
Deposition of litter and nutrients in leaves and twigs in different plant communities of northeastern Mexico
2018-09-07 03:06HumbertoGonzlezRodrguezRoqueGonzaloRamrezLozanoIsraelCantSilvaMarcoViniciomezMezaEduardoEstradaCastillJosRamArvalo
Journal of Forestry Research 2018年5期
Humberto González-Rodríguez•Roque Gonzalo Ramírez-Lozano•Israel Cantú-Silva •Marco Vinicio Gómez-Meza •Eduardo Estrada-Castillón •JoséRamón Arévalo
Abstract Studies on litterfall and decomposition provide estimations of decomposition rates of different ecosystems.This is key information to understanding ecosystem dynamics and changes in a scenario of global warming.The objective of this research was to assess litterfall production,the potential deposition of macro and micronutrients through leaf and twig fall as well as macronutrient—use efficiency in three forest ecosystems at different altitudes:a pine forest mixed with deciduous species(S1);a Quercus spp.forest(S2);and,a Tamaulipan thornscrub forest(S3).Total annual litterfall deposition was 594,742 and 533 g m-2for S1,S2 and S3.Leaf litter was higher(68%)than twigs(18%),reproductive structures(8%)or miscellaneous material(6%).Micronutrient leaf deposition was higher for Fe followed by Mn,Zn and Cu.Macronutrient leaf deposition was higher for Ca followed by K,Mg and P.Even though P deposition in leaves and twigs was lower than other macronutrients,its nutrient use efficiency was higher than Ca,Mg or K.Altitude and species composition determine litter and nutrient deposition,with higher values at mid-altitudes(550 m).Altitude is an important factor to consider when analyzing litter production as well as nutrient deposition as shown in this study.Litter production and nutrient deposition are expected to change in a scenario of global warming.
Keywords Deciduous species·Litter deposition·Leaf/twig litter nutrients·Pine forest·Tamaulipan thornscrub
Introduction
Studies on litterfall and decomposition have become common since the 1960s because of its role in understanding the dynamics of nutrient cycling(Zhou et al.2007),allowing for the estimation and comparison of decomposition rates amongst sites and limiting nutrients of productivity to be determined(Prescott 2010).The concern over global warming in the last two decades associated with rising atmospheric carbon dioxide concentrations makes the understanding of the decomposition process necessary in order to predict the impact of global warming.In thornscrub ecosystems,litter is the largest source of organic material and nutrients in the soil humus layer(Isaac and Nair 2006).It represents a main pathway through which soils,short of nutrient uptake and percolation,are recharged.In addition,litter represents one of the first relationships between producer and decomposer organisms.Microbial decomposition plays a crucial role to make nutrients available easily for plants.However,this process requires adequate temperature and humidity since a lack of moisture inhibits decomposition(Nakane 1975).
Leaf litter is the major component of aboveground litterfall in thornscrub ecosystems(González-Rodríguez et al.2011).Leaf litter was highly correlated with spatio-temporal factors(López-Hernández et al.2013).In addition,annual litterfall can vary considerably and is related to weather conditions that differ from year to year(González-Rodríguez et al.2013).Mineral concentrations in foliar litter are affected by factors such as species,soil properties,climate,and tree vigour(Lindroos et al.2006;Ukonmaanaho et al.2008).Nutrient pathways and use efficiencies are critical characteristics when considering the adaptability of plant species to different conditions present in thornscrub stands undergoing restoration(Lugo et al.2004).Nutrient-use efficiency(NUE;[(leaves g m-2)/(nutrients in leaves g m-2)],according to Vitousek(1982),can be used as an index of efficiency(Binkley et al.2004).Litter composition correlates with nutrient availability because decomposition of the dominant foliage type controls mineralization rates(McClaugherty et al.1985).
In this study,the hypothesis is that,at lower altitudes and with high temperature and adequate rainfalls,the nutrient deposition is higher due to a rapid turnover.The objective is to assess litterfall production,the deposition of macro and micronutrients through leaf and twig fall as well as macronutrient-use efficiency in three forest ecosystems representing an altitudinal gradient in the northeastern region of Mexico.Different altitudes,which generate different forest types,soils,and meteorological conditions,are important tools in the analysis of processes like litter production and nutrient efficiency.The three sites are assumed to be in a stationary state and as litter production is related to net primary production(NPP),we will be able to infer nutrient limitations at these sites.
Materials and methods
Study area
This study was carried out at three undisturbed sites in the state of Nuevo Leon,northeastern Mexico.Site 1(Bosque Escuela,S1)is in aPinus pseudostrobusLindl.forest mixed with deciduous species at the Experimental Forest Research Station of the Universidad Autónoma de Nuevo León in the Sierra Madre Oriental Mountain range,Iturbide county,(24°43′N;99°52′W;and 1600 m,a.s.l.).The mean annual temperature is 13.9°C and the average annual rainfall approximately 639 mm.Soils are mainly rocky and comprise Upper Cretaceous lutite or siltstone.Site 2(Crucitas,S2)is situated in the ecotone of aQuercusspp.forest and the Tamaulipan thornscrub(24°46′N;99°41′W;and 550 m,a.s.l.).Mean annual temperatures are 21°C and average annual rainfall 755 mm(González-Rodríguez et al.2004).Site 3(Campus,S3)is at the Experimental Research Station of the Faculty of Forest Sciences of the Universidad Autónoma de Nuevo León in a Tamaulipan thornscrub or subtropical thornscrub woodlands(24°47′N;99°32′W;and 350 m,a.s.l.).Mean annual temperatures range from 14.7 °C in January to 22.3 °C in August,although daily high temperatures of 45°C are common during the summer.The average annual precipitation is 805 mm with a bimodal distribution.S2 and S3 are located in Linares County and the soils are deep,dark-gray or lime-gray vertisols with montmorillonite,which shrink and swell in response to changes in soil moisture content.Mean monthly temperatures(°C)and cumulative monthly rainfall(mm)during the sampling period are shown in Table 1.
The plant community revealed high tree species richness at S2(28 species),dominated byHavardia pallens(Benth.)Britton&Rose,Quercus virginianaMill.andRandia rhagocarpaStandl.(40%of the accumulative importance value),followed by S3 (22 species)dominated byBernardia myricifolia(Scheele)S.Watson,Forestieria angustifoliaTorr.andH.pallens(36%of accumulative importance value),and finally S1(14 species),dominated byP.pseudostrobus.andQuercus canbyiCory and Parks(50%of accumulative importance value).Further information about the three study sites is provided in Ramírez-Lozano et al.(2013).
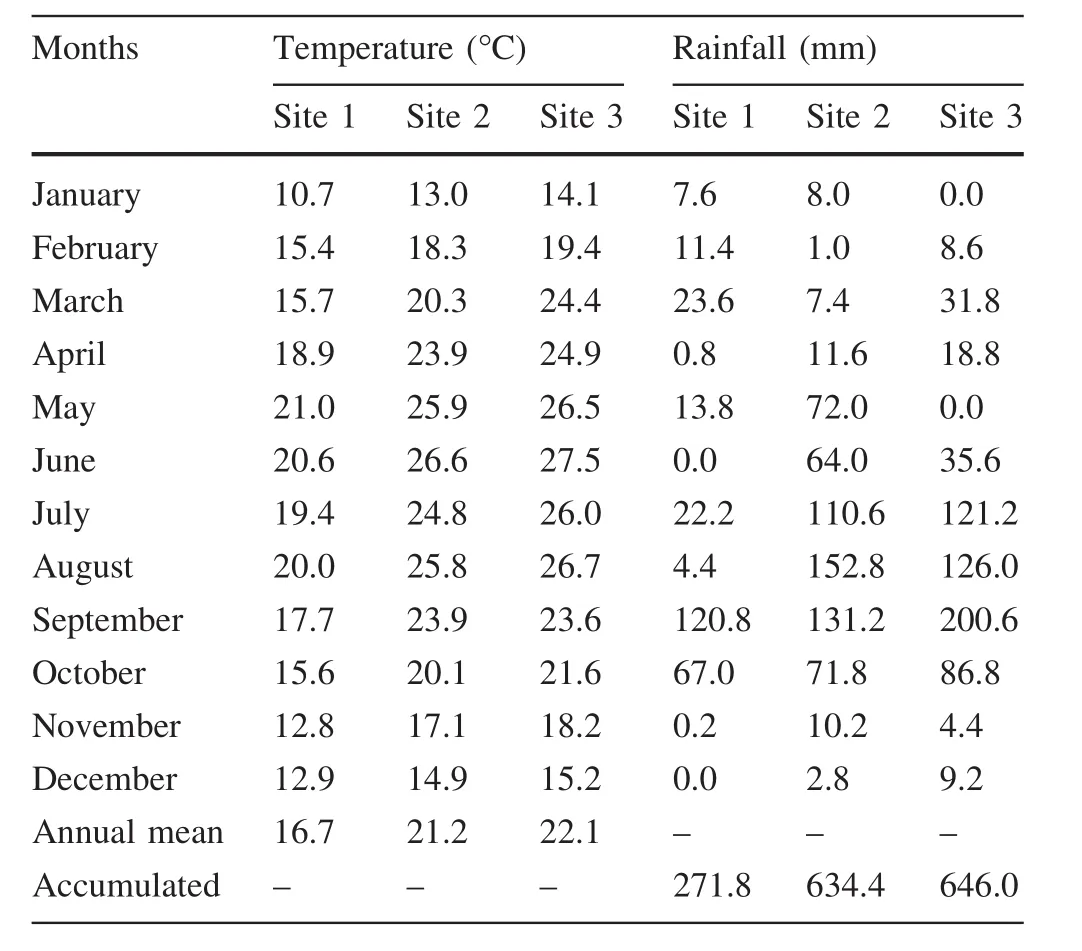
Table 1 Mean monthly air temperature(°C)and cumulative monthly rainfall(mm)during 2008
Litterfall collections
Ten litterfall trays(a 1 m×1 m basket)with wooden sides fitted with nylon netting at the bottom(1 mm mesh size)were randomly located(2500 m2)at each site,and used for collection(all trays were at least 10 m apart).Tray contents were collected at 15-day intervals between January and December 2008.Litter content was manually sorted into:leaves,reproductive structures( flowers,fruits and seeds),twigs(<2 cm in diameter),and miscellaneous residues(unidentified materials— fine plant tissue such as bark,pieces of insect bodies or feces).Samples were oven dried to a constant weight at 65°C for 72 h.Dry samples were ground in a Wiley mill(Thomas Scientific)to pass through a 1.0 mm mesh sieve and kept in closed paper envelopes.
Chemical composition of samples
Leaf and twig litter samples were subjected to mineral analysis by incinerating samples in a muffle oven at 550°C for 5 h.The ash was digested in a solution of HCl and HNO3using the wet digestion technique(Cherney 2000).Ca(oxide nitrous/acetylene flame),K,Mg,Cu,Fe,Mn,and Zn (air/acetylene flame)were estimated by atomic absorption spectrophotometry(Varian,model SpectrAA-200).P was determined spectrophotometrically using a Perkin-Elmer spectrophotometer(Model Lamda 1A)at 880 nm(AOAC 1997).Nutrient deposition at each site was calculated by multiplying leaf or twig litter production on each sampling date by nutrient concentration for the same collection date and site,and adding together over the year.The accumulated values were an estimate of annual nutrient deposition.Leaf litter annual deposition values of Ca,K,Mg and P were used to estimate nutrient use efficiency(LLNUE),defined as the ratio of annual litter mass(leaf litter g ha-1)to annual litter nutrient content(leaf nutrient g ha-1)(Vitousek 1982;Read and Lawrence 2003).
Statistical analyses
We performed a one-way,distance-based permutational ANOVA(Anderson et al.2008)for the comparison of annual litter production(ten litter trays)in the different categories(leaves,twigs,reproductive structures,miscellaneous,and total litter)between the three sites S1,S2 and S3.The same analysis was done for LLNUE values of Ca,K,Mg and P.Pairwise posteriori comparisons using t-statistics were applied to significant factors and interactions.The analyses were based on Euclidean distances of the raw data,withpvalues(p<0.05)obtained with 9999 permutations and a Monte Carlo correction where necessary.Primer 6 and Permanova+(PRIMER-E Ltd,Plymouth,UK)were used to perform all PERMANOVA statistical procedures.
Principal Components Analysis was used(standardized PCA,using CANOCO;ter Braak and Šmilauer 1998)to examine the nutrient litter composition.In this analysis,we used the annual deposition values(mg m-2a-1)of micro and macronutrients,discriminating among leaves and twigs for the three studied sites.Percentages of the nutrients in the leaves and twigs were used to analyze nutrient composition and not absolute values(much higher in leaves than twigs).
Results
Yearly litterfall deposition revealed differences for the four fractions analyzed(Fig.1),however,pairwise analyses after permanova,only revealed differences between sites in the leaves(Fig.1a),twigs(Fig.1b)and miscellaneous materials(Fig.1d),with higher annual values for leaves and twigs at S2,and higher values for miscellaneous materials(lower values of litter deposition)at S1.However,leaves become the main fraction and determined that total litterfall(for all fractions)was significantly different between sites(although no pairwise differences were found).
Monthly deposition of leaves(Fig.2a),twigs(Fig.2b),reproductive structures(Fig.2c),miscellaneous materials(Fig.2d)and total litterfall(Fig.2e)reveal differences in most months.
The dynamics of leaf deposition in S1 ranged from 18.1 g m-2in June to 56.6 g m-2in May.At S2,this fraction was 19.9 in July and 65.1 in March.At S3,the values varied from 10.6(July)to 55.7(January;Fig.2a),with higher values during the periods February to May and October to November.Twig deposition ranged from 0.9 g m-2(October)to 17.0(February)g m-2in S1.At S2,it varied from 7.6(October)to 34.9(May)and at S3,the variation was from 3.2 in September to 22.7 in March(Fig.2b),with higher values found between March and May.The deposits of reproductive structures at S1 ranged from 0.6 g m-2(January)to 12.6(May)g m-2.At S2,they varied from 1.0 in January to 10.1 in April,and at S3,from 0.3(February)to 4.9(August)(Fig.2c).Miscellaneous materials(feces,insects and unknown materials)at S1 fluctuated from 1.4 g m-2in April to 16.7 g m-2in May.At S2,it varied from 0.3(February)to 7.5(March),while at S3,it varied from 0.3 in January to 1.8 in October(Fig.2d).
Total litterfall(leaves,twigs,reproductive structures and miscellaneous materials)at S1 varied from 20.0 g m-2(October)to 91.5 g m-2(May).At S2,it ranged from 35.3(July)to 89.2(March),while at S3 litterfall fluctuated from 16.6(April)to 71.9(October)(Fig.2e).Total litterfall followed the same trend as leaf deposition.
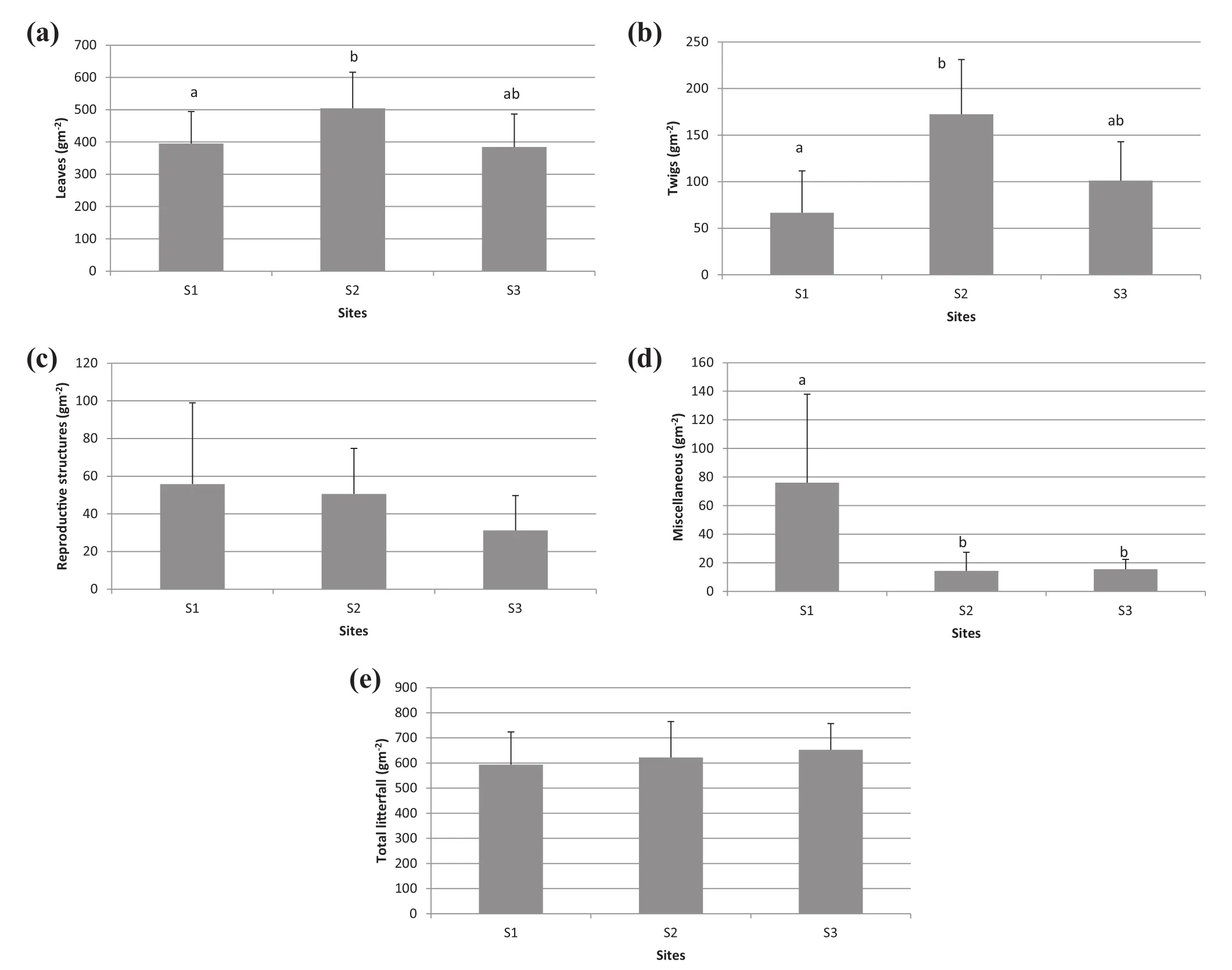
Fig.1 Annual litterfall deposition(g m-2)for leaves,twig,reproductive structures,miscellaneous and total litter for S1,S2,and S3.Bars reflect standard deviation.Identical letters above the bars indicate non-significant differences.(Permanova results:Leaves Pseudo-F2,27=4.05, p<0.03; Twigs Pseudo-F2,27=12.09,p<0.001;Reproductive Structures Pseudo-F2,27=1.07,non-significant;Miscellaneous Pseudo-F2,27=9.2,p<0.001 and total litterfall Pseudo-F2,27=0.53,non-significant)
In the case of leaf and twig deposition,S1 revealed lower nutrient values,higher values at S2,except in the case of Fe for leaves which was higher at S3.These values were in some cases much more elevated as in the case of Mn or Ca(Table 2).An analysis of nutrient composition(standardized in percent)for the three sites discriminating between leaves and twigs,revealed again all the nutrients are dominant in S2 for leaves and twigs(Fig.3).
The annual deposition of Cu was higher at S2 than at S3 or S1(Table 2).Mn was higher at S2 compared to the other sites.Deposition of leaf Zn was similar among sites.Leaf micronutrient deposition occurred as:Fe>Mn>Zn>Cu(Table 2).Total annual deposition of micronutrients(Cu+Mn+Fe+Zn) was 180.3, 138.1 and 133.9 mg m-2at S2,S3 and S1,respectively.Total annual deposition of macronutrients(Ca+K+Mg+P)for S1,S2 and S3 were 5.6,16.6 and 18.8 g m-2,respectively.
Leaf litter nutrient use efficiency(LLNUE)for Ca,Mg,K and P exhibited the order:S1>S2>S3(Table 3)and were significantly different.Among sites,LLNUE was higher for P followed by Mg,K,and Ca.
Discussion
The three forest ecosystems differed significantly not only in terms of the contribution of different litterfall fractions either on a monthly or year basis,but also in terms of the potential nutrient deposition through leaves and twigs and nutrient use efficiency.These differences have been previously documented in different forest ecosystems(González-Rodríguez et al.2013;Mendoza et al.2014;Lopes et al.2015).
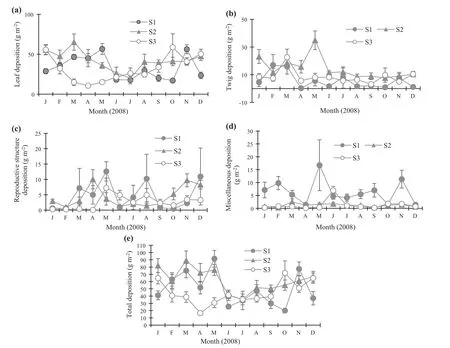
Fig.2 Litterfall monthly deposition for leaves(a),twigs(b),reproductive structures(c),miscellaneous(d)and total litter(e).Plotted values are average(n=10)±standard errors.Site 1( filled circle).Site 2(triangle).Site 3(open circle)
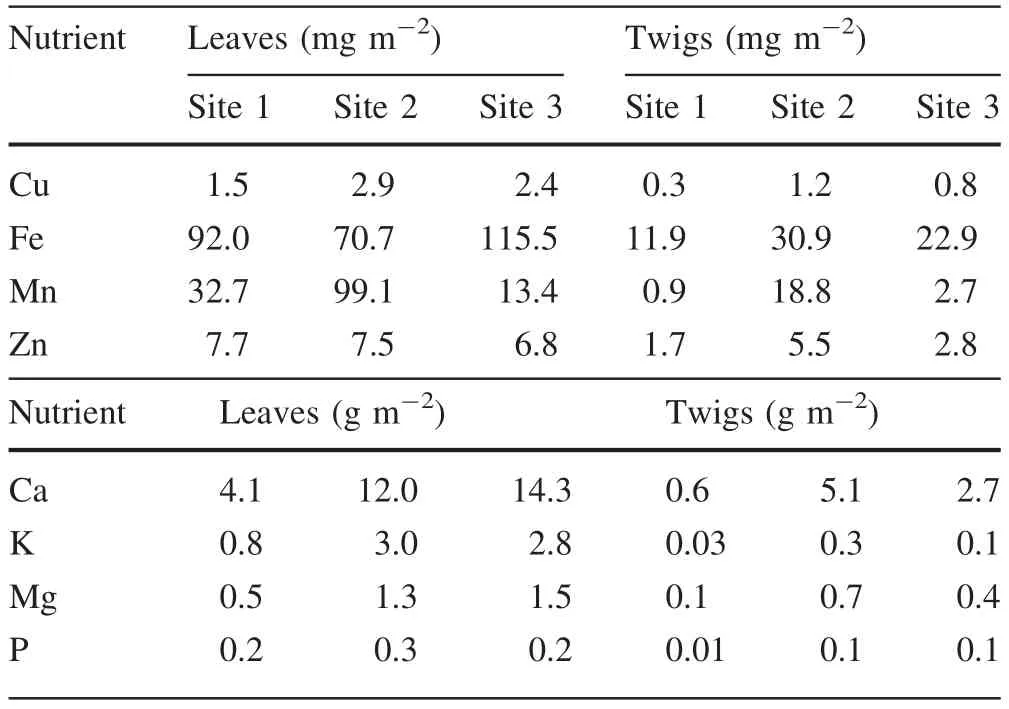
Table 2 Annual deposition of micro(Cu,Fe,Mn and Zn)and macronutrients(Ca,K,Mg and P)in fallen leaves and twigs
Site S2 had the highest total annual litterfall deposition followed by S1,and S3.The same pattern occurred for leaves and twigs.However,annual reproductive structures and miscellaneous materials depositions were higher in S1(Fig.1).These values are within the range of litterfall deposition observed in studies carried out in the same region(González-Rodríguez et al.2011;López-Hernández et al.2013)of the microphyllous desert scrubland of northeastern Mexico(González-Rodríguez et al.2013),and in a xerophyllous community in Spain(Merino 1983).However,lower quantities(202.7,100.2 and 35.2 g m-2)were reported by Martínez-Yrízar et al.(1999)in three Sonora Desert communities,northern Mexico.
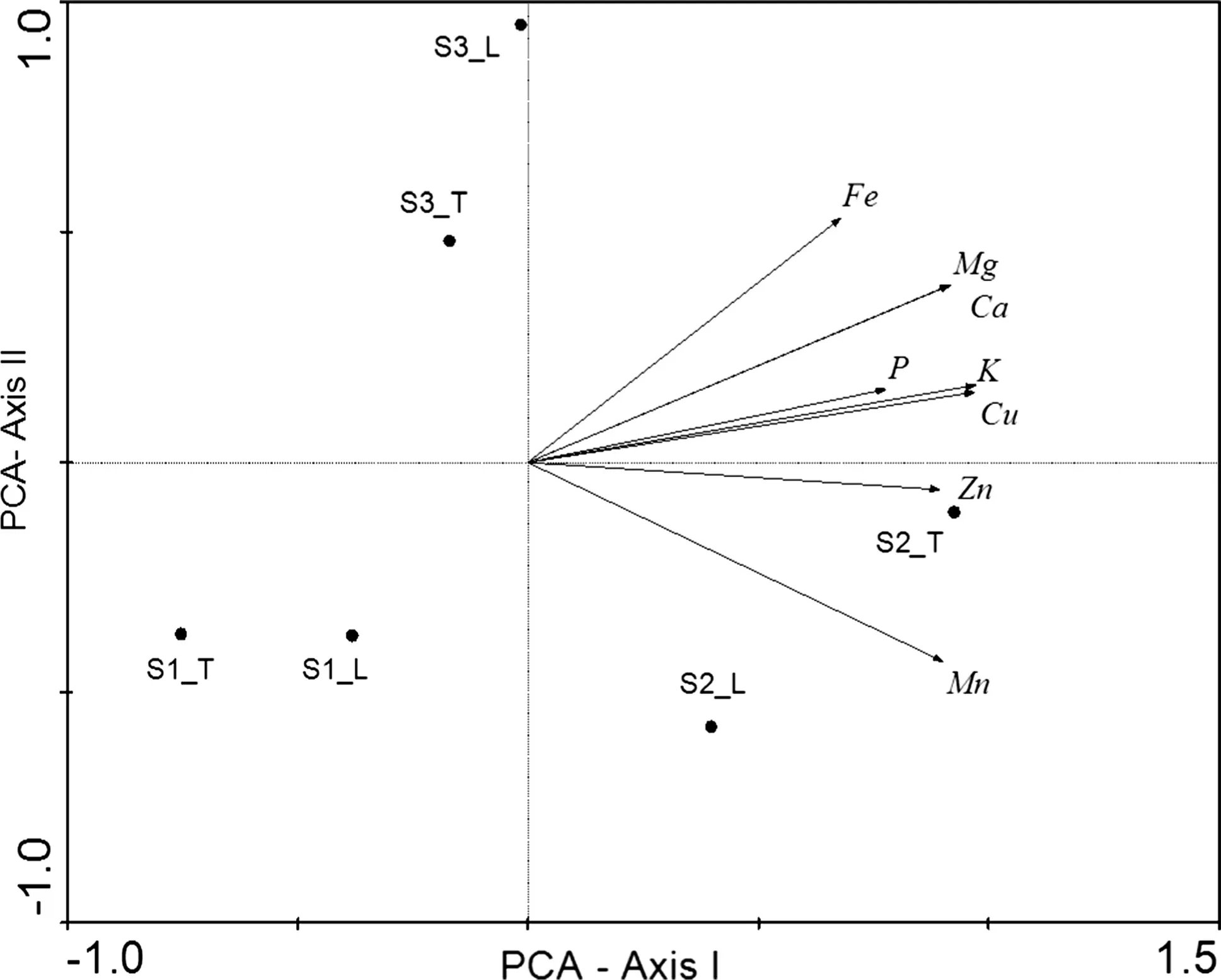
Fig.3 Site and nutrient scores for leaf and twig deposition.Principal components analysis of the three sites(‘‘L’’following the site means annual leaf nutrient composition and ‘‘T’’average twig nutrient composition).Macro and Micronutrients appear as arrows and the sites as symbols(eigenvalues for axis I:0.82,eigenvalue of axis II:0.12,cumulative percentage of variance explained for axes I and II:94%)
In this study,leaves represented the main component of litterfall(Caritat et al.2006;Santa-Regina and Tarazona 2001a,b;Patricio et al.2012;Lopes et al.2015).The percentage of leaves produced with respect to the total biomass that returns to the soil depends on the state of development and age of the plant,achieving the main contribution in periods of higher litterfall production,coinciding with the highest rates of photosynthesis and respiration(Martínez-Alonso et al.2007).Moreover,differences in leaf fall in S2 and S3 compared to S1 could be related to the continuous shedding of leaves of deciduous species,a characteristic of the Tamaulipan thornscrub ecosystem which is a morphological and physiological mechanism to cope with water deficits and high temperatures to avoid tissue dehydration and water loss through transpirational flux(González-Rodríguez et al.2004).
Differences in the dynamics of litterfall deposition among sites also may be due to variations in species composition.However,temperature and rainfall are the main determinants of the species composition in this region(Estrada et al.2015).In a study carried out in the semiarid region of Brazil,litterfall production was found to be related to tree density but no link was found to species richness.The observed temporal heterogeneity of litterfall production demonstrates a strong link between rainfall and the dynamics of nutrient cycling(Lopes et al.2015;Allen et al.2012).
We suggest that climatic conditions in the study area influenced litterfall deposition.Higher deposition occurred in months with lower rainfall and lower temperatures(Table 1).Similar responses were reported by López-Hernández et al.(2013)and by González-Rodríguez et al.(2013).Moreover,Martinez-Alonso et al.(2007)indicated that increases in temperatures and dry periods may influence the dynamics of productivity in warm,temperate ecosystems.
Leaf and twig micronutrient(Fe>Mn>Zn>Cu)and macronutrient(Ca>K>Mg>P)depositions were similar to those reported by López-Hernández et al.(2013)and Gonzalez-Rodriguez et al.(2013)for thornscrub and microphyllous vegetation in northeastern Mexico,and by Andivia et al.(2012)for aQuercus suberL.ecosystem in the Mediterranean area.In this study,S2 was the dominant site for litter production as well as for macronutrients and micronutrients(Fig.3).
Although P deposition in leaves and twigs was lower than for other macronutrients,its nutrient use efficiency was relatively higher than Ca,Mg or K.A similar trend has been reported by del Valle-Arango(2003),Swamy et al.(2004),Safou-Matondo et al.(2005),González-Rodríguez et al.(2011)and López-Hernández et al.(2013)for different ecosystems.It has been suggested that P could be translocated to other plant structures before leaf senescence to contribute to the generation of new plant structures or physiological processes(Palma et al.2000).Also,P may be considered the limiting nutrient at the three sites,followed by Mg and K.In addition,values of LLNUE are significantly higher at S1,with an indication of lower availability of these nutrients(also related to the lowest species richness).
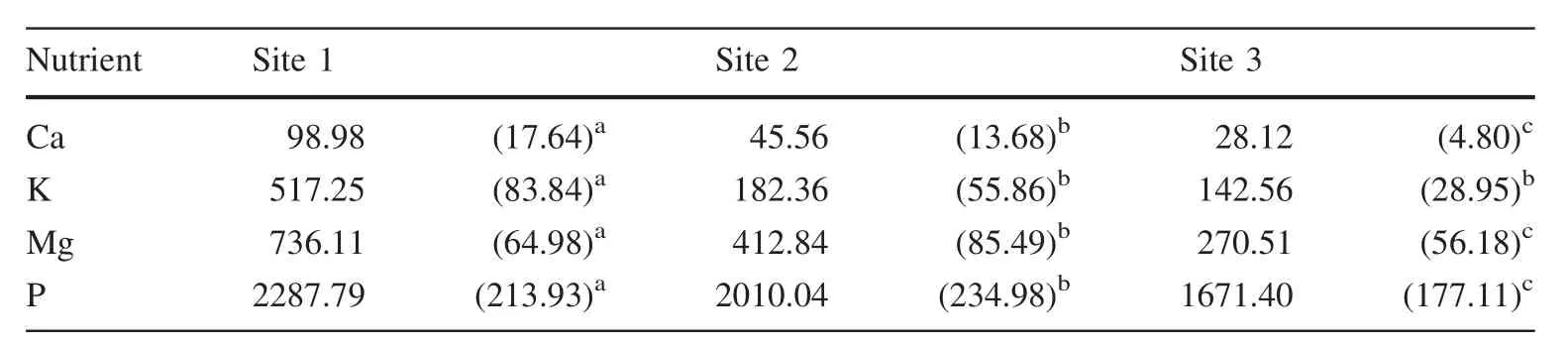
Table 3 Leaf litter(g ha-1)nutrient use efficiency(LLNUE)of leaf macronutrients(Ca,K,Mg and P;average and standard deviation,different letters a,b or c,indicate significant differences in leaves of the three sites)
In general,larger deposition of leaves and twigs indicate higher potential mineral depositions.In addition,higher rainfall in S2 and S3(Table 1)might have increased the mineral depositions in S2 and S3 compared to S1(Ramírez-Lozano et al.2013).Increase in mineral content due to higher rainfall has also been reported by Ramírez-Lozano et al.(2010)who evaluated mineral profiles of native shrubs from northeastern Mexico.Altitude is an important element to consider when analyzing litter production and nutrient deposition as shown in this study.It is also important to note that the nature of litter chemistry(different species composition along the altitudinal gradient)also plays an important role in decay and decomposition rates.
This study reveals differences in litterfall deposition and nutrient composition among the sites(S1:Pinus pseudostrobusLindl.forest mixed with deciduous;S2:an ecotone of aQuercusspp.forest and the Tamaulipan thornscrub and S3:Site 3:a subtropical thornscrub woodland)with higher values for S2(mid-altitude with respect to the other two sites).S2(Quercusspp.ecosystem)has climatic conditions that enhance plant productivity and this is reflected in higher litterfall and levels of nutrient use efficiency,as well as greater species richness due to the environmental conditions and microenvironment heterogeneity.Based on LLNUE,S1(pine-deciduous spp.mix)has more limited productivity that can be the result of its low species richness.Also,the P deposition in leaves and twigs had higher nutrient use efficiency than other macronutrients.Differences in the potential mineral return to the soil may be related to rainfall,soil fertility,botanical composition,litter quality and plant parts which influence nutrient use efficiency(Lambers et al.2000).In a Korean pine-mixed forest,researchers found that temperature drives litter fall and nutrient cycles,and there is a lineal variation together with the altitudinal gradient(Zhou et al.2015),while in a mixed forest(Guangdong province in Southern China),studies indicated that precipitation played an important role in litter production(Zhou et al.2007).
We consider that temperature and rainfall variations,following the Intergovernmental Panel on Climate Change(IPCC)predictions,will become important drivers of change in plant communities in the future.In fact,climatic changes predicted by the IPCC are likely to lead to significant changes in forest composition.Even moderate scenarios of the IPCC predict a severe decrease in precipitation and a rise of 3–4 °C in average temperatures(Christensen et al.2007).Specifically,modifications in rainfall and runoff,and a replacement of semiarid by arid vegetation are expected in northern Mexico due to the effects of land use and climatic changes(Bates et al.2008),leading to decreases in species richness.
AcknowledgementsValuable technical assistance provided by Elsa Dolores González Serna and Manuel Hernández Charles is gratefully recognized.The authors appreciate and wish to thank the land owner of site 2 for providing the facilities to establish the experimental sites.This research was funded in part by Universidad Autónoma de Nuevo León(PAICYT Grant CT259-15)and Consejo Nacional de Ciencia y Tecnologıá(Grant 250732).
 Journal of Forestry Research2018年5期
Journal of Forestry Research2018年5期
- Journal of Forestry Research的其它文章
- Environmental load of solid wood floor production from larch grown at different planting densities based on a life cycle assessment
- An overview of proven Climate Change Vulnerability Assessment tools for forests and forest-dependent communities across the globe:a literature analysis
- Assessing the vulnerability of a forest ecosystem to climate change and variability in the western Mediterranean sub-region of Turkey:future evaluation
- Effects of management regimes on carbon sequestration under the Natural Forest Protection Program in northeast China
- Mixed-effects modeling for tree height prediction models of Oriental beech in the Hyrcanian forests
- Estimation of a basal area growth model for individual trees in uneven-aged Caspian mixed species forests