
Comparable water use of two contrasting riparian forests in the lower Heihe River basin,Northwest China
2018-09-07 03:06TengfeiYuQiFengJianhuaSiXiaoyouZhangHaiyangXiChunyanZhao
Journal of Forestry Research 2018年5期
Tengfei Yu•Qi Feng•Jianhua Si•Xiaoyou Zhang•Haiyang Xi•Chunyan Zhao
Abstract Understanding forest ecosystem evapotranspiration(ET)is crucial for water-limited environments,particularly those that lack adequate quantified data such as the lower Heihe River basin of northwest China which is primarily dominated by Tamarix ramosissima Ledeb.and Populus euphratica Oliv.forests.Accordingly,we selected the growing season for 2 years(2012 and 2014)of two such forests under similar meteorological conditions to compare ET using the eddy covariance(EC)technique.During the growing seasons,daily ET of T.ramosissima ranged from 0.3 to 8.0 mm day-1with a mean of 3.6 mm day-1,and daily ET of P.euphratica ranged from 0.9 to 7.9 mm day-1with a mean of 4.6 mm day-1for a total of 548 and 707 mm,respectively.The significantly higher ET of the P.euphratica stand was directly linked to high soilevaporation rates under sufficient water availability from irrigation.When the soil evaporation was disregarded,water use was comparable to two contrasting riparian forests,a P.euphratica forest with a total transpiration of 465 mm and a T.ramosissima forest with 473 mm.Regression analysis demonstrated that climate factors accounted for at least 80%of ET variation in both forest types.In conclusion,water use of the riparian forests was low and comparable in this arid region,that suggest the long-term plant adaptation to the local climate and conditions of water availability.
Keywords Eddy covariance·Evapotranspiration·Heihe River basin·Riparian forest
Introduction
Evapotranspiration(ET)is a major component of catchment water balances.BecauseETin most large basin ecosystems is limited by rainfall,riparian corridors are hot spots for atmospheric water loss for large arid-and semiarid basins(Sala et al.1996;Smith et al.1998).In the lower Heihe River basin(HRB),northwestern China,riparian forests are dominated by phreatophytes,such asPopulus euphraticaOliv.tree species andTamarix ramosissimaLedeb.shrub species(Si et al.2007).Phreatophytes are deep-rooted plants that obtain their water from the water table or the soil above it.Over the past several decades,the eco-environment of riparian forests has markedly deteriorated to the extent that riparian forests have decreased in size and their ecological functions have declined(Cheng et al.2014;Feng and Cheng 1998;Si et al.2007).In order to restore riparian forests in the lower HRB,the Ecological Water Conveyance Project(EWCP)was initiated in 2000 by the government of China to rechargewater back into the lower HRB(Guo et al.2008).AlthoughP.euphraticaforests were maintained after this water conveyance program was put into practice (from 295.69 km2in 1982 to 294.30 km2in 2010),T.ramosissimaforests have expanded in area(from 1003.71 km2in 1982 to 1529.56 km2in 2010).Accordingly,it is important to quantify water consumption by these two forest types in order to effectively utilize their limited water resources.

Table 1 Mean values±SD of meteorological variables,including total precipitation (P,mm),mean daylight hours(N,h),air temperature(Ta,°C),relative humidity(RH,%),saturated vapor pressure deficit(VPD,kPa),and wind speed(U,m s-1)from the Ejin meteorological station for the years 2012 and 2014
AlthoughP.euphraticais an important structural tree species of riparian forests,only a limited number ofETstudies have been conducted in this region.Studies mainly focused on a tree-level scale(Si et al.2007;Zhang et al.2006)and rarely on a stand-level scale(Hou et al.2010).Annual transpiration measured through sap flow was 214.9 mm (Si et al.2007),less than the totalET(447.0 mm)estimated by the Bowen ratio energy balance(BREB)method at the same site(Hou et al.2010).In addition,T.ramosissimais a native phreatophytic that originates from arid regions in Central Asia,and has widely invaded numerous western states(Di Tomaso 1998;Hatler and Hart 2009;Shafroth et al.2005).Generally,T.ramosissimais recognized as a prolific water user.Data taken from flux towers(Cleverly et al.2002,2006;Devitt et al.1998)and large area estimates using remote sensing techniques(Bawazir et al.2009;Dennison et al.2009;Nagler et al.2009)have yielded moderate water use rates(from 800 to 1450 mm year-1).In contrast to the western U.S.,ETfromT.ramosissimastands was approximately 500 mm year-1in the lower Tarim River basin(Yuan et al.2014)and only 280 mm year-1in the lower HRB in northwestern China(Si et al.2005).It may therefore be deduced thatETfromT.ramosissimastands is lower in its native regions than in regions it has invaded,but it remains unclear whetherT.ramosissimastands have higherETrates than other native species(e.g.,P.euphratica).
In most previously published studies,the eddy covariance(EC)method has been widely used to estimateETusing direct measurements of latent heat fluxes(Sun et al.2008).The objectives of this study were to determine the magnitude ofETforP.euphraticaandT.ramosissimastands in the lower HRB,and compared them to other regions around the world.For this purpose,we gathered nearly 2 years of EC observations from two stands in the lower HRB.In addition,we concurrently observed meteorological variables and groundwater tables in the same plot used to determine environmental controls ofET.
Materials and methods
Study site
The climate in this region is extremely arid with a mean annual precipitation of only 37.4 mm,of which>75%occurs between June and August.Annual pan evaporation is 3390.3 mm,which is greater than precipitation by a factor of 90.Mean annual temperatures in July and January are 27.0 and-11.7°C,respectively,as recorded at the Ejin meteorological station between 1959 and 2011(Yu et al.2013b).Mean values±standard deviation(SD)of meteorological conditions for 2012 and 2014 are provided in Table 1.Given that both the highest rainfalls and temperatures occur concurrently in this region,most rainfall is depleted by evaporation before it in filtrates into the soil.Therefore,the effect of rainfall on the water balance is minimal.In addition to rainfall,groundwater recharged by runoff from the Heihe River sustains both local residents and regional ecosystems(Si et al.2014).
We selectedT.ramosissimaandP.euphraticastands,two typical forest types in arid regions,to conduct EC measurements.TheT.ramosissimastand (42°01′N,101°03′E;926 m a.s.l.)is situated in a north–south direction in the Alxa Desert Ecohydrology Experimental Research Station,Chinese Academy of Sciences.TheP.euphraticastand(41°59′N,101°10′E;920 m AMSL)is located at theP.euphraticaForest National Nature Refuge at Qidaoqiao(Fig.1).We initially conducted a comprehensive quadrat sampling of vegetation in 2010 for both forest types(Table 2).Leaf area index(LAI)of canopies was measured using a plant canopy analyzer(LAI-2200,LI-COR Biosciences,Inc.,Lincoln,NE,USA)with one wand for both above-and below-canopy measurements under diffused light conditions during sunrise and sunset.This was carried out approximately once every month throughout the growing season in both years.The understory was composed of shallow-rooted grasses,predominantlySophora alopecuroidesL.,Karelinia caspica(Pall.)Less.,andAchnatherum splendens(Trin.)Nevski.Soil characteristics from theT.ramosissima(Yu et al.2013a)andP.euphraticastands were determined(Gao et al.2016b).
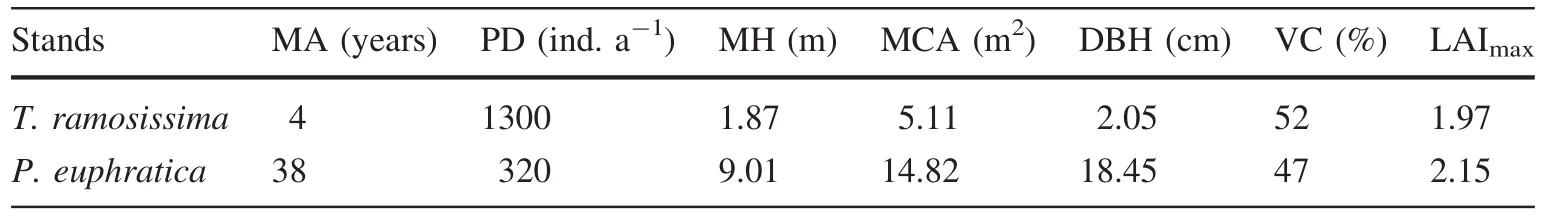
Table 2 Statistical features of the T.ramosissima and P.euphratica stands
Flux and meteorological measurements
Momentum flux,sensible heat(H,W m-2),and latent heat(LE,W m-2)were measured using a three-dimensional sonic anemometer(CSAT3,Campbell Scientific,Inc.,North Logan,UT,USA)and an open path CO2/H2O analyzer(LI-7500A,LI-COR Biosciences,Inc.,USA).In addition,meteorological variables were recorded,including net radiation(Rn,W m-2),air temperature(Ta,°C),and relative humidity(RH,%)in conjunction with online processed 0.5 h mean horizontal wind speeds(U,m s-1)and CO2/H2O flux rates using the CR3000 datalogger(Campbell Scientific Inc.,USA).Rnwas determined using a four-component net radiometer(CNR4,Kipp&Zonen,Delft,the Netherlands),whileTaandRHwere measured using a relative humidity and temperature sensor probe(HMP45C,Campbell Scientific,Inc,USA).The vapor pressure deficit(VPD,kPa)was calculated based onTaandRH(Campbell and Norman 1998).Soil heat flux(G,W m-2)was measured using a heat flux plate(HFP01,Hukse flux Thermal Sensors B.V.,Delft,the Netherlands)at a 5 cm depth dispersed 0.5 m belowground horizontally.We conductedT.ramosissimastand flux and meteorological measurements from May 2011 to November 2012 at 3 m above-ground level.Subsequently,this method was applied to a 20 m high flux tower to measureP.euphraticastand flux and meteorological measurements from June 2013 to the present(Gao et al.2016b).To compare,we selected the entire 2012 and 2014 growing seasons(i.e.,from May to September)under similar meteorological conditions for analysis.In addition,two wells 30 m apart from the EC tower were installed and groundwater depth(GD,m)measured at 5 day intervals during the growing seasons,which provided results of a maximum of 2.65 m for theT.ramosissimastand and 2.25 m for theP.euphraticastand.Water was introduced via the Heihe River EWCP(set up to provide local herdsmen a means to irrigate their pastures)to theP.euphraticastand after September 16,2014.
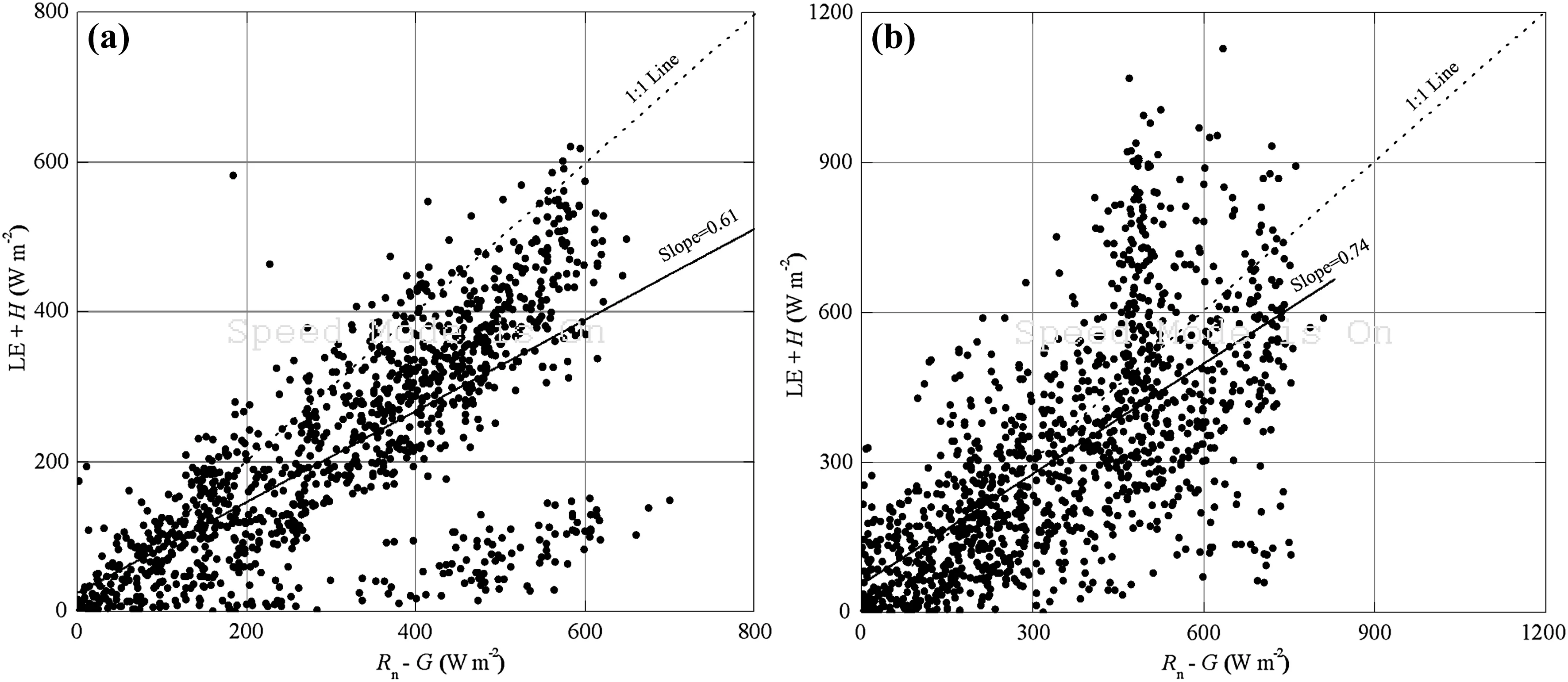
Fig.2 Energy balance closures of eddy covariance(EC)for T.ramosissima(a)and P.euphratica(b)during 2012 and 2014,respectively.The 1:1 line(dotted line)and linear regression(solid line)between Rn-G and LE+H is shown
Data processing and gap filling
Raw CSAT3 and CO2/H2O flux data were processed using EddyPro software(LI-COR Biosciences,Inc.,USA),which includes features such as double coordinate rotation,block average detrending,time lag compensation via covariance maximization (with defaults),and density fluctuation compensation via the use/convert features to the mixing ratio(Burba et al.2012).Statistical analysis of the raw time series data was carried out,including spike count/removal,amplitude resolution,dropouts,absolute limits,and skewness and kurtosis,according to Vickers and Mahrt(1997).Quality checks/ flagging were conducted according to the CarboEurope standard(Foken et al.2004).After conducting a detailed post production on data,HandLEwere calculated to obtain a 0.5 h average.We screened 0.5 h turbulent fluxes and meteorological variables to remove spurious data points caused by sensor malfunctions,rain events,and sensor maintenance.
The energy balance was analyzed from the EC data that includeRn,LE,andHandGcalculated from the average of the two heat flux plate measurements.We disregarded canopy storage heat and photosynthetic energy consumption due to the sparseness of canopy cover.The relationship between the half-hourly data of net radiation subtracted by the ground heat flux(Rn-G)and the half-hourly data of the total turbulent flux(LE+H)indicated that EC underestimated turbulent fluxes by 22%(slope=0.78;R2=0.91)and 21%(slope=0.79;R2=0.80)for theT.ramosissimaandP.euphraticastands,respectively(Fig.2).LEdata was also removed from the energy unbalance.In total,34 and 19%of the originalLEdata was rejected during data quality processing of theT.ramosissimaandP.euphraticastands,respectively.
Gap filling was carried out when totalEThad to be calculated by filling shorter gaps(i.e.,<2 h)in the data through linear interpolation,and longer gaps inLEflux using energy balance analysis.If EC measuredLEwas greater thanRn+G-H or null,we then forcedLEto close according to the energy balance analysis(Yuan et al.2014),that is,LE=Rn+G-H,otherwiseLEwas retained.ET(mm h-1)fromLEwas calculated hourly according to Yuan et al.(2014),the actualET(mm h-1)from combined data with EC measuredET(ETEC,mm h-1),and the estimatedETfrom the energy balance analysis(ETEB,mm h-1)(Fig.3).When hourlyETwas negative,it was set to zero.DailyET(mm day-1)was the sum of hourlyETduring a single day.In addition,the daily referenceET(ET0,mm day-1)was calculated from the FAO Penman–Monteith equation(Allen et al.1998)on a daily scale.RnandGdata were used to calculate dailyET0derived from observations from flux tower measurements,and other daily data from the Ejin meteorological station.The crop coefficient(Kc=ET/ET0)was then determined.
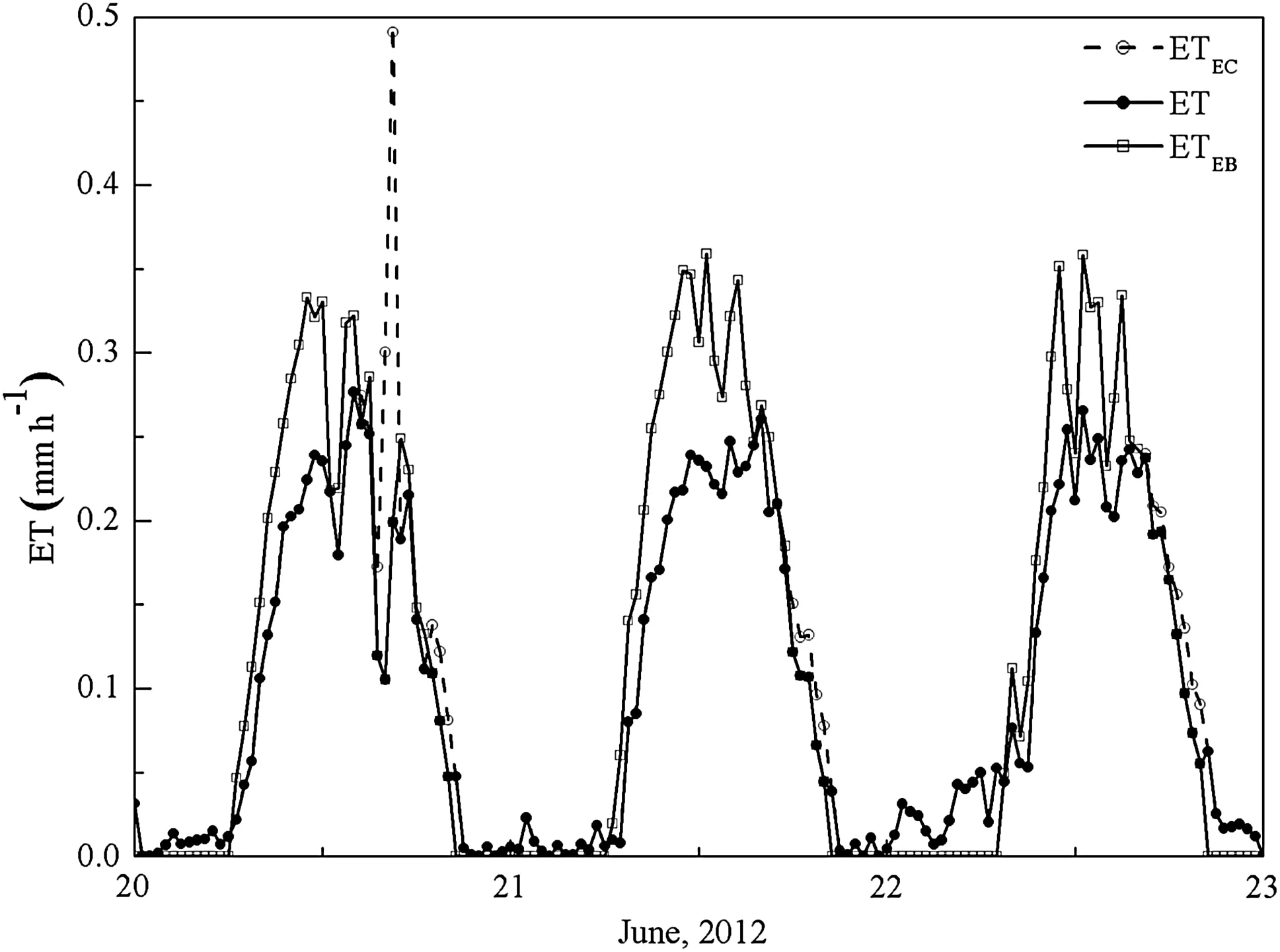
Fig.3 Hourly variation in measured evapotranspiration from eddy covariance(EC)(ETEC,mm h-1),estimated evapotranspiration derived from energy balance analysis(ETEB,mm h-1),and actual evapotranspiration calculated from the combination of the two(ET,mm h-1)for the T.ramosissima stand from June 20 to June 23,2012
Data analysis
Basic statistical features of mean values and SD for meteorological variables were analyzed and significant differences tested using one-way ANOVA atP=0.05.Meteorological data collected from the site were used,includingRn,Ta,RH,U,andVPD,to explain seasonalETvariability by ANOVA as well as correlation and regression analyses provided in the SPSS Statistics 19 software package(IBM,USA).Finally,Origin 8.0 software was used to generate drawings(Origin Lab Corp.,Northampton,MA,USA).
Results
Comparisons of environmental variables
Limited precipitation,lowRHas well as highTaandVPDwere the main climatic variables used for the study area.Precipitation was 32.7 mm in 2012 and 17.2 mm in 2014.Two extreme precipitation events,8.3 and 8.9 mm on June 6 and July 24,respectively,could account for the high precipitation in 2012.AtP=0.05,population means ofN,Ta,VPD,andUwere not significantly different for either study year with the exception ofRHfrom the meteorological station(Table 1).
For theT.ramosissimastand,dailyRnincreased during the spring and progressed into the summer before progressively decreasing during the autumn from 3.0 to 21.0 MJ m-2day-1with an average of 12.4 MJ m-2day-1.Moreover,Gwas coincident withRn(Fig.4a).DailyTaremained positive at an average of 18.5°C throughout the growing seasons,particularly in August when maximum dailyTareached 31.6°C.Seasonal variation inRHchanged followingRnandTawith an average of 31.3%(Fig.4b)and was particularly low from March to May compared to other months(20.2 vs.35.9%,P=0.01),which may be attributable to the highUobserved during this period(2.1 vs.1.4 m s-1,P=0.05)(Fig.4c).MeanVPDwas 1.8 kPa,and seasonal variation inVPDwas closely correlated toRnandTa(Fig.4c).
For theP.euphraticastand,Rnwas significantly higher with an average of 14.1 MJ m-2day-1compared to theT.ramosissimastand(P=0.01)during the growing seasons,andGcoincided withRn(Fig.4d).TaandRH,with an average of 19.0°C and 29.1%,was not significantly different from theT.ramosissimastand(P=0.50 andP=0.07,respectively)(Fig.4e).UndifferentiatedTaandRHresulting fromVPDwas also not significantly different(1.8 kPa,P=0.75)to theT.ramosissimastand(Fig.4f).Height(20 m)of measuredUresults was significantly higher(3.4 vs.1.6 m s-1,P=0.01)compared to theT.ramosissimastand.
Comparison of evapotranspiration between the two stands
Energy balance closures(LE+H)/(Rn-G)were 79 and 78%in 2012 and 2014,respectively(Fig.2).During the dormantseason,dailyETwas generally less than 0.5 mm day-1.It is noteworthy thatETremained weak throughout March and April butET0remained relatively high(Fig.5a),from whichTa,VPD,andUincreased rapidly(Fig.4b,c).For the growing season,dailyETincreased during the spring and progressed into the summer before progressively decreasing in the autumn from 0.3 to 8.0 mm day-1with an average of 3.6 mm day-1(Fig.5a).However,there was pronounced lowerETvalue in June that was consistent with the lowerUfor the same period,although it rose sharply afterwards in conjunction with the highVPDin August with a maximum of 7.7 mm day-1.In contrast,ET0was high during the late spring and autumn,indicating a lowETtoET0ratio(Kc:from 0.09 to 0.22)during these periods.In comparison,Kcremained high(0.36–0.86)during the summer and early autumn with an average of 0.46 in 2012.This phenomenon indicated that soil evaporation was weak because plant transpiration could not have occurred in March and April.MonthlyETincreased from 34.1 mm in May to 180.1 mm in July and decreased to 93.1 mm in September for a total of 548 mm in 2012(Table 3).
Fig.4 Daily variation in meteorological variables,including net radiation(Rn,W m-2),soil heat flux(G,W m-2),air temperature(Ta,°C),relative humidity(RH,%),vapor pressure deficit(VPD,
For theP.euphraticastand,ETincreased earlier and ended later than for theT.ramosissimastand,after which it was consistent withET0(Fig.5b).DailyETwas less than 1 mm prior to March(Ta<0 °C)during the dormant season(Fig.5b).However,ETincreased rapidly with a peak of 3 mm day-1after March,at which timeTa,VPDandUincreased rapidly(Fig.4e and f),andET0was relatively high(Fig.5b).During the growing season,dailyETranged from 0.9 to 7.9 mm with an average of 4.6 mm day-1(Fig.5b),and monthlyETincreased from 131.3 mm in May to 170.0 mm in July and decreased to 98.5 mm in September for a total of 707 mm in 2014(Table 3).kPa),and wind speed(U,m s-1)forT.ramosissima(a–c)andP.euphratica(d–f)during 2012 and 2014 growing seasons
Comparison of variables associated with evapotranspiration
For theT.ramosissimastand,dailyETwas positive and significantly correlated toRn,Ta,andVPDand negative and significantly correlated toRH,but not significantly correlated toU.Due to the high correlations betweenVPDandRH,the latter was removed during stepwise regression analysis,for which the results revealed that climate variables accounted for at least 81%ofETvariation.For theP.euphraticastand,dailyETalso positive and significantly correlated toRn,Ta,andVPDand negative and significantly correlated toRH(Ubeing the exception),and climate factors,includingRn,Ta,andVPD,accounted for at least 80%ofETvariation(Table 4).
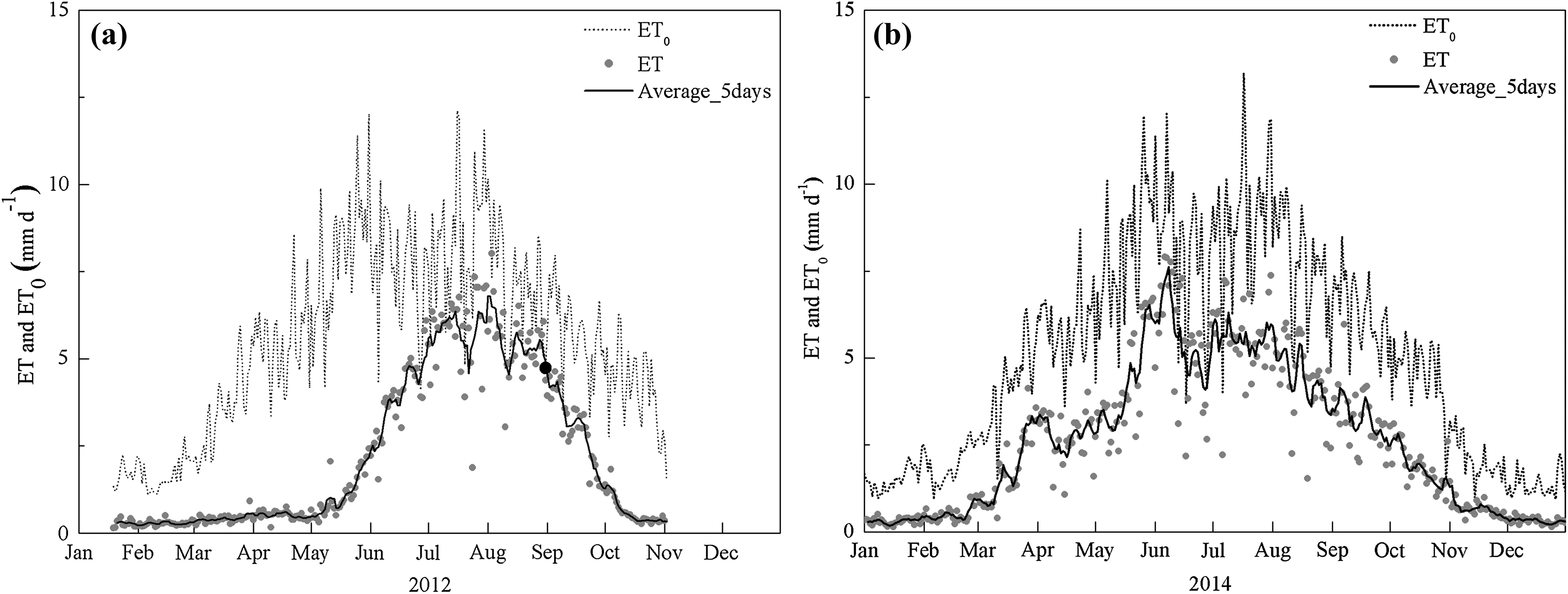
Fig.5 Seasonal patterns in daily reference evapotranspiration(ET0,mm day-1)and evapotranspiration(ET,mm day-1)for T.ramosissima(a)and P.euphratica(b)in 2012 and 2014.The 5-day average value of ET(black solid line)is shown
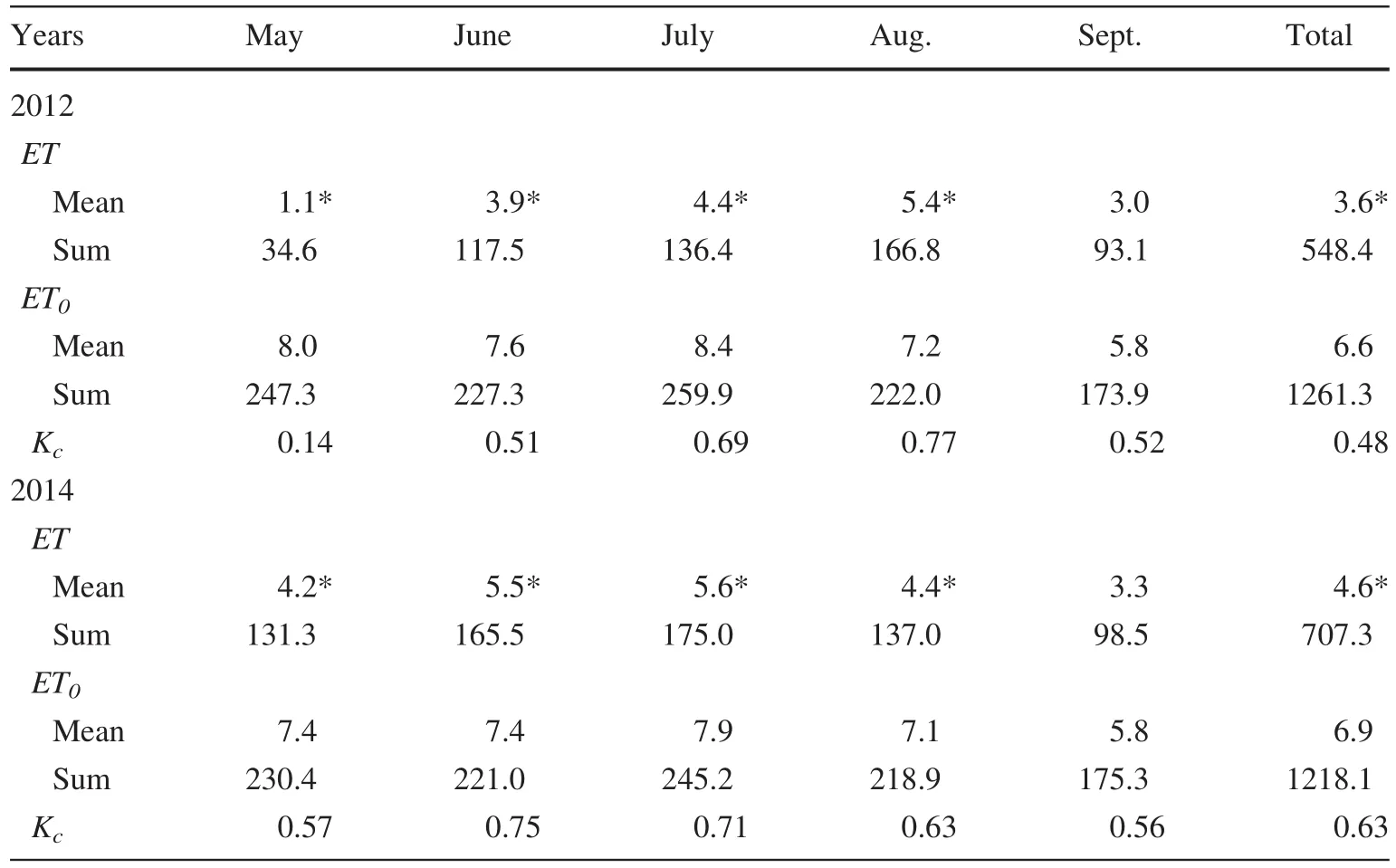
Table 3 Monthly variation in evapotranspiration(ET,mm),reference evapotranspiration(ET0,mm),and crop coefficient(Kc=ET/ET0)of T.ramosissima and P.euphratica during the 2012 and 2014 growing seasons
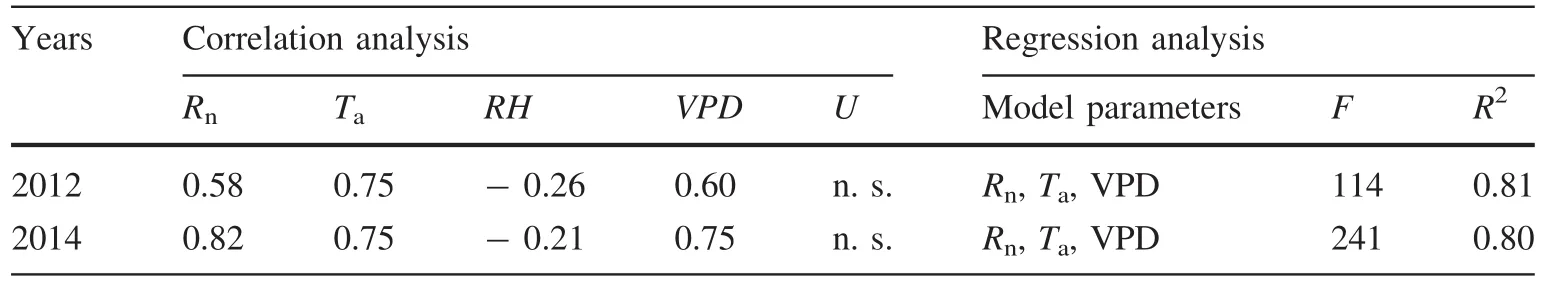
Table 4 Statistical summary of evapotranspiration patterns(dependent variables)to climate parameters for T.ramosissima and P.euphratica during the 2012 and 2014 growing seasons
Discussion
Comparison of evapotranspiration between the two stands
Energy balance closures for the two stands(Fig.2)were comparable with FLUXNET sites(Wilson et al.2002)as well as with other EC measurements, especially in the arid region of northwestern China(Hao et al.2007;Yuan et al.2014).This suggests that directETmeasurements using the EC method were reliable.The totalETofT.ramosissimaandP.euphraticaduring the growing season has been estimated at tree-level(Si et al.2007;Zhang et al.2006)and stand-level scales(Houet al.2010;Siet al.2005;Wanget al.2014).The BREB estimatedETwas approximately248 mm with an average of1.6 mm day-1forT.ramosissima(Siet al.2005)and447 mm with an average of3.2 mm day-1for theP.euphraticastand(Houet al.2010).In addition,annual transpiration measured through sap flow was 214.9 mm forP.euphratica(Si et al.2007).The daily ET rate during the summer months(June–August)was estimated to be0.63–0.73 mmforT.ramosissimaand1.89–2.33 mm forP.euphraticausing the water table fluctuation(WTF)method(Wanget al.2014).A comparison of results from this study and those of the aforementioned studies indicate that theETrates estimated using the EC method for both stands were higher than values determined through sap flow,BREB,and WTF methods.
Previous studies(Cleverly et al.2006;Devitt et al.2011)have shown that climate factors,vegetation parameters,soil properties,and water table depths are principal factors controllingETvariation in phreatophytic communities.Climate variables,soil types,plant community composition,and water table depth for the two stands were similar for both years(Tables 1,2).Thus,their differentETvalues were likely caused by differing soil moisture levels induced by irrigation and further different soil evaporative conditions(Fig.5).Before 2013,theP.euphraticastand had no access to irrigation and groundwater depth may have been 4 m.After 2013,the irrigation was applied in autumn.Thus,we deduced that the significantly higherETfound in theP.euphraticastand was directly related to the high soil evaporation rate at this site.Reasons for this are discussed below.
Comparison of evapotranspiration between the study sites to other regions
Due to its wide distribution,ETfromTamarixspecies,an introduced,stress-tolerant shrub from Central Asia,has been extensively investigated throughout the world.TheETresults from theT.ramosissimastand using EC were similar to results in the lower Tarim River basin,China,with a meanETof 500 mm a-1and an averageETof 3.85 mm day-1(Yuan et al.2014).A comparison of results obtained from this study and one in the lower Tarim River(Yuan et al.2014)showed thatT.ramosissimagrowth in both did not experience water stress.Thus,results from both studies may represent potentialETfor nativeT.ramosissimastands in Central Asia.It is noteworthy that the slightly higherETfound in our study(compared to the results of Yuan et al.2014)may be attributed to the relatively higher LAI(1.97 vs.1.15,respectively)and the lower groundwater depth(2.65 vs.6.62 m,respectively).
However,totalETobtained from EC flux tower measurements(Cleverly et al.2002,2006;Devitt et al.1998;Westenburg et al.2006)in combination with remote sensing data(Bawazir et al.2009;Nagler et al.2005,2009;Taghvaeian et al.2014)yielded a broad range from 800 to 1450 mm year-1in the western United States.It may therefore be concluded that observedETunder arid conditions in native regions was clearly low compared to invasiveT.ramosissimastands in non-native regions.These differences could be attributable to two specific reasons.First,soil evaporation inT.ramosissimastands in arid regions could be either weak or negligible(Yuan et al.2014).Second,although LAI can have a deterministic impact on totalET(Dahm et al.2002;Sala et al.1996;Yuan et al.2014),values from the study and those of Yuan et al.(2014)were lower than those in the middle Rio Grande district(2.5 and 3.5 for flooding and non- flooding,respectively)(Cleverly et al.2006;Nagler et al.2005)and the lower Colorado River(2.54)(Nagler et al.2009),both in the western U.S.
Recent studies have shown that water use by individualT.ramosissimatrees is comparable to the native species that it typically replaces such as cottonwood and willow(Glenn and Nagler 2005;Nagler et al.2003).However,in these cases tree-level soil evaporation had not been taken into account.Although soil evaporation was weak for theT.ramosissimastand as reported by Yuan et al.(2014),this was not the case for theP.euphraticastand due to the highETobserved during the dormant season(Fig.5b).Based on the simplified Shuttleworth–Wallace(SSW)model,the ratio of canopy transpiration toETranged from 0.53 to 0.66(Gao et al.2016a).Thus,we estimated that transpiration was 465 mm for theP.euphraticastand which was comparable to 473 mm for theT.ramosissimastand when soil evaporation was disregarded(a total of 77 mm a-1with an average of 0.5 mm day-1determined by measuringETduring the dormant season in conjunction with results from Yuan et al.(2014).For greater accuracy,further research is required to partitionETinto plant transpiration and soil evaporation.
Factors and variables associated with evapotranspiration
ETrates responded toGD,atmospheric surface layer conditions,stomatal coupling in conjunction with atmospheric humidity levels,energy balances,and LAI(Cleverly et al.2006).In desert riparian regions,given the limited rainfall,groundwater provides an ample water supply to riparian forests(Si et al.2014),permitting greater annualETrates than that provided by rainfall alone(Table 3).Some studies have shown that growth and ecophysiological parameters of riparian forests were less severely affected byGDwithin a depth of 6 m in arid regions in China(Chen et al.2006;Feng et al.2012;Hao et al.2009).MeasuredETofT.ramosissimaandP.euphraticastands using the EC method in the lower HRB was comparable to results in the Tarim River basin(Yuan et al.2014).In contrast,the clear differences inETin native and invasive regions indicate that climate factors and water availability,which both impact plant eco-physiological characteristics,were the primary controllers of water use for these species.
Regression analysis demonstrate that climate and soil water variables accounting for at least 80%ofETvariation in the two riparian forests investigated(Table 4).In addition,plant eco-physiological characteristics,particularly LAI,determined spatial patterns ofETfor riparian phreatophytes.Moreover,LAI in combination with plant physiological parameters such as stomatal conductance,would impact seasonal variation inET(Hatler and Hart 2009).Desert riparian species adapted to arid climates may use of deep soil water and groundwater at the same time.In contrast,greater amounts of rainfall and flooding may supply sufficient available water in shallow soil layers of the arid southwestern U.S.This suggests that plant water use is the result of long-term adaption to local climates and water availability.
Conclusions
During the growing season,ETof theT.ramosissimastand was 548 mm a-1with a mean of 3.6 mm day-1,similar to that of native forests in Central Asia(500 mm a-1)but clearly lower than that of invasive regions in the western U.S.(from 800 to 1450 mm a-1).ETof theP.euphraticastand was 707 mm with a mean of 4.6 mm day-1,which was directly linked to high soil evaporation rates under sufficient water availability induced by irrigation.When soil evaporation was disregarded,total transpiration was 473 and 465 mm for theP.euphraticaandT.ramosissimastands,respectively,indicating comparable water use of the two contrasting riparian forests.Furthermore,regression analysis demonstrated that climate factors accounted for at least 80%ofETvariation for both stands.Thus,our results show that plant water use is the result of long-term adaptation to local climates and water availability.
AcknowledgementsThe authors would like to thank two anonymous reviewers for their constructive and valuable assessment and comments,which helped to improve this article.
 Journal of Forestry Research2018年5期
Journal of Forestry Research2018年5期
- Journal of Forestry Research的其它文章
- Environmental load of solid wood floor production from larch grown at different planting densities based on a life cycle assessment
- An overview of proven Climate Change Vulnerability Assessment tools for forests and forest-dependent communities across the globe:a literature analysis
- Assessing the vulnerability of a forest ecosystem to climate change and variability in the western Mediterranean sub-region of Turkey:future evaluation
- Effects of management regimes on carbon sequestration under the Natural Forest Protection Program in northeast China
- Mixed-effects modeling for tree height prediction models of Oriental beech in the Hyrcanian forests
- Estimation of a basal area growth model for individual trees in uneven-aged Caspian mixed species forests