
Morphological phylogeny among 15 accessions of bamboos growing in Kokrajhar district of the Bodoland Territorial Area Districts,Assam
2018-09-07 03:06AnusritiBasumatarySushilKumarMiddhaTalambeduUshaSamikBhattacharyaBirendraKumarBrahmaArvindKumarGoyal
Journal of Forestry Research 2018年5期
Anusriti Basumatary•Sushil Kumar Middha•Talambedu Usha•Samik Bhattacharya•Birendra Kumar Brahma•Arvind Kumar Goyal
Abstract Bamboo is nature’s wonderful gift historically associated with the rural population of the Bodoland Territorial Area Districts.However,indiscriminate use of the resource and the ambiguity in the identification of bamboos in the absence of flowering characters pose a potential risk to the bamboo germplasm.Therefore,we considered revising the taxonomic positions and phylogenetic relationships among 15 tropical bamboo species using 35 key morphological descriptors(MD)to assess the reliability of vegetative characters in identifying species.The phylogenetic tree constructed from the similarity matrix,derived from an unweighted pair group method of analysis(UPGMA)of the 35 MDs,marginally deviated from the conventional taxonomic positions of the bamboo species.While the two varieties,one forma of Bambusa vulgaris(var.Vittata,var.Vulgaris,fo.Waminii)and two species of the genus Dendrocalamus(D.giganteus and D.hamiltonii)clustered together in a single clade,and D.strictus was distantly placed from their congeners.This discrepancy with the conventional classification might have ensued since only vegetative characters such as culm anatomy,culm sheath,leaf,and leaf sheath were considered in the present study.While the phylogenetic relationship among the 15 tropical bamboos(computed from exhaustive MDs)grossly corroborate the conventional taxonomic grouping,a molecular-marker-based multidisciplinary approach should resolve the remaining conflicts in bamboo identification.
Keywords Bamboo·Bodoland·Culm ·Culm-sheath·Leaf-sheath·Phylogenetic relationship conflict
Introduction
The northeastern states of India are among one of the richest repositories of bamboo.In this region,bamboo also has high species diversity(Barooah and Borthakur 2003;Hore 1998)and age-old connections with the rural people(Mukherjee et al.2010).It continues to play a key role in the lifestyle and economy of Asian countries due to its many uses,and a large part of society still heavily relies on it(Brahma et al.2014;Das et al.2008).Also,bamboo has always been a cultural feature of Southeast Asia.In India,one of the largest reserves of bamboos in the world,are 136 exotic and indigenous species(Goyal and Sen 2015)of bamboos,and over 50%of the genotypes are found in the northeastern states[Assam,Arunachal Pradesh,Sikkim,Nagaland,Meghalaya,Mizoram,and Manipur including West Bengal(North Bengal,Himalaya)](Goyal and Brahma 2014;Biswas 1988;Bahadur and Jain 1983).Assam has a geographical area of about 78,438 km2,1813 km2of which shelters bamboo(Basumatary et al.2015;Sharma et al.2010).
In world trade,bamboo,is the most unique,hard and dominant flora that has been utilized traditionally for various purposes.Also,due to its high desirability,sustainability and growth rate,bamboo has regained attention and recognition worldwide(Vengala and Lindt 2013;Ghosh et al.2012).Around the globe,bamboo has a notable biodiversity of species,and its wide genetic diversity necessitates appropriate identification of the plant materials that will serve as the basis for successful conservation,plant improvement,and utilization(Tamang et al.2013).
Overexploitation,loss of bamboo species and its genetic erosion have made it necessary for the taxonomic classification,conservation and collection of the germplasm and characterization of bamboos with the help of its morphological characters(Yeasmin et al.2015;Loh et al.2000).Thus,to maintain the conservation of biodiversity and germplasms,studying and assessing the relationship among different species of bamboos is indispensable(Goyal et al.2012).
A group of tall arborescent grasses(Poaceae),the bamboos comprise about 1575 species among 111 genera in the subfamily Bambusoideae(Ghosh et al.2011;Goyal and Sen 2016).Bambusoideae,perhaps the most primordial subfamily of the grasses,is usually divided into the tropical woody tribe(Bambuseae),temperate woody tribe(Arundinarieae)and herbaceous tribe(Olyreae)(Sungkaew et al.2009).The taxonomy of woody bamboos(Bambuseae)is still in a state of flux due to their erratic flowering,different interpretations of morphological features and the terminology used for different parts(Goyal et al.2014).In this study,35 key morphological descriptors are used for evaluating the phylogenetic relationships among the 15 accessions of tropical woody bamboos.
Materials and methods
Plant material
Healthy culms,culm sheaths,leaves and leaf sheaths of 15 bamboo species were collected in situ from the Kokrajhar district of BTAD,Assam,India for morphological and phylogenetic analyses:Bambusa assamica(B1),Bambusa balcooa(B2),Bambusa bambos(B3),Bambusa garuchokua(B4),Bambusa multiplex(B5),Bambusa pallida(B6),Bambusa tulda(B7),Bambusa vulgarisvar.Vittata(B8),Bambusa vulgarisvar.Vulgaris(B9),Bambusa vulgarisfo.Waminii(B10),Dendrocalamus giganteus(B11),Dendrocalamus hamiltonii(B12),Dendrocalamus strictus(B13),Melocanna baccifera(B14),Schizostachyumsp.(B15).The species were sampled over a region of 3165 km2in the state of Assam,known as a bamboo biodiversity hotspot(Fig.1,Table 1).
Morphological descriptor scoring
Each of the 15 species was considered as a separate independent operational taxonomical unit(OTU).For each ofthe 15 OTUs,at least four replicate samples were studied from different populations in the field for 35 key morphological descriptors(MD,14 culm,14 culm-sheath and 7 leaf and leaf-sheath).Details of observed MDs and the specific incremental values assigned for qualitative variations within each of them are described in Appendix S1.For each of the quantitative MDs,mean values obtained from all of the independent replications were used as the representative data for the OTUs.
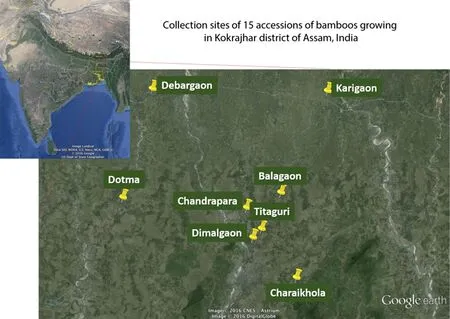
Fig.1 Map of collection sites in Assam with inset showing location in India
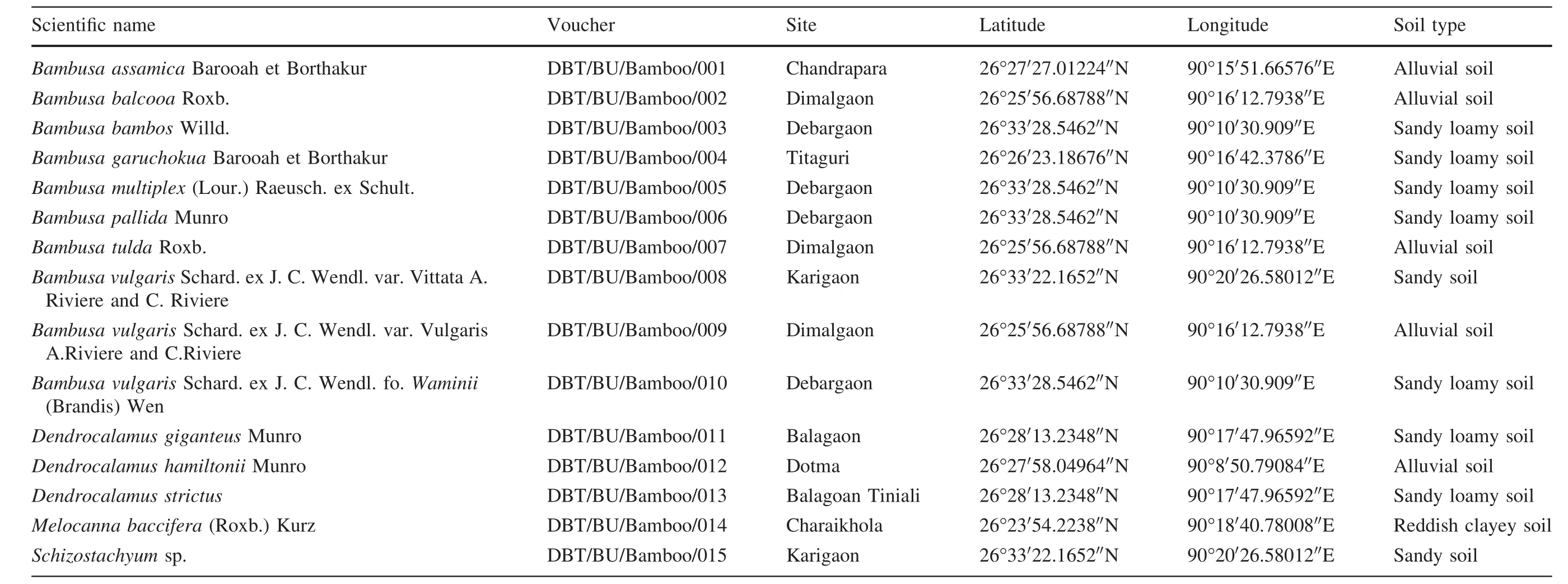
Table 1 Accessions with collection site in Kokrajhar district and soil type that were examined morphologically
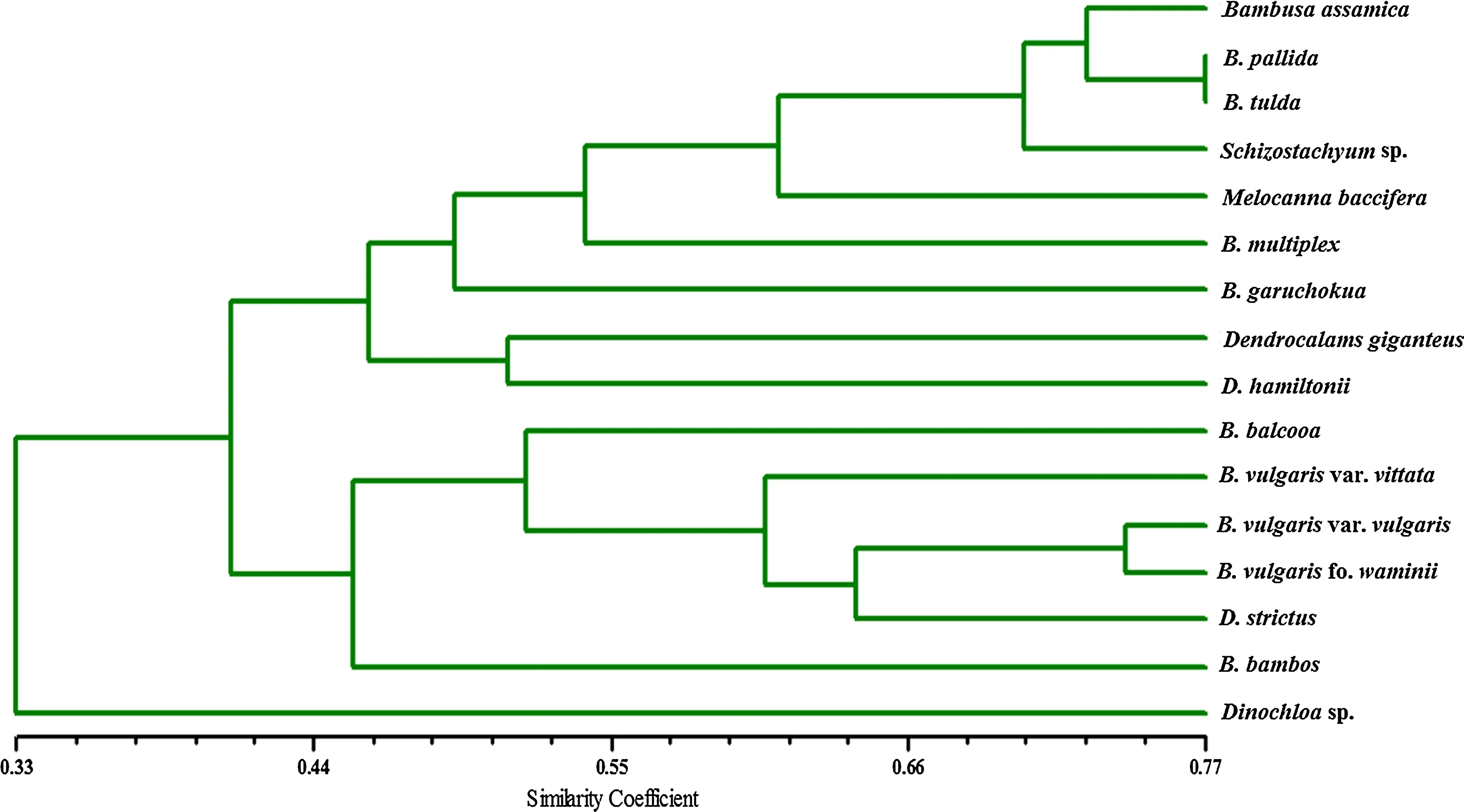
Fig.2 Dendrogram generated from UPGMA cluster analysis based on 35 key morphological descriptors for 15 bamboo accessions
We also includedDinochloasp.in our analysis to form an outgroup because of its classical taxonomic diversity from our 15 OTUs.However,due to the unavailability of a living sample ofDinochloaor other potential outgroup species in our sampling location,we extracted morphological data from the literature(Clayton et al.2002;Barooah and Borthakur 2003).
Phylogenetic analysis of data
Standardized quantitative and qualitative interval data among the OTUs were used to compute the similarity matrix with the UPGMA method(Sneath and Sokal 1973)in NTSYS-pc ver.2.2 for constructing the dendrogram(Das et al.2007;Rohlf 2000).Based on the coefficient of similarity,the same software was used to further calculate the pairwise genetic distances(GD)among the OTUs(Nei and Li 1979).Moreover,the relative discriminatory value of a descriptor is estimated from the polymorphism information content(PIC)of each MDs which is calculated by the following equation:

where,PICiis the polymorphic information content of theith descriptor calculated as the square of the frequency(Pij)of thejth value for that descriptor and summed over all binning array(n)values(Das et al.2007;Ni et al.2002).In simple words,when a descriptor(i)has several possible binning arrays(e.g.,0,1,2 and so on),the frequency of the first binning array(i.e.,0)isPi1=AAA,and that of second binning array isPi2=BBB and so on.The PIC for the descriptor is then calculated as the summation of the squares of all the frequencies(i.e.,AAA2+BBB2+···)(Das et al.2007;Ni et al.2002).
We also transformed the replicate morphological data to extract the most effective morphological character(s)in discriminating the OTUs by principal coordinate analysis(PCA)with the web-based analytical software MetaboA-nalyst 3.0(Xia and Wishart 2016).
Results and discussion
Bamboo flowering still remains a great mystery because the timing of flowering of woody bamboos is uncertain(Goyal et al.2014;Mukherjee et al.2010;Ramanayake et al.2007).This unique feature of the unusually long
sexual cycle of bamboos limits the availability of their floral characters.Thus,in the absence of flowers,attempts to develop conventional keys have very often caused confusion in the determination of accurate evolutionary interrelationship and in taxonomic groupings(Ghosh et al.2012).Consequently,various intricate features of the culm and culm heath,as well as other vegetative features,have been considered as potential descriptors by bamboo taxonomists.Culm,culm sheath,leaf and leaf sheath characters are treated as major taxonomic keys for identifying bamboos in the absence of fruit and flower characters(Appendix S1).
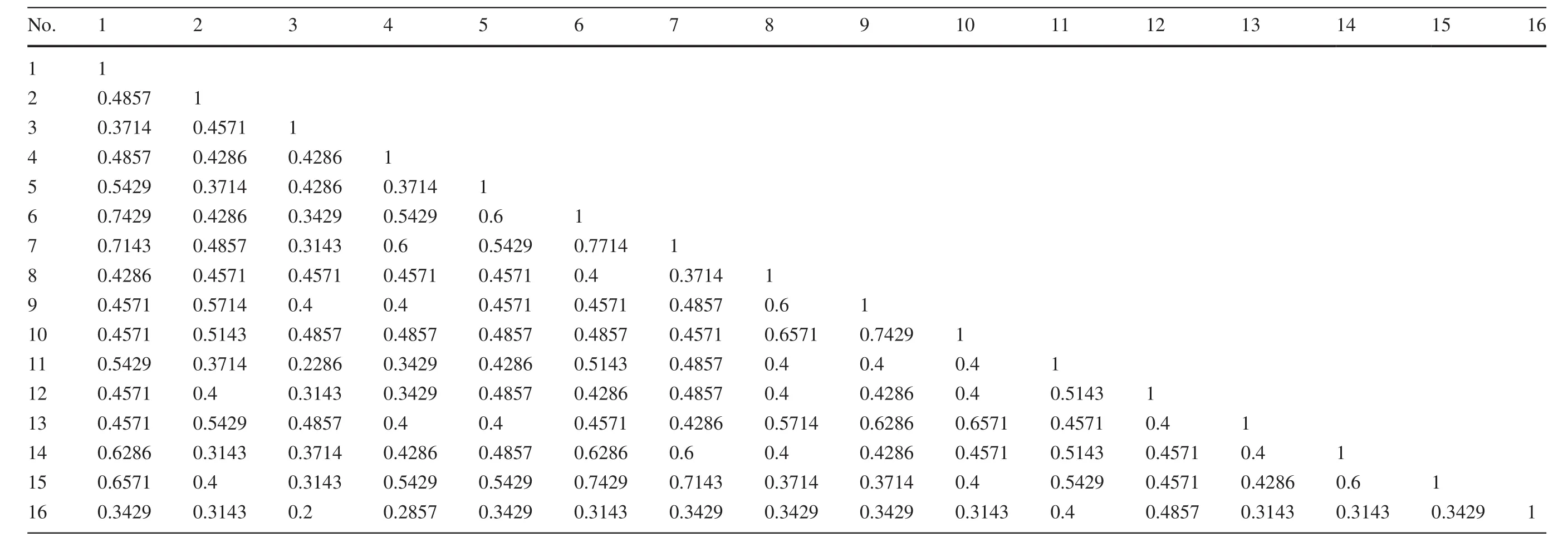
Table 2 Simple matching (SM) m/n distance coefficient matrix for 15 bamboo species (OTUs) generated through morphometric data of 35 key morphological characters
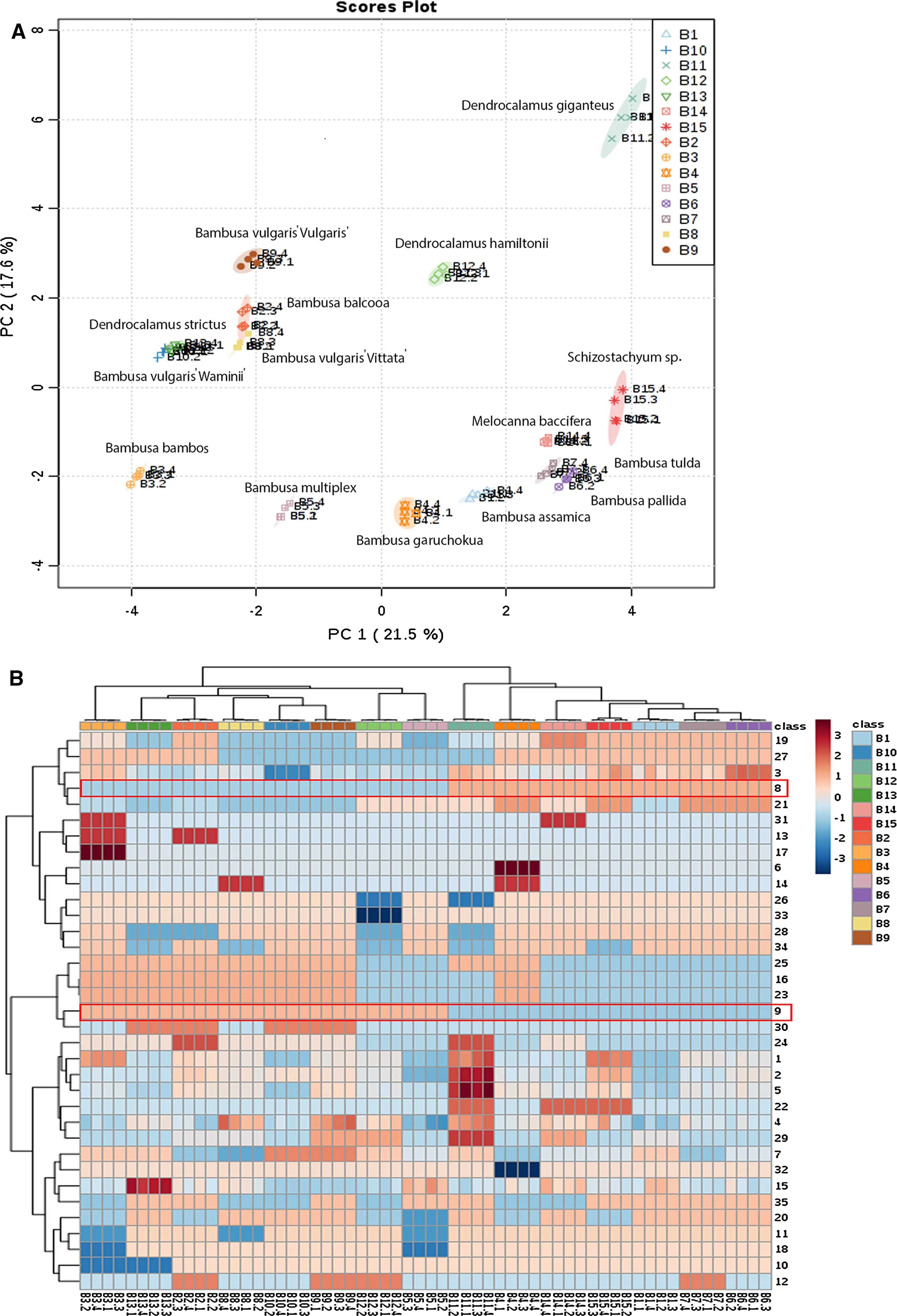
Fig.3 Principle component analysis(PCA)and two-way heat map of the characters and operational taxonomic units(OTUs)identify the most discriminatory morphological descriptors(MDs).Four replicate data of 35 MDs for 15 OTUs were analyzed to generate PCA score plot depending on the eigen values of the two most discriminatory components.Most discriminatory morphological descriptors(#8 and#9)were highlighted with a red rectangles in the two-way heat map showing the effect of each of the 35 MDs in discriminating the 15 OTUs
The phylogenetic tree constructed from the 35 morphological descriptors revealed two distinct clades(apart from the outgroup taxonDinochloasp.)that included 10 members of the genusBambusa,except forBambusa pallidaandBambusa tulda,that were further clustered together(Fig.2).Bambusa tuldaandB.pallidawere very similar morphologically,except for their culm diameter and color(Appendix S1),and therefore expected to group together with the highest coefficient of similarity(77.14%,Table 2).
One of the three accessions ofDendrocalamuswas distantly placed,showing considerable divergence among the three in contrast to traditional taxonomy.This divergence among species of the same genus might be due to their growth habit or morphology.D.giganteuswas clustered withD.hamiltonii,sharing a node at 51.43%,whereasD.strictuswas grouped withB.vulgarisvar.Vittata,B.vulgarisvar.Vulgaris,andB.vulgarisfo.Waminii.Two varieties and one form a of the same species,Bambusa vulgariswere close to each other.B.vulgarisvar.Vittata andB.vulgarisvar.Vulgaris were 60%similar,being varieties of the same species.Similarly,B.vulgarisvar.Vittata andB.vulgarisvar.vulgaris with 65.71 and 74.29%similarity were found closely linked withB.vulgarisfo.Waminii.This close proximity has already been documented by different authors(Nayak et al.2003;Das et al.2007;Mukherjee et al.2010).Furthermore,Bambusa bamboswas found totally segregated,showing a strong indication of the polyphyletic origin of the genusBambusa.Similar views have been held by different workers(Soderstrom and Ellis 1988;Loh et al.2000;Das et al.2007;Mukherjee et al.2010).
While it is difficult to identify the root of the divergence among the members of the genusBambusawithout complete genetic information,we could identify the most discriminatory morphological characters that segregated them in separate clusters in the dendrogram.Principle coordinate analysis(PCA)of the replicate morphological data revealed the presence/absence of powdered culm and swollen node are the most discriminatory factors for PCA1(21.5%,Fig.3)that segregated the studied OTUs into two major clusters.Interestingly,four of 10Bambusaspecies(B.pallida,B.tulda,B.assamicaandB.garuchokua),which have the powdered culm and lack of swollen nodes,were clustered away from the rest.However,the same character set was not the entire discriminatory factor to groupD.strictusalong with most of theBambusaspecies.
According to the traditional taxonomic classification of bamboos proposed by Gamble(1896),all the bamboo plants belong to the family Poaceae and recognized as tribe Bambuseae.The genusDendrocalamuswas included in the subtribe Dendrocalameae,while the genusBambusawas placed in subtribe Eubambuseae(Das et al.2007).However,the branching pattern in the dendrogram does not fully concur with the classical taxonomic classification of bamboos.
Conclusion
Phylogenetic and taxonomic difficulties are common within the giant grass,bamboo.The phylogenetic relationship amongst 15 bamboo species obtained from the key morphological descriptors does not fully agree with the traditional taxonomic classification.This discrepancy is due to the unique life cycle of bamboo,which severely limits the availability of the flowering and fruiting characters and thus,relies wholly on vegetative characters for the analysis,while the classical system of classification is based on both vegetative and reproductive character.Thus,the genome may not truly be reflected by just morphological traits(Brown-Guedira et al.2000).In this study,we identified a set of morphological characters that are more discriminatory than the others in defining the phylogenetic relationship,close to the classical viewpoint.Multidisciplinary and molecular approaches are now still needed for the further study and evaluation of the phylogenetic relationships of bamboo species and for accurate taxonomic demarcation.
AcknowledgementsThe authors are thankful to Mr.Jagajit Brahma,Ms.Bijanta Bala Brahma and Karma Goyari,laboratory/ field attendant for helping in maintaining the germplasm and the DBT-BIF centre,MLACW,for necessary software help.
 Journal of Forestry Research2018年5期
Journal of Forestry Research2018年5期
- Journal of Forestry Research的其它文章
- Environmental load of solid wood floor production from larch grown at different planting densities based on a life cycle assessment
- An overview of proven Climate Change Vulnerability Assessment tools for forests and forest-dependent communities across the globe:a literature analysis
- Assessing the vulnerability of a forest ecosystem to climate change and variability in the western Mediterranean sub-region of Turkey:future evaluation
- Effects of management regimes on carbon sequestration under the Natural Forest Protection Program in northeast China
- Mixed-effects modeling for tree height prediction models of Oriental beech in the Hyrcanian forests
- Estimation of a basal area growth model for individual trees in uneven-aged Caspian mixed species forests