键节间相一致时,株高则降低。在雌雄蕊原基分化至药隔形成期,倒四节间长、倒三节间长和倒二节间长均显著缩短,并且对株高的总体贡献最大,因此株高表现出显著降低趋势;当处理温度低于-5℃左右时,部分植株的茎部因出现明显冻害症状而死亡,存活植株的整体高度急剧降低,株高在该温度值附近存在突变特性,且不同品种间有一定差别。株高与穗粒数、千粒重、单株产量之间存在幂函数曲线关系,当单株产量因冻害胁迫而降低至1.5 g以下时,随株高继续降低而不再明显减少,此时穗粒数变化也已不大。
[1] ZHANG N, FAN X L, CUI F, ZHAO C H, ZHANG W, ZHAO X Q, YANG L J, PAN R Q, CHEN M, HAN J, JI J, LIU D C, ZHAO Z W, TONG Y P, ZHANG A M, WANG T, LI J M. Characterization of the temporal and spatial expression of wheat (L.) plant height at the OTL level and their influence on yield-related traits., 2017, 130(6): 1235-1252.
[2] 李春燕, 杨景, 张玉雪, 姚梦浩, 朱新开, 郭文善. 小麦分蘖期冻害后增施恢复肥的产量挽回效应及其生理机制. 中国农业科学, 2017, 50(10): 1781-1791.
LI C Y, YANG J, ZHANG Y X, YAO M H, ZHU X K, GUO W S. Retrieval effects of remedial fertilizer after freeze injury on wheat yield and its mechanism at tillering stage., 2017, 50(10): 1781-1791. (in Chinese)
[3] FOWLER D B, N’DIAYE A, LAUDENCIA-CHINGCUANCO D, POZNIAK C J. Quantitative trait loci associated with phenological development, low-temperature tolerance, grain quality, and agronomic characters in wheat (L)., 2016, 11(3): e0152185.
[4] WHALEY J M, KIRBY E J M, SPINK J H, FOULKES M J, SPARKES D L. Frost damage to winter wheat in the UK: the effect of plant population density., 2004, 21(1): 105-115.
[5] MAHFOOZI S, LIMIN A E, AHAKPAZ F, FOWLER D B. Phenological development and expression of freezing resistance in spring and winter wheat under field conditions in northwest Iran., 2006, 97(2): 182-187.
[6] WU Y F, ZHONG X L, HU X, REN D C, LÜ G H, WEI C Y, SONG J Q. Frost affects grain yield components in winter wheat., 2014, 42(3): 194-204.
[7] 胡新, 任德超, 倪永静, 黄建英, 宋吉青, 武永峰. 冬小麦籽粒产量及其构成要素随晚霜冻害变化规律研究. 中国农业气象, 2014, 35(5): 575-580.
HU X, REN D C, NI Y J, HUANG J Y, SONG J Q, WU Y F. Impacts of late frost on grain yield and its components of winter wheat., 2014, 35(5): 575-580. (in Chinese)
[8] ZHANG J N, HAO C Y, REN Q, CHANG X P, LIU G R, JING R L. Association mapping of dynamic developmental plant height in common wheat., 2011, 234(5): 891-902.
[9] WOODRUFF D, DOUGLAS N, FRENCH V.. Queensland, Brisbane: Department of Primary Industries, 1997.
[10] CROMEY M G, WRIGHT D S C, BODDINGTON H J. Effects of frost during grain filling on wheat yield and grain structure., 1998, 26(4): 279-290.
[11] 武永峰, 胡新, 钟秀丽, 吕国华, 任德超, 宋吉青. 农田尺度下冬小麦晚霜冻害空间差异及原因分析. 中国农业科学, 2014, 47(21): 4246-4256.
WU Y F, HU X, ZHONG X L, LÜ G H, REN D C, SONG J Q. Study on spatial differences of late frost injury to winter wheat and its reasons at field scale., 2014, 47(21): 4246-4256. (in Chinese)
[12] SHROYER J P, MIKESELL M E, PAULSEN G M.. Manhattan: Kansas State University, 1995.
[13] FREDERIKS T M, CHRISTOPHER J T, FLETCHER S E H, BORRELL A K. Post head-emergence frost resistance of barley genotypes in the northern grain region of Australia., 2011, 62(9): 736-745.
[14] FREDERIKS T M, CHRISTOPHER J T, SUTHERLAND M W, BORRELL A K. Post-head-emergence frost in wheat and barley: defining the problem, assessing the damage, and identifying resistance., 2015, 66(12): 3487-3498.
[15] 任德超, 胡新, 陈丹丹, 张建涛, 倪永静, 刘红杰, 黄绍华, 李国强. 不同低温处理对小麦光合特性和产量性状的影响. 中国农学通报, 2016, 32(21): 44-50.
REN D C, HU X, CHEN D D, ZHANG J T, NI Y J, LIU H J, HUANG S H, LI G Q. Effects of different low temperature treatments on photosynthetic characteristics and yield traits of wheat., 2016, 32(21): 44-50. (in Chinese)
[16] 冯玉香, 何维勋, 孙忠富, 钟秀丽. 我国冬小麦霜冻害的气候分析. 作物学报, 1999, 25(3): 335-340.
FENG Y X, HE W X, SUN Z F, ZHONG X L. Climatological study on frost damage of winter wheat in China., 1999, 25(3): 335-340. (in Chinese)
[17] 张雪芬, 郑有飞, 王春乙, 陈怀亮, 任振和, 邹春辉. 冬小麦晚霜冻害时空分布与多时间尺度变化规律分析. 气象学报, 2009, 67(2): 321-330.
ZHANG X F, ZHENG Y F, WANG C Y, CHEN H L, REN Z H, ZOU C H. Spatial-temporal distribution and multiple-temporal scale variation analyses of winter wheat late freezing injury., 2009, 67(2): 321-330. (in Chinese)
[18] 武永峰, 胡新, 吕国华, 任德超, 蒋卫国, 宋吉青. 晚霜冻影响下冬小麦冠层红边参数比较. 光谱学与光谱分析, 2014, 34(8): 2190-2195.
WU Y F, HU X, LÜ G H, REN D C, JIANG W G, SONG J Q. Comparison of red edge parameters of winter wheat canopy under late frost stress., 2014, 34(8): 2190-2195. (in Chinese)
[19] 张朝生, 章申, 张立成, 王立军. 长江水系河流沉积物重金属元素含量的计算方法研究. 环境科学学报, 1995, 15(3): 257-264.
ZHANG C S, ZHANG S, ZHANG L C, WANG L J. Calculation of heavy metal contents in sediments of the Changjiang river system., 1995, 15(3): 257-264. (in Chinese)
[20] 柴元方, 李义天, 李思璇, 朱博渊, 王靖宜. 长江流域近期水沙变化趋势及成因分析. 灌溉排水学报, 2017, 36(3): 94-101.
CHAI Y F, LI Y T, LI S X, ZHU B Y, WANG J Y. Analysis of recent variation trend and cause of runoff and sediment load variations in the Yangtze river basin., 2017, 36(3): 94-101. (in Chinese)
[21] 陈贵菊, 陈明丽, 王福玉, 高国良, 江涛, 尹逊利, 李根英, 宋国琦. 药隔期低温对小麦生长发育的影响. 山东农业科学, 2015, 47(2): 25-28.
CHEN G J, CHEN M L, WANG F Y, GAO G L, JIANG T, YIN X L, LI G Y, SONG G Q. Influences of low temperature in connectivum period on wheat growth and development., 2015, 47(2): 25-28. (in Chinese)
[22] 刘立伟. 药隔、减数分裂、开花期低温对小麦产量的影响与补救研究[D]. 扬州: 扬州大学, 2016.
LIU L W. Effects of low temperature at anther connective formation, meiosis and anthesis stages on grain yield in wheat and remedial techniques[D]. Yangzhou: Yangzhou university, 2016. (in Chinese)
[23] 靖华, 亢秀丽, 马爱平, 崔欢虎, 王娟玲, 刘建华. 晋南旱垣春季低温对不同播种期小麦冻害的影响. 中国农学通报, 2011, 27(9): 76-80.
JING H, KANG X L, MA A P, CUI H H, WANG J L, LIU J H. Effect of spring low temperature on different sowing date winter wheat frozen injury on the arid area of southern Shanxi province., 2011, 27(9): 76-80. (in Chinese)
[24] AL-ISSAWI M, RIHAN H Z, EL-SARKASSY N, FULLER M P. Frost hardiness expression and characterisation in wheat at ear emergence., 2013, 199(1): 66-74.
[25] FULLER M P, FULLER A M, KANIOURAS S K, CHRISTOPHERS J, FREDERICKS T. The freezing characteristics of wheat at ear emergence., 2007, 26(4): 435-441.
[26] LIVINGSTON Ⅲ D P, TUONG T D, ISLEIB T G, MURPHY J P. Differences between wheat genotypes in damage from freezing temperatures during reproductive growth.2016, 74: 164-172.
[27] 陈广凤, 陈建省, 田纪春. 小麦株高相关性状与SNP标记全基因组关联分析. 作物学报, 2015, 41(10): 1500-1509.
CHEN G F, CHEN J S, TIAN J C. Genome-wide association analysis between SNP markers and plant height related traits in wheat., 2015, 41(10): 1500-1509. (in Chinese)
[28] 朱新开, 郭文善, 李春燕, 封超年, 彭永欣. 小麦株高及其构成指数与产量及品质的相关性. 麦类作物学报, 2009, 29(6): 1034-1038.
ZHU X K, GUO W S, LI C Y, FENG C N, PENG Y X. Relationship of plant height component indexes with grain yield and quality in wheat., 2009, 29(6): 1034-1038. (in Chinese)
[29] LIVINGSTON Ⅲ D P, TUONG T D, MURPHY J P, GUSTA L V, WILLICK I, WISNIEWSKI M E. High-definition infrared thermography of ice nucleation and propagation in wheat under natural frost conditions and controlled freezing., 2018, 247(4): 791-806.
[30] JI H T, XIAO L J, XIA Y M, SONG H, LIU B, TANG L, CAO W X, ZHU Y, LIU L L. Effects of jointing and booting low temperature stresses on grain yield and yield components in wheat., 2017, 243: 33-42.
[31] BANATH C L, SINGLE W V. Frost injury to wheat stems and grain production., 1976, 27(6): 749-753.
[32] 刘凯, 邓志英, 张莹, 王芳芳, 刘佟佟, 李青芳, 邵文, 赵宾, 田纪春, 陈建省. 小麦茎秆断裂强度相关性状OTL的连锁和关联分析. 作物学报, 2017, 43(4): 483-495.
LIU K, DENG Z Y, ZHANG Y, WANG F F, LIU T T, LI Q F, SHAO W, ZHAO B, TIAN J C, CHEN J S. Linkage analysis and genome-wide association study of OTLs controlling stem-breaking- strength-related traits in wheat., 2017, 43(4): 483-495. (in Chinese)
[33] 胡昊, 李莎莎, 华慧, 孙蒙蒙, 康娟, 夏国军, 王晨阳. 不同小麦品种主茎茎秆形态结构特征及其与倒伏的关系. 麦类作物学报, 2017, 37(10): 1343-1348.
HU H, LI S S, HUA H, SUN M M, KANG J, XIA G J, WANG C Y. Research on stalk morphological structure characteristics and its relationship between the lodging of different wheat varieties., 2017, 37(10): 1343-1348. (in Chinese)
(责任编辑 杨鑫浩)
Reduction of Plant Height in Winter Wheat and Its Relationship with Grain Yield under Late Frost Stress
WU YongFeng1, HU Xin2, REN DeChao2, SHI Ping1, YOU SongCai1
(1Institute of Environment and Sustainable Development in Agriculture, Chinese Academy of Agricultural Sciences/Key Laboratory of Agricultural Environment, Ministry of Agriculture, Beijing 100081;2Wheat Research Institute, Shangqiu Academy of Agriculture and Forestry Sciences, Shangqiu 476000, Henan)
【Objective】 The objective of this study was to explore the effects of late frost stress on plant height and its components in winter wheat, and to clarify the characteristics of plant height reduction and its relationship with internode length, ear length and grain yield, in order to provide a basis for establishing the evaluation index of late frost damage. 【Method】By using two frost simulation means based on the Cold Climate Chamber and the Field Movable Climate Chamber, six pot experiments and three plots experiments were carried out with the developmental progresses (floret primordia differentiation (FPD), pistil and stamen primordia differentiation (PSPD), anther connective tissue formation (ACTF), tetrad formation (TF) and heading phase) of young ear and the subfreezing treatment temperatures (-1 °C, -3 °C, -5 °C, -7 °C, -9 °C and -11 °C) as the gradients. Culm injury was investigated after each frosting treatment, and plant height and its components, and grain yield were measured at maturity. Variance analysis, regression function fitting and test method for sudden change were used to study the rule of plant height reduction, the contribution of each component to plant height, and the regression relationship of plant height and grain yield. 【Result】 (1) In the stages of PSPD to late ACTF, plant height generally showed a decreasing trend with the decrease of treatment temperature. When the treatment temperature was lower than -5℃ or so, a mutation began, and there were certain differences between individual plants and varieties. At the same treatment temperature, the greatest decline in plant height occurred in the late ACTF stage. (2) In the stages of PSPD, early ACTF and late ACTF, fourth internode length from the top (FIL), antepenultimate internode length (AIL), penultimate internode length (PIL) and ear length (EL) all showed shortening trends in different degrees under the frost stress. In these periods, the key two internodes contributing to plant height were FIL and AIL, AIL and PIL, and PIL and FIL, respectively, that showed a strongly significant correlation (<0.001) with plant height. Correspondingly, plant height significantly decreased. (3) The regression curves between plant height and kernel number per ear, 1 000-kernel weight, and yield per plant were well fitted by the power function models. The reduction in yield per plant showed a trend from the rapid to the slow as plant height decreased. When yield per plant was reduced to 1.5 g or less because of the further frost stress, it became relatively stable as plant height continued to decrease, also, kernel number per ear did not change significantly any more at that time. 【Conclusion】 Under the simulated late frost stress, a significantly shortening trend occurred only in the imminent or elongated internodes and young ear, but not in the end of elongation. When the shortening internodes were consistent with the internodes that mainly contributed to the plant height, plant height significantly decreased. Using the shortening feature of the internodes, and the power function model between plant height and yield per plant, a new approach could be provided for the risk assessment of plant lodging in late growth and yield loss of winter wheat affected by late frost.
winter wheat; late frost stress; developmental progress; plant height; length of internode and ear; grain yield
10.3864/j.issn.0578-1752.2018.18.004
2018-03-23;
2018-06-19
国家自然科学基金面上项目(31771681)
武永峰,E-mail:wuyongfeng@caas.cn。通信作者游松财,E-mail:yousongcai@caas.cn
猜你喜欢
今日农业(2022年4期)2022-06-01
农业灾害研究(2022年1期)2022-05-07
少儿科学周刊·儿童版(2021年21期)2021-12-11
今日农业(2021年4期)2021-06-09
今日农业(2020年20期)2020-12-15
今日农业(2020年20期)2020-12-15
河北果树(2020年4期)2020-11-26
农村百事通(2019年17期)2019-10-08
现代农业科技(2017年1期)2017-03-06
江苏农业科学(2016年8期)2017-02-15
