
遮荫和氮养分对三裂叶蟛蜞菊生长、构件生物量分配和叶片PSⅡ功能的影响
2018-09-10 05:37:02陈文王桔红朱慧吴晓蓉颜玉娟汪翠丽
生态科学 2018年4期
陈文, 王桔红, 朱慧, 吴晓蓉, 颜玉娟, 汪翠丽
遮荫和氮养分对三裂叶蟛蜞菊生长、构件生物量分配和叶片PSⅡ功能的影响
陈文1, 王桔红2*, 朱慧2, 吴晓蓉2, 颜玉娟3, 汪翠丽2
1. 韩山师范学院地理与旅游管理学院, 潮州 521041 2. 韩山师范学院食品工程与生物科技学院, 潮州 521041 3. 中南林业科技大学风景园林学院, 长沙 410004
为探查外来入侵植物三裂叶蟛蜞菊()对遮荫和氮养分的响应及其入侵性, 采用盆栽实验研究了遮荫(轻度遮荫-相对光强为42%; 重度遮荫-相对光强为12%)和不同氮养分(0、0.1、0.2和0.4 g·kg–1)对三裂叶蟛蜞菊生长、生物量分配以及叶片PSⅡ功能的影响。结果表明: 1)经轻度遮荫的植物叶片数、分枝数、总生物量、生长速率显著大于重度遮荫; 同一遮荫条件下, 随氮养分的增加, 茎生物量比增加、叶生物量比下降。2)重度遮荫降低了三裂叶蟛蜞菊叶片实际光化学效率(ФPSⅡ)和表观光合电子传递速率(ETR), 对叶片PSⅡ反应中心有一定破坏作用; 对最大光化学量子产量(Fv/Fm)的影响不显著(除0.2 g·kg–1氮水平外)。3)氮养分对叶片PSⅡ活性的影响随遮荫程度而变化, 轻度遮荫下, 氮养分对叶片PSⅡ功能影响较小; 重度遮荫下, 随氮养分的增加, ФPSⅡ和ETR先上升后下降, Fv/Fm变化不显著。4)光强和氮养分的交互作用在三裂叶蟛蜞菊种群生长、竞争以及成功入侵过程中发挥着重要作用, 在光照较充足的生境中容易成功入侵。
三裂叶蟛蜞菊; 遮荫; 氮养分; 生长特性; PSII功能
1 前言
我国多样化的生境导致生态系统高度多样化, 外来植物入侵非常严重, 尤其是亚热带地区。国内已报道有515 种外来入侵植物, 有260 多种分布在我国暖湿的东南部亚热带地区[1]。根据 “十数定律”, 到达某一地区的外来种大约只有千分之一能够成为具有危害性的入侵种[2], 对入侵种入侵机制及防控的研究是入侵生态学的核心问题[3]。国内外众多学者从物种生物学特性以及生态系统可入侵性等多个角度尝试解释外来生物成功入侵的原因, 并提出了多样性阻抗、天敌逃避、生态位机遇、Novel Weapons 等假说[4-6]。近年来,有学者从表型可塑性方面对入侵植物开展了深入研究, 认为外来种成功入侵的原因是入侵物种比本地种和非入侵种的可塑性强, 而且大部分入侵物种的种群在入侵地发生了适应性进化[7], 使得物种获得了更宽的生态幅、广幅的环境耐受性,从而迅速占据多样化的生境[8-10]。因而, 表型可塑性可能在外来植物的成功入侵和随后的扩散中起到至关重要的作用[11], 较高的表型可塑性可能是外来种成功入侵和适应新生境的重要手段[7] [12-13]。三裂叶蟛蜞菊()是菊科(Asteraceae)多年生草本植物, 原产南美洲及中美洲, 20世纪70年代作为地被植物引入我国, 逃逸为野生, 能在路旁、田边、林下、林缘等多种生境中通过匍匐茎蔓延繁殖形成斑块状单优群落, 是一种适应性强、分布广的重度入侵植物, 严重威胁着当地生态系统及物种多样性[14]。国内外学者从化感作用[15-16]、入侵地根际土壤酶和土壤理化性质[17-18]以及对干旱[19]、高温[20]的表型可塑性等方面对三裂叶蟛蜞菊的入侵性开展了研究。然而, 生境光资源和氮养分发生波动时, 三裂叶蟛蜞菊生长、生物量分配[21-22]和光合生理将产生哪些响应, 目前所开展的研究较少。明确不同光资源和氮养分下三裂叶蟛蜞菊的生长特征、生物量分配格型和光合生理特性, 对揭示入侵植物对异质生境的适应及其入侵机制有重要作用。本研究设置2种遮荫条件和4种氮养分梯度, 探查三裂叶蟛蜞菊在不同遮荫和氮养分环境下其生长、构件生物量分配、叶绿素荧光特征及其对环境响应的可塑性反应, 以期为揭示外来植物的入侵机理和生物防治提供基础信息和有益参考。
2 材料与方法
2.1 研究地概况
研究地位于粤东地区 (22°37'N-24°91'N; 114°54'E—117°10'E), 该地域以丘陵平原地形为主, 属亚热带海洋性季风气候, 年均气温21-22 ℃, 日照1900-2400 h, ≥4 ℃的连续积温达7300-7890 ℃,年平均降雨量1300-2400 mm。极端最高气温为39.6 ℃, 极端最低气温为–0.5 ℃。
2.2 研究方法
2.2.1 研究材料
在广东潮州市郊区选取三裂叶蟛蜞菊重度入侵的斑块状单种优势群落, 剪取平均株高为12 cm 带芽茎段若干株进行30 d的水培生根。选取生长健壮、长势一致的再生小苗移栽到16.5 cm(上口径)×12.0 cm (下口径)×14.0 cm(高)塑料盆中, 每盆3株、进行栽种。栽培基质为由细沙和黄土充分均匀混合而成的混合土(2.6 kg·盆–1), 土壤理化性质为: 全碳含量47.6702 g·kg–1、全氮含量0.0980 g·kg–1、全磷含量0.0906 mg·kg–1、速效氮 40.13 mg·kg–1, 速效磷13.27 mg·kg–1。
2.2.2 盆栽实验
根据对三裂叶蟛蜞菊生境的调查, 实验设置两种遮荫条件: 轻度遮荫(Mild shade, MS), 相对光强为42%; 重度遮荫(Heavy shade, HS), 相对光强为12%。每种遮荫处理下设4种氮养分梯度, 分别为0(CK)、0.1、0.2、0.4 g·kg–1。所施氮肥为尿素, 采用水溶液形式进行添加。共施肥2 次, 分别在盆栽实验开始时以及30 d后各施入总添加量的50%(施肥后浇透水), 以满足植物的正常生长。每种处理4盆, 每盆栽种3棵植株, 每天浇足等量自来水, 保证植株不受水分限制, 适时防治病虫害; 盆栽实验90 d。
2.2.3 测定项目
(1)形态和生长: 栽种之前称量三裂叶蟛蜞菊植株生物量; 试验结束后测定其形态和生长参数, 包括株高、叶片数、分枝数、总生物量和相对生长速率(relative growth rate, RGR)。
相对生长速率(RGR) =(lnW2-lnW1)/(t2-t1)
式中, W1盆栽实验前三裂叶蟛蜞菊的总生物量(g), W2分别为实验结束时三裂叶蟛蜞菊的总生物量(g), 12个重复; (t2-t1)表示2次测定的时间间隔(90 d)。
(2)构件生物量分配: 试验结束时, 收集不同处理下植物的叶、茎、根, 在105 ℃烘箱杀青 30 min、85℃烘干至恒量后, 称量得到各构件干重,按Poorter等[23]、Yamashita等[24]计算各构件生物量分配指数:
叶生物量比(leaf mass fraction, LMF) = 叶生物量/植株总生物量
茎生物量比(stem mass fraction, SMF) = 茎生物量/植株总生物量
根生物量比( root mass fraction, RMF) = 根生物量/植株总生物量
总生物量(total biomass, TM)= 全株生物量/g
根冠比(root mass/crown mass, R/C) =根生物量/植株地上部分生物量
(3)叶绿素荧光指标: 叶绿素荧光参数采用PAM- 2500型便携式叶绿素荧光仪测定。涉及到的主要参数如下: Fo: 黑暗中最小荧光产量; Fm: 黑暗中最大荧光产量; Fv=Fm-Fo: 黑暗中最大可变荧光强度; Fv/Fm: 光合系统Ⅱ(PSⅡ)最大光化学量子产量; Fv/Fo: PSⅡ的潜在光化学活性; ETR:表观光合电子传递速率; ФPSⅡ:PSⅡ的实际光能转化效率[25]。
(4)表型可塑性指数: 按Valladares等[26]计算某一指标的可塑性指数:
对遮荫(氮)响应的可塑性指数=[某指标对遮荫(氮)响应的最大平均值-最小平均值]/最大平均值。
2.3 数据处理
采用 SPSS 17. 0 统计分析软件对数据进行统计分析和处理, 其中以一般线性模型(General Lineal Model)的单因素方差分析和双因素方差分析研究单个变量和2个自变量对植物生长指标的影响, 以最小显著差异法(LSD)比较不同处理数据间均值的差异性, 显著性水平设定为α=0.05。
3 结果与分析
3.1 遮荫和氮养分对三裂叶蟛蜞菊生长的影响
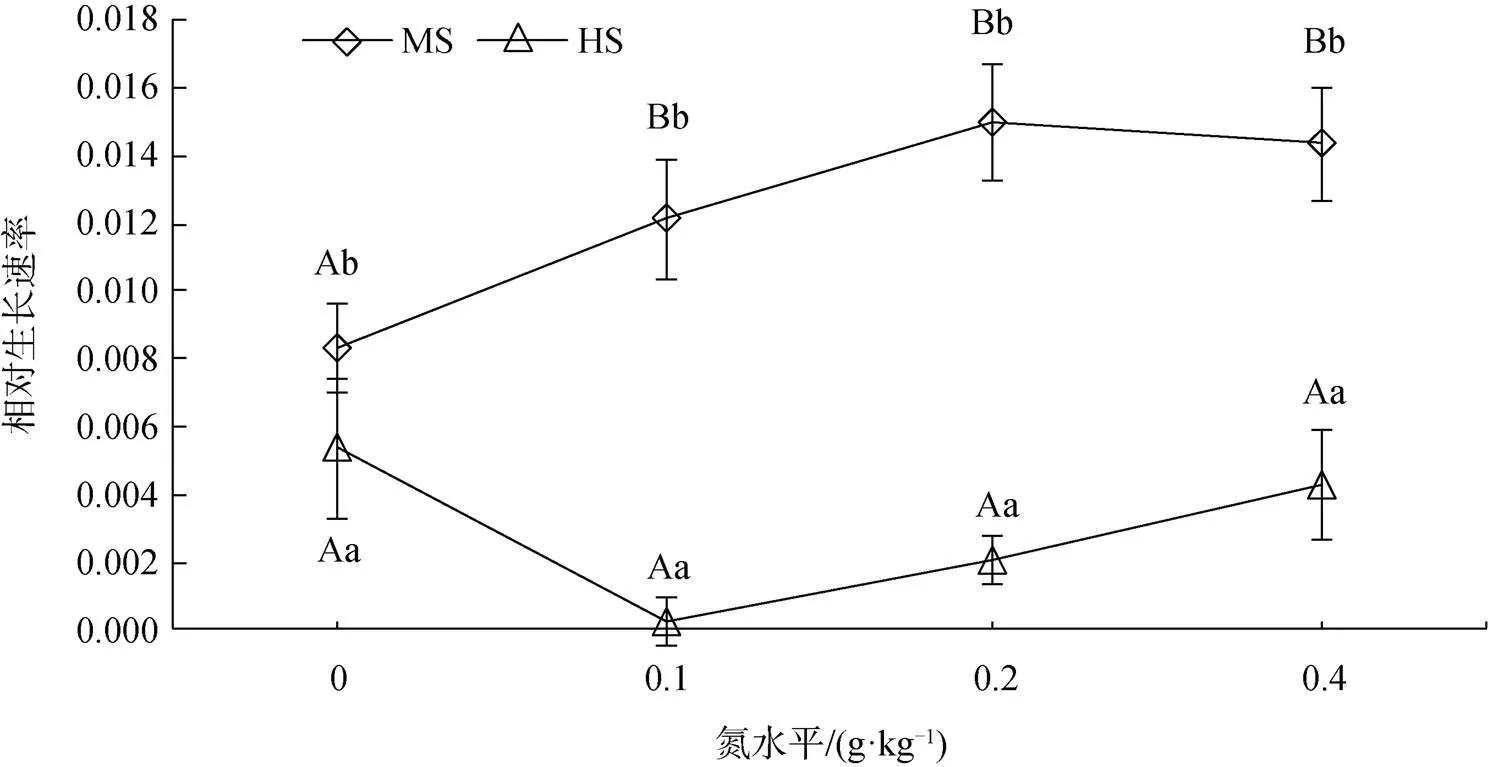
除株高外, 不同遮荫和氮养分对三裂叶蟛蜞菊生长特征均有不同程度的影响。在各种氮养分水平下, 轻度遮荫后植物叶片数、分枝数、总生物量显著大于重度遮荫下(<0.05)(除对照下叶片数)。轻度遮荫下, 叶片数和分枝数随氮养分的增加显著增大(<0.05), 总生物量几乎不受氮养分的影响(>0.05); 重度遮荫下, 叶片数、分枝数、总生物量几乎不受氮养分的影响(>0.05)(表1)。在各种氮养分下(除对照外), 轻度遮荫下植物生长速率显著大于重度遮荫下(<0.05)。在轻度遮荫下, 植物的生长速率随氮养分的增加而增大, 但不显著(>0.05); 重度遮荫下植物生长速率几乎不受氮养分的影响(>0.05)(图1)。

表1 不同遮荫和氮养分下三裂叶蟛蜞菊生长特性 (平均值±标准差)
MS: 轻度遮荫(相对光照强度42%); HS: 重度遮荫(相对光照强度12%)。
注: 大写字母表示不同遮荫水平下差异性, 小写字母表示不同氮水平下差异性;不同字母为差异性显著(<0.05)。下同。
双因素方差分析显示, 遮荫是影响植物叶片数和分枝数的主要因素, 其贡献率分别为83%和87%; 遮荫和氮养分的交互作用对叶片数和分枝数也有显著影响(<0.05), 其贡献率分别为14%和9.4%。遮荫是影响总生物量和生长速率的主要因素, 贡献率分别为97%和86%,氮养分对总生物量和生长速率的影响不显著(>0.05)(表2)。
3.2 遮荫和氮养分对三裂叶蟛蜞菊构件生物量分配的影响
遮荫和氮养分水平对三裂叶蟛蜞菊构件生物量分配有不同的影响。轻度遮荫下叶生物量比随氮养分的增加呈现下降趋势(<0.01), 重度遮荫下叶生物量比为先下降后上升的趋势, 但不显著(>0.05); 2种遮荫水平下茎生物量比随氮养分增加呈现先上升后下降趋势(<0.05)。不同遮荫处理对叶生物量比和茎生物量比的影响较小(>0.05) (图2A)。在0和0.1 g·kg–1氮养分水平下, 不同遮荫对根生物量比和根冠比的影响较小或没有影响(>0.05), 在0.2 g·kg–1和0.4 g·kg–1氮养分水平下, 轻度遮荫下的根生物量比和根冠比显著大于重度遮荫下(<0.01), 遮荫显著降低了根生物量比和根冠比。不同氮养分对根生物量比和根冠比的影响较小或没有影响(>0.05)(图2B)。
双因素方差分析显示, 氮养分是影响叶生物量比和茎生物量比的主要因素, 其贡献率分别为89%和88%; 遮荫是影响根生物量比和根冠比的主要因素, 其贡献率分别为80%和81%(表2)。
3.3 遮荫和氮养分对三裂叶蟛蜞菊叶片PSII功能的影响
遮荫对叶片PSⅡ功能有显著影响。在各种氮养分水平下, 轻度遮荫的叶片初始荧光F0显著小于重度遮荫(<0.05); 叶片实际光化学效率(ФPSⅡ)、表观光合电子传递速率(ETR)、最大光化学量子产量Fv/Fm(0.2 g·kg–1氮养分下)显著大于重度遮荫下(<0.05)。氮养分对叶片PSⅡ功能的影响随遮荫程度而变化。轻度遮荫下, 不同氮养分对三裂叶蟛蜞菊叶片PSII功能影响较小(>0.05); 重度遮荫下, 氮养分对叶片F0(=5.984,<0.01)、ФPSⅡ(=6.672,<0.01)、ETR(=7.766,<0.01)有显著影响, 随氮养分的增加, F0、ФPSⅡ和ETR先上升后下降, 在氮养分水平为0.2 g·kg–1时ФPSⅡ和ETR达到最高; 2种遮荫条件下, 不同氮养分对Fv/Fm无显著影响(>0.05)(表3)。
3.4 三裂叶蟛蜞菊生长性状对遮荫和氮养分响应的可塑性指数
由表4可知, 植物叶片数、分枝数、总生物量、叶生物量比和茎生物量比对光强响应的可塑性指数大于对氮养分的响应, 植物株高、根冠比对氮养分响应的可塑性指数大于对光强的响应。
注: 大写字母表示不同遮荫水平下差异性, 小写字母表示不同氮水平下差异性;不同字母为差异性显著(P<0.05)。
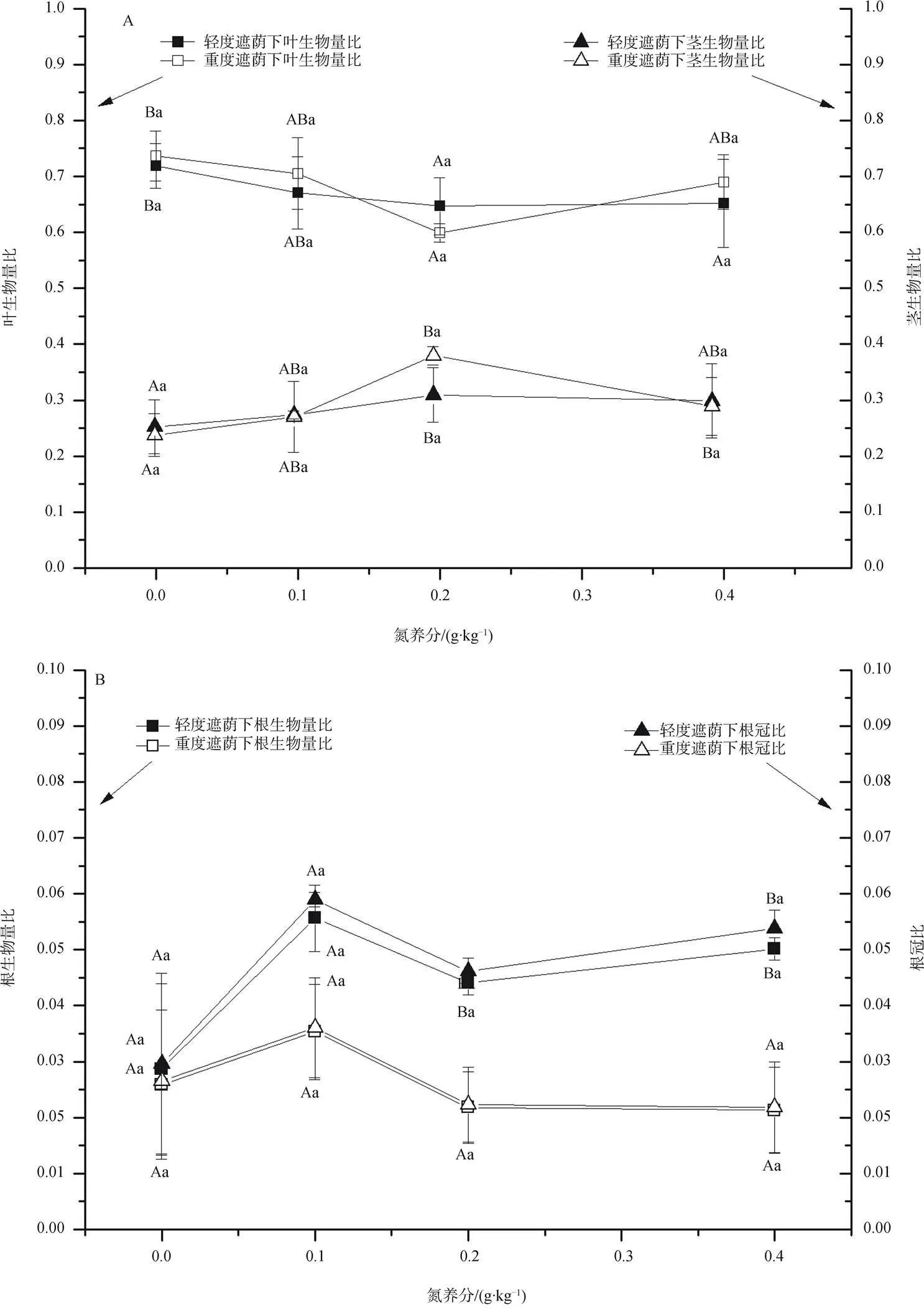
(A: 叶生物量比和茎生物量比; B: 根生物量比和根冠比)
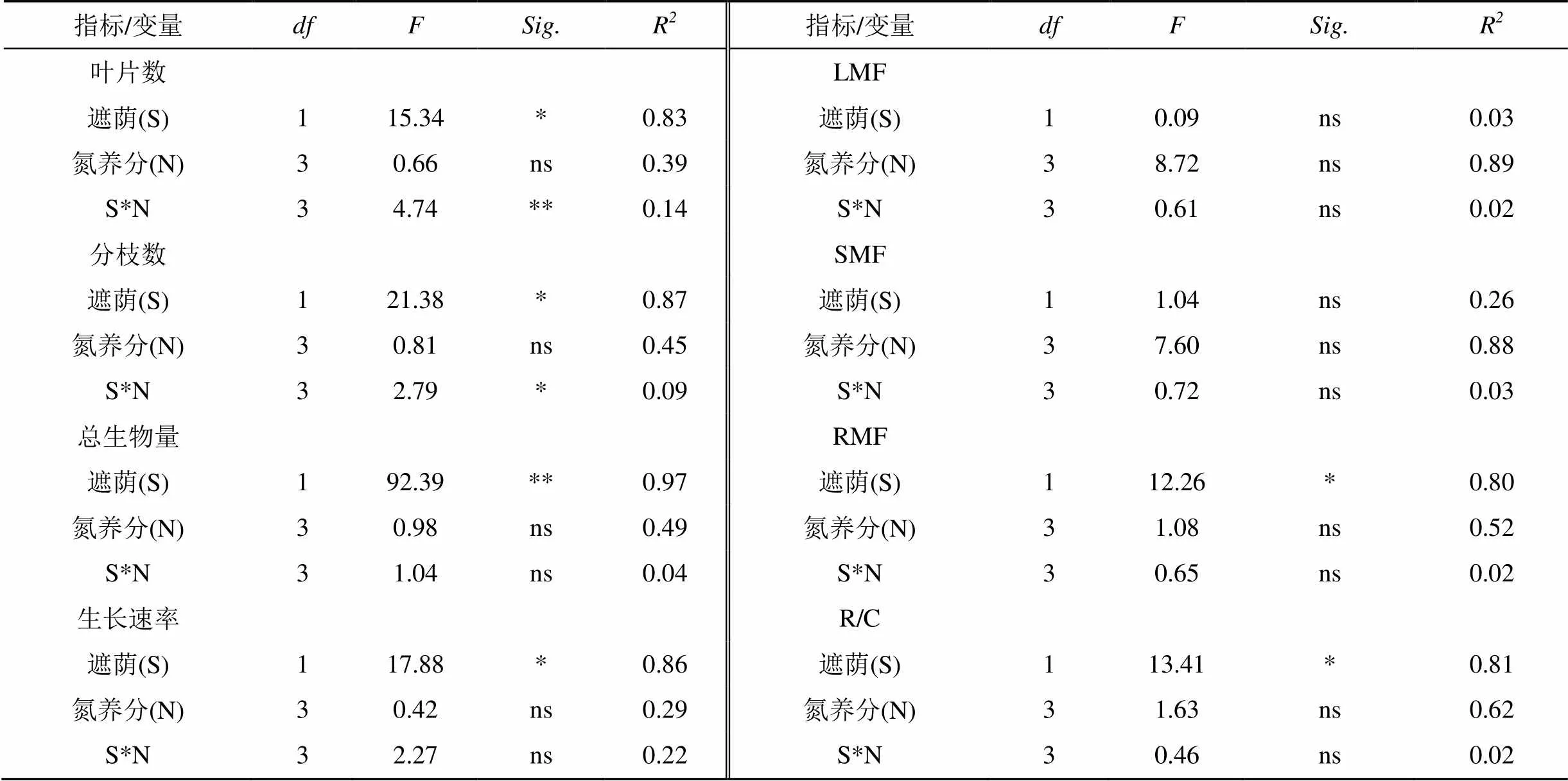
表2 遮荫和氮养分对三裂叶蟛蜞菊生长和构件生物量分配影响的双因素方差分析
**<0.01, *<0.05, ns为差异不显著>0.05

表3 不同遮荫和氮养分下三裂叶蟛蜞菊的叶绿素荧光参数 (平均值±标准差)
注: 大写字母表示不同遮荫水平下差异性, 小写字母表示不同氮水平下差异性;不同字母为差异性显著(<0.05)。

表4 三裂叶蟛蜞菊生长对遮荫和氮养分响应的可塑性指数
*表示同一生长指标的最大可塑性指数。
4 结论与讨论
植物的表型可塑性是同一基因型对不同环境条件表现出的不同表型特性[26]。在高度异质性的野外生境中, 可塑性强的入侵种能够通过自身调节机制及生长策略对不利的环境条件产生可塑性反应, 增强对新环境资源的适应性。本研究显示, 三裂叶蟛蜞菊叶片数、分枝数、生长速率在轻度遮荫下大于重度遮荫下; 在土壤氮养分达到0.2 g·kg–1和0.4 g·kg–1, 根生物量比和根冠比在轻度遮荫下显著大于重度遮荫下。因此, 三裂叶蟛蜞菊在低荫蔽生境通过产生较多叶片、分枝数、较快的生长速率以及较大的地下生物量以增大竞争和扩张力。
叶绿素荧光是光合作用的探针, 通过荧光参数可以认识光合机构内部一系列重要的调节过程。初始荧光FO是判断PSⅡ反应中心运转情况的重要指标, FO上升表明PSⅡ反应中心受到破坏或可逆失活; Fv/Fm表示最大光化学量子产量, 反映PSⅡ原初光能转化效率及反应中心内禀光能转换效率, 当植物处于非逆境条件下时, 该参数一般为0.75-0.85, 胁迫条件下该参数明显下降; ФPSⅡ是PSⅡ的实际光能转化效率, 与 PSⅡ的活性呈正相关; ETR反映光照条件下的表观电子传递效率, 与植物净光合速率呈显著正相关[23]。各项叶绿素荧光指标显示, 遮荫对三裂叶蟛蜞菊叶片PSII功能有显著影响。随遮荫程度加重, FO值上升, PSII反应中心受到一定破坏; ΦPSII和ETR值在轻度遮荫下大于重度遮荫, 说明充足的光照对叶片PSII反应中心的光能捕获和传递起积极作用。Fv/Fm值在轻度遮荫和重度遮荫下差异不显著(除氮养分0.2 g·kg–1以外), 显示三裂叶蟛蜞菊对重度遮荫仍有一定耐受性。因此, 在较充足的光强下, 三裂叶蟛蜞菊PSⅡ功能有所增强, 利于生长和扩张; 弱光或重度遮荫下, 生长和光合明显受抑。该结果同许凯等(2005)对入侵植物喜旱莲子草()[27]、王俊峰和冯玉龙(2004)对紫茎泽兰()[28], 王蕊等(2012)对三裂叶豚草()[29]以及孙晓方等(2008)对入侵植物加拿大一枝黄花()的研究结果[30]一致, 说明光强是影响入侵植物生长、竞争的扩张的重要因子之一。
研究还发现, 在轻度遮荫(相对光强为42%)条件下, 三裂叶蟛蜞菊各生长指标(叶片、分枝、生长速率)和茎生物量分配随氮养分的增加而增大, 说明三裂叶蟛蜞菊在氮养分增加的环境中通过增大营养生长来增强竞争力。然而, 也有研究发现, 依赖种子繁殖的植物如飞机草()其茎生物量随养分增加而减小[31-32], 其机制有待进一步研究。双因素方差分析进一步说明光照强度和氮养分的交互作用是三裂叶蟛蜞菊种群生长、竞争以及成功入侵和扩散的重要因子, 其中光照是主导因子, 光照比养分更能影响三裂叶蟛蜞菊的入侵生长。
综上所述, 三裂叶蟛蜞菊可通过生长、构件生物量分配以及光合生理等特征适应生长环境的光强变化和土壤氮养分变化。光照强度是影响三裂叶蟛蜞菊生长、光合的主要因素, 三裂叶蟛蜞菊在光照较充足的生境中容易成功入侵。
[1] 闫小玲, 刘全儒, 寿海洋, 等. 中国外来入侵植物的等级划分与地理分布格局分析[J]. 生物多样性, 2014, 22(5):667–676.
[2] WILLIAMSON M. Biological Invasions[M]. London: Chapman and Hall, 1996: 20
[3] ALPERT P, BONE E, HOZAPFEL C. Invasiveness, invisibility and the role of environmental stress in the spread of nonnative plants[J]. Perspectives in Plant Ecology Evolution and Systematics, 2000, 3: 52–66.
[4] CALLAWAY R M, ASCHEHOUG E T. Invasive plants versus their new and old neighbors: a mechanism for exotic invasion[J]. Science, 2000, 290: 521-523.
[5] WOLFE L M. Why alien invaders succeed: support for the escape-from-enemy hypothesis[J]. American Naturalist, 2002, 160: 705–711.
[6] 吴锦容, 彭少麟. 化感—外来入侵植物的“Novel Weapons”[J]. 生态学报, 2005, 25(11): 3093–3097.
[7] 耿宇鹏, 张文驹, 李博, 等. 表型可塑性与外来植物的入侵能力[J]. 生物多样性, 2004, 12(4): 447–455.
[8] SULTAN S E, BAZZAZ F A. Phenotypic plasticity in. Ⅲ. The evolution of ecological breadth for nutrient environment[J]. Evolution, 1993, 47: 1050–1071.
[9] 潘晓云, 耿宇鹏, 张文驹, 等. 喜旱莲子草沿河岸带不同生境的盖度变化及形态可塑性[J]. 植物生态学报, 2006, 30(5): 835–843.
[10] GENG Yupeng, PAN Xiaoyun, Xu Chengyuan, et al. Phenotypic plasticity rather than locally adapted ecotypes allows the invasive alligator weed to colonize a wide range of habitats[J]. Biological Invasions, 2007, 9: 245–256.
[11] 宫伟娜, 万方浩, 谢丙炎, 等. 表型可塑性与外来入侵植物的适应性[J]. 植物保护, 2009, 35(4): 1–7.
[12] REN Mingxun, ZHANG Quanguo. The relative generality of plant invasion mechanisms and predicting future invasive plants[J]. Weed Research, 2009, 49: 449–460.
[13] 蒲高忠, 唐赛春, 潘玉梅, 等. 入侵植物银胶菊在不同生境下表型可塑性和构件生物量[J]. 广西植物, 2010, 30(5): 641–646.
[14] 吴彦琼, 胡玉佳, 廖富林. 从引进到潜在入侵的植物—南美蟛蜞菊[J]. 广西植物, 2005, 25(5): 413–418.
[15] 江贵波, 曾任森. 入侵物种三裂叶蟛蜞菊挥发物的化感作用研究[J]. 生态环境, 2007, 16(3): 950–953.
[16] 曾任森, 林象联, 谭惠芬, 等. 蟛蜞菊根分泌物的异种克生作用及初步分离[J]. 生态学杂志, 1994, 13(1): 51–56.
[17] 柯展鸿, 邱佩霞, 胡东雄, 等. 三裂叶蟛蜞菊入侵对土壤酶性和理化性质的影响[J]. 生态环境学报, 2013, 22(3): 432–436.
[18] 许华, 邓晓勇, 程碧军. 三裂叶蟛蜞菊和薇甘菊复合群落对土壤化学和生物学特征的影响[J]. 湖北农业科学, 2015, 54(16):3932–3935.
[19] 宋莉英, 孙兰兰, 舒展, 等. 干旱和复水对入侵植物三裂叶蟛蜞菊叶片叶绿素荧光特性的影响[J]. 生态学报, 2009, 29(7):3715–3721.
[20] 宋莉英, 孙兰兰, 张强, 等. 高温对入侵种三裂叶蟛蜞菊叶片 PSII 功能和光能分配的影响[J]. 植物生理学通讯, 2009, 45(5): 464–468.
[21] 杨东娟, 赵锐明, 回嵘, 等. 不同生长方式对2种入侵植物形态特征和生物量分配的影响[J]. 西北林学院学报, 2014, 29(4): 69–73.
[22] 袁伟影, 冯进, 张晓雅, 等. 入侵植物南美蟛蜞菊和本土蟛蜞菊生长对土壤养分的响应[J]. 生态学杂志, 2017, 36(4): 1–9.
[23] POORTER L. Growth responses of 15 rainforest tree species to a light gradient: The relative importance of morphological and physiological traits[J]. Functional Ecology, 1999, 13: 396–410.
[24] YAMASHITA N, KOIKE N, ISHIDA A. Leaf ontogenetic dependence of light acclimation in invasive and native subtropical trees of different successional status[J]. Plant, Cell and Environment, 2002, 25: 1341–1356.
[25] 尤鑫, 龚吉蕊. 叶绿素荧光动力学参数的意义及实例辨析[J]. 西部林业科学, 2012, 41(5): 90–94.
[26] VALLADARES F, WRIGHT SJ, LASSO E, et al. Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest[J]. Ecology, 2000, 81: 1925–1936.
[27] 王蕊, 孙备, 李建东, 等. 不同光强对入侵种三裂叶豚草表型可塑性的影响[J]. 应用生态学报, 2012, 23(7): 1797– 1802.
[28] 许凯, 叶万辉, 李国民, 等. 入侵种喜旱莲子草对光照强度的表型可塑性反应[J]. 武汉植物学研究, 2005, 23(6): 560–563.
[29] 王俊峰,冯玉龙. 光强对两种入侵植物生物量分配、叶片形态和相对生长速率的影响[J]. 植物生态学报, 2004, 28(6): 781–786.
[30] 孙晓方, 何家庆,黄训端, 等. 不同光强对加拿大一枝黄花生长和叶绿素荧光的影响[J]. 西北植物学报, 2008, 28(4): 0752–0758.
[31] 全国明, 毛丹鹃, 章家恩, 等. 不同养分水平对飞机草生长与生物量分配的影响[J]. 生态科学, 2015, 34(2): 27– 33.
[32] 全国明, 毛丹鹃, 章家恩, 等. 飞机草的繁殖能力与种子的萌发特性[J]. 生态环境学报, 2011, 20(1): 72–78.
Effects of shade and nutrient level on plant growth, module biomass allocation and PSII functions of
CHEN Wen1, WANG Juhong2*, ZHU Hui2, WU Xiaorong2, YAN Yujuan3, WANG Cuili2
1. College of Geography and Tourism Management, Hanshan Normal University, Chaozhou 521041, Guangdong, China 2. College of Food Technology and Life Science, Hanshan Normal University, Chaozhou 521041, Guangdong, China 3. Central South University of Forestry and Technology, Shangsha 410000, Hunan, China
In order to research the response of alien speciesto shading condition and nitrogen nutrient and further to reveal the invasion, the effects of shade and nutrient levels on plant growth, biomass allocation patterns, and PSII functions of invasivewere studied by using a series of greenhouse pot experiment. Two shading conditions and four nitrogen level were set up with relative light intensity of 42% (mild shade) and of 12% (heave shade), and with nitrogen level of 0, 0.1, 0.2, and 0.4 g·kg–1. The results showed that shading condition and nutrient levels have important influence on plant growth, biomass allocation patterns in varying degrees, except for plant height. Under the various nitrogen levels, the leaf number, branching number, total biomasses and relative growth rate in mild shade were significantly greater than those in heave shade. Under the same shading condition, stem biomass increased, while leaf biomass decreased with increasing of nutrient concentration. In addition, under heave shade conditions, the actual photochemical efficiency of PSⅡ(ФPSⅡ) and photosynthetic electron transport rate (ETR) decreased, minimum fluorescence (F0) increased, the value of the maximal quantum yield of PSⅡ(Fv/Fm) had no or less change comparison with the mold shade, implying low light level might have some damage for PSII functions. The effects of nitrogen level on the leaf PSII functions varied with the degree of shade. Under mold shading condition, the nitrogen level had no or less influence on leaf PSII functions, but under heave shade the ФPSⅡand ETR first increased, then decreased with increasing of nitrogen level; the value of Fv/Fm had no or less change. The interaction effect of light intensity and nitrogen nutrient play an important role on growth, competition and invading of, in which light intensity is the dominant factor. It is easier to invade forin habitats with sufficient light than in heavy shade habitat.
; shading condition; nitrogen nutrients; growth traits; photosystem II functions
10.14108/j.cnki.1008-8873.2018.04.020
Q142.9
A
1008-8873(2018)04-160-08
2017-08-31;
2017-12-19
国家自然科学基金项目(31770584); 广东省科技发展专项资金(2016A030303063)
陈文(1963—), 男, 甘肃张掖人, 副研究员, 主要从事环境与生态学研究, Email:cyw1018@sina.com
王桔红(1963-), 女, 博士, 教授, 主要从事入侵生物学研究, E-mail : wjuh1918@163.com
陈文, 王桔红, 朱慧, 等. 遮荫和氮养分对三裂叶蟛蜞菊生长、构件生物量分配和叶片PSⅡ功能的影响[J]. 生态科学, 2018, 37(4): 160-167.
CHEN Wen, WANG Juhong, ZHU Hui, et al. Effects of shade and nutrient level on plant growth, module biomass allocation and PSII functions of[J]. Ecological Science, 2018, 37(4): 160-167.
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28 07:43:34
昆明医科大学学报(2021年8期)2021-08-13 08:59:20
昆明医科大学学报(2021年6期)2021-07-31 07:40:08
落叶果树(2021年6期)2021-02-12 01:28:54
今日农业(2020年19期)2020-12-14 14:16:52
文苑(2020年12期)2020-04-13 00:54:10
海峡姐妹(2019年1期)2019-03-23 02:42:46
中国科技信息(2015年2期)2015-11-16 08:18:32
西南医科大学学报(2015年1期)2015-08-22 13:01:48
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
