
不同陆地棉品种花芽分化与茎尖内源激素的关系
2018-09-07 08:54张金宝吴国峰陈宗全王晓玲谭阳光王清连
江苏农业科学 2018年16期
马 亮, 李 飞, 张金宝, 吴国峰, 陈宗全, 王晓玲, 谭阳光, 王清连
(现代生物育种河南省协同创新中心/河南科技学院棉花研究所,河南新乡 453003)
陆地棉(GossypiumhirsutumL.)是世界四大棉花栽培种之一,于19世纪末期传入我国。目前,除新疆有种植少量海岛棉外,我国大部分产棉区均种植陆地棉材料。花芽分化是植物由营养生长向生殖生长转变的重要生理和形态标志,对花芽分化的研究始于1877年,Askenasy对酸樱桃花芽分化作了相关报道[1]。棉花的花芽是形成棉铃和纤维产量的基础[2],花芽分化的迟早影响到结铃是否适期,花芽分化的多少关系到棉纤维产量的高低[3],同时花芽分化与早熟也密切相关[4],因此研究陆地棉花芽分化具有极其重要的意义。孙济中等指出,陆地棉花芽分化按照各花器官原基向心分化的自然顺序进行,并将其划分为花原基伸长期、苞片原基分化期、花萼原基分化期、花瓣原基分化期、雄蕊原基分化期和心皮分化期等6个时期[5]。此外,植物花芽分化是一个重要且复杂的形态建成过程,由内外因子相互协调共同完成。马焕普指出,在该过程中营养是花芽分化的基础,内源激素调节是花芽分化的关键,基因表达是花芽分化的途径[6]。近年来,对花卉和果树花芽分化的研究表明,高浓度的玉米素核素(ZR)能促进花芽分化,而生长素(IAA)、脱落酸(ABA)和赤霉素(GA3)对花芽分化的作用尚不统一[7-9]。本试验试图利用扫描电镜观察不同陆地棉花芽的形态分化,并在此基础上分析上述4种内源激素含量及比例的变化,以期了解花芽分化的生理学基础,为陆地棉花期的化学调控和品种最适移栽期研究提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为不同熟性的4个陆地棉品种(百棉1号、中棉所41、鲁棉研21号和DP99B),种子均经过多代自交,经硫酸脱绒后晒干,选择成熟饱满的籽粒,2014年5月1日种植于河南科技学院辉县棉花育种试验田,单行区,行长5 m,行距 1 m,每行15~17株,每个材料种植15行,按常规大田管理。
1.2 取样与保存
供试材料取样时期为1~8叶期,每个叶期分别取幼苗置于福尔马林-乙酸-乙醇(formalin-aceto-alcohol,简称FAA)固定液中固定保存;另取茎尖若干,经液氮罐速冻后,置于-80 ℃冰箱内保存待测。
1.3 解剖学观察
对于保存于FAA固定液中的幼苗,切下其茎尖,在体视镜下解剖,去除幼叶、较大叶原基和托叶等组织,露出生长点;然后用不同浓度的乙醇梯度脱水,叔丁醇逐步替换,4 ℃放置12 h;在真空干燥箱(BPZ-6033)中干燥后,粘贴于粘有双面胶的铝制样品台上,在扫描电镜(Quanta 200)下使用15 kV电压观察拍照[10]。此外,幼苗各叶位的腋芽在体视镜下解剖、观察和记录。
1.4 内源激素含量的测定
在解剖学的基础上,称取供试材料的每个叶期0.2 g左右的样品(保存于冰箱内的茎尖),称取3次。内源激素的提取、纯化和含量的测定均由中国农业大学农学与生物技术学院完成。其中,选用80%甲醇[内含1 mmol/L 二丁基羟基甲苯(butylated hydroxytoluene,简称BHT)]作为提取液,C18固相萃取柱用于纯化激素,采用酶联免疫法(ELISA)[11-12]进行内源激素含量(鲜质量)的测定,试剂盒由中国农业大学提供,测定激素为IAA、ABA、GA3和ZR,每个样品重复3次。
1.5 数据处理
采用Excel 2010和SPSS 19.0进行数据整理和分析。
2 结果与分析
2.1 陆地棉花芽分化的解剖学观察
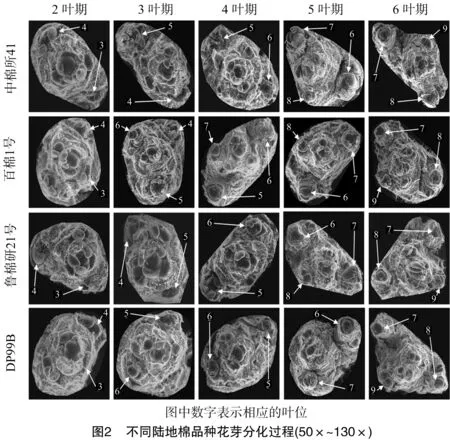
参照孙济中等对陆地棉花芽分化时期的划分[5],对陆地棉品种百棉1号的茎尖顶芽和侧芽进行精细解剖。花原基伸长期(图1-a):腋芽生长锥伸长呈圆柱形突起;苞片原基分化期(图1-b、图1-c、图1-d):花原基先分化出1片边缘光滑的半椭圆形苞片原基,随后分化出第2和第3片苞片原基,同时随着苞片原基的增大,边缘出现锯状苞齿;花萼原基分化期(图1-e、图1-f):苞片原基内部形成5片环状花萼原基,基部相连,顶部凸起;花瓣原基分化期(图1-g、图1-h):花萼原基内部组织继续分化,中间凹陷,周围出现1圈隆起,随后圈状隆起边缘形成5个突起的花瓣原基。在陆地棉花芽分化的整个过程中,苞片原基分化期通常作为花芽形态分化开始的标志,又称为形态分化初期。

由图2可知,4个品种花芽形态分化呈现出较大差异。百棉1号一般在3叶期即可观察到5~6叶位已进入苞片分化期;而中棉所41在5叶期才能观察到6~8叶位进入苞片分化期;鲁棉研21号和DP99B 2个品种的花芽初始分化发生在6~7叶位,在4叶期可明显观察到分化。此外,在6叶期,百棉1号、鲁棉研21号和DP99B 3个品种的第7叶位已进入花瓣原基分化期,而中棉所41仍处于花萼原基分化期。值得指出的是,百棉1号、鲁棉研21号、DP99B、中棉所41分别于3叶期、4叶期、4叶期、5叶期进入花芽形态分化初期。
2.2 陆地棉花芽分化期间茎尖内源激素含量的变化
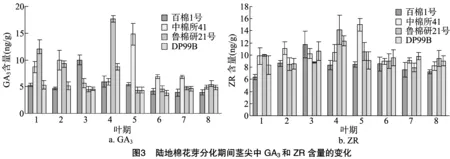
2.2.1 GA3含量的变化 由图3-a可知,在陆地棉花芽分化过程中的8个叶期,4个品种的GA3含量变化具有明显差异。百棉1号在2叶期时,GA3含量略有降低,在3叶期出现最高峰,随后逐渐降低;中棉所41在2叶期时,GA3含量稍有上升,随后降低,在4叶期回升,在5叶期时出现最高峰,然后逐渐降低;鲁棉研21号在1叶期时GA3含量较高,随后降低,在3叶期出现低谷,紧接着4叶期迅速达到最高峰,然后5叶期降低至接近3叶期的水平,随后缓慢上升;DP99B在3叶期时,GA3含量略有降低,4叶期出现最高峰,随后降低,至6叶期出现低谷,然后缓慢上升。
2.2.2 ZR含量的变化 由图3-b可知,4个陆地棉品种在8个叶期ZR含量的变化情况有所不同。百棉1号在3叶期时,ZR含量达到最大值,为11.8 ng/g,4叶期降低,随后缓慢上升,6~8叶期缓慢降低;中棉所41在2叶期时ZR含量略有上升,3叶期稍有降低,随后上升,至5叶期时出现最高峰,6叶期急速下降,7叶期稍有上升,8叶期略有降低;鲁棉研21号在2叶期时ZR含量略有降低,随后上升,4叶期具有最大值,为14.2 ng/g,接着下降,至7叶期出现最小值,在8叶期稍有上升;DP99B的ZR含量在1~4叶期逐渐上升,4叶期具有最大值,为12.3 ng/g,5叶期降低,随后缓慢上升,8叶期略有降低。
2.2.3 ABA含量的变化 由图4-a可知,4个陆地棉品种的ABA含量在花芽分化过程中呈现出相似的变化趋势,即先逐渐上升,达到最高峰后再逐步下降。不同的是百棉1号在3叶期达到最高峰,中棉所41达到高峰时间为5叶期,而鲁棉研21号和DP99B为4叶期。
2.2.4 IAA含量的变化 由图4-b可知,花芽分化过程中4个陆地棉品种IAA含量的变化趋势略有差异。百棉1号的IAA含量变化呈现出上升—下降—上升的趋势,在3叶期出现最低谷;中棉所41和鲁棉研21号均呈现出先下降再上升的趋势,分别在5叶期和4叶期出现最低谷;DP99B呈现出下降—上升—下降—上升的趋势,在4叶期出现最低谷。

2.3 花芽分化期间茎尖内源激素比值的变化
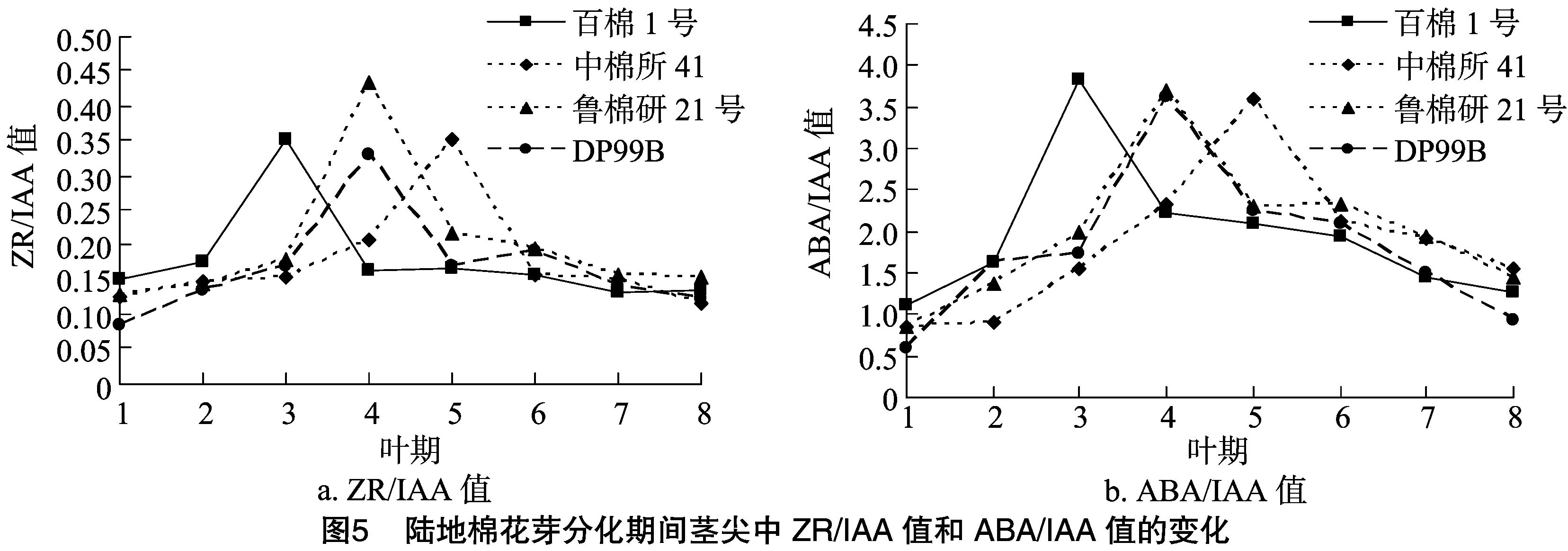
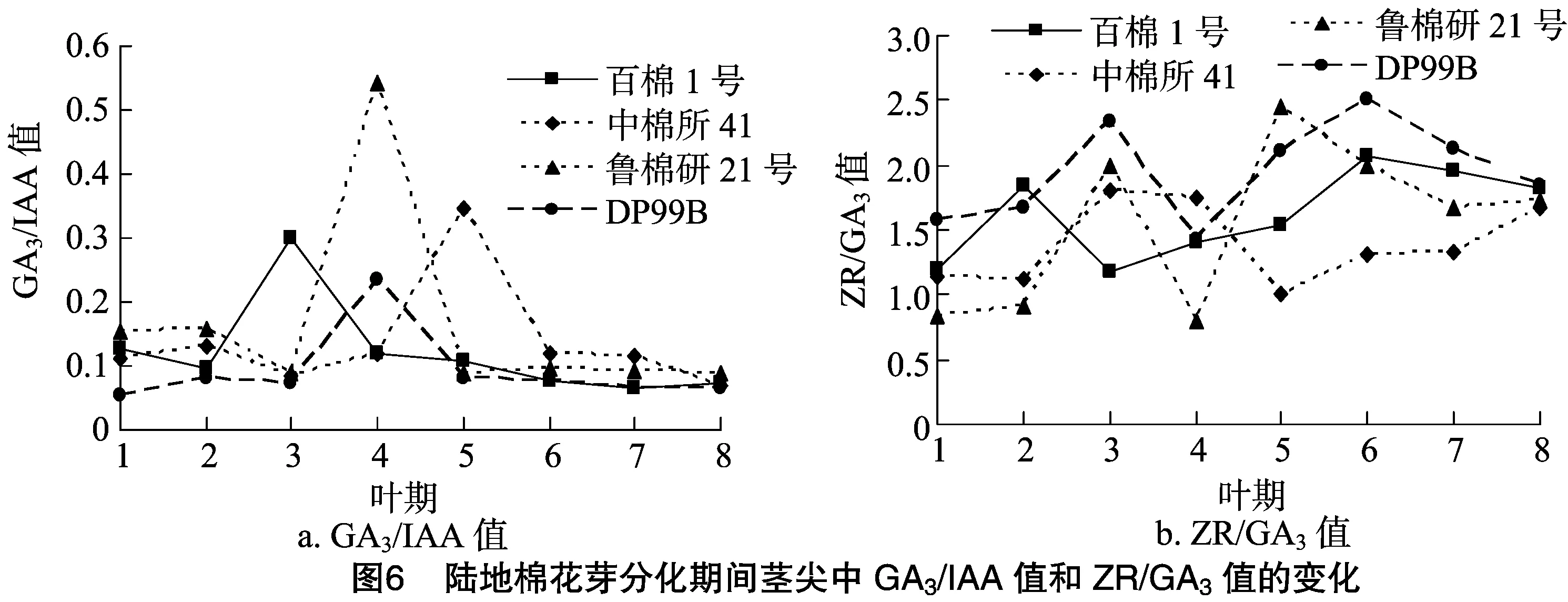
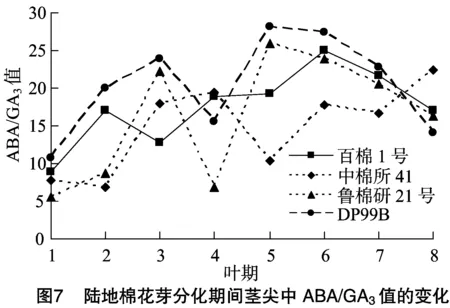
由图5、图6、图7可知,花芽分化期间4个陆地棉品种的内源激素比值具有规律性变化。ZR/IAA、ABA/IAA和 GA3/IAA 值变化趋势相似,整体上呈先逐步上升后急速下降最终趋于平缓的趋势。其中,百棉1号的3个比值在3叶期达到最大值;中棉所41的3个比值在5叶期达到最大值;鲁棉研21号和DP99B的3个比值在4叶期达到最大值。此外,鲁棉研21号的ZR/IAA、GA3/IAA值在4个品种内具有最高峰,分别为0.43、0.54;百棉1号的ABA/IAA值具有最高峰,为3.83。ZR/GA3和ABA/GA3值整体上均呈“M”形变化趋势,百棉1号、中棉所41、鲁棉研21号和DP99B 4个品种的 ZR/GA3值分别于3叶期、5叶期、4叶期和4叶期出现低谷。其中,鲁棉研21号的变化趋势最为明显。
3 结论与讨论
长期以来,我国粮棉争地问题严重,陆地棉营养钵育苗是提高复种指数、实现粮棉双丰收的有效措施。同时,与传统种植方式相比,营养钵育苗在产量和质量方面均有所提高。花芽分化作为植物由营养生长向生殖生长转变的标志,对育苗最适移栽期的选择具有重要的指导意义。百棉1号在3叶期开始花芽分化,其最适移栽期即为3叶期。同样,中棉所41的最适移栽期为5叶期,鲁棉研21号和DP99B的最适移栽期为4叶期。
3.1 内源激素与花芽分化的关系
赤霉素对种子萌发和植物生长具有促进作用。目前,GA3作为一种抑花激素,在苹果[13]、平阴玫瑰[14]、油橄榄[15]、芒果[16]、大樱桃[17]、光皮树[18]等植物中已有报道。但也有研究表明,GA3可以促进植物花芽分化。程华等研究发现,GA3有利于板栗花序原基的形成,且花序原基分化期和花簇原基分化期均需要维持较高的GA3浓度[9];在大葱花芽分化期间,根、假茎和叶片中GA3含量逐渐上升,出现峰值的时间与完成花芽分化时期相吻合[19]。此外,GA3对花芽分化的作用可能与其类型有关。Ulger等指出,GA3浓度过高会抑制橄榄的花芽分化,然而高浓度的GA3对橄榄的成花有促进作用[20]。本研究发现,4个陆地棉品种的GA3含量均在花芽形态分化初期达到最大值,说明高浓度的GA3能促进陆地棉花芽由营养生长转向生殖生长。
ZR作为细胞分裂素的一种,参与许多植物的成花转变过程。李秉真等分别在对梨和柑橘的研究中发现,在花芽分化的各个时期,花芽中的ZR含量均高于叶芽[21-22]。油桐在花芽形态分化期,ZR含量呈现前期上升、中期波动、末期上升的变化情况[23]。在本研究中,陆地棉的ZR含量在形态分化初期出现高峰,进入形态分化期后开始下降,说明ZR对陆地棉花芽分化具有促进作用。这与胡绍庆等关于桂花的研究结果[24]一致。
生长抑制剂ABA对花芽分化的作用说法不一。多数人认为ABA是一种重要的促花激素[25-26]。郭蕊等对百合内源激素与鳞茎休眠关系的研究发现,ABA可以打破休眠,是花芽开始分化的关键物质[27]。Okuda等通过研究温州蜜橘认为,ABA可以同GA3拮抗,促进花芽孕育[28]。本研究发现,ABA在陆地棉花芽未分化时含量较低,随着生长发育的进行,ABA含量逐渐增高,到花芽形态分化初期达到最高峰,表明高浓度的ABA有利于陆地棉花芽分化。
IAA与植物成花的关系一直存在争议。彭桂群等研究表明,IAA是一种抑花激素,能抑制平阴玫瑰的花芽分化[14];而孔德政等对碗莲的研究发现,较高浓度的IAA对碗莲花芽分化起促进作用[29]。目前普遍认为,低浓度的IAA是花芽分化所必需的,但高浓度IAA能抑制开花。本研究中,4个陆地棉品种的IAA含量在花芽形态分化初期出现低谷,随后有所升高,但升高幅度不大,说明低浓度的IAA有利于陆地棉花芽分化。
3.2 激素平衡与花芽分化的关系
虽然各类激素在陆地棉花芽分化过程中呈现出一定的变化规律,但植物成花并不是由单一的某种激素决定的,往往还依赖于激素之间的平衡。各种激素在时间和空间上相互作用、相互促进、相互拮抗而形成一种动态平衡,这种平衡是花芽分化能够顺利进行的物质基础[30-31]。分析陆地棉花芽分化与激素平衡的关系表明,ZR/IAA、ABA/IAA和GA3/IAA值在陆地棉花芽分化期间呈现先上升后下降的过程,在花芽形态分化初期达到最大值。说明高水平的ZR/IAA、ABA/IAA和GA3/IAA值有利于陆地棉花芽分化,这与程华等对板栗的研究结果[9]一致。ZR/GA3和ABA/GA3值在花芽形态分化初期出现低谷,说明低水平的ZR/GA3和ABA/GA3值能促进棉花由营养生长向生殖生长转变。
总之,内源激素和激素平衡对陆地棉花芽分化起着十分重要的作用,任何一种激素含量的变化都能影响花芽分化,因此可以借助化学调控外施相应的内源激素使花芽分化提前或者推迟。值得注意的是,不同熟性的陆地棉品种花芽分化的情况具有一定的差异,在花芽分化起始阶段各类内源激素的含量也不尽相同,如4个供试品种中虽然百棉1号花芽分化最早,但是花芽形态分化初期GA3含量却并非最高。此外,核酸、蛋白质、碳水化合物等也能影响陆地棉花芽分化,具体作用还有待进一步研究探讨。
猜你喜欢
上海农业科技(2022年6期)2022-12-14
新疆农业科学(2020年4期)2020-04-07
浙江农业科学(2019年1期)2019-01-28
计算机应用(2018年10期)2018-11-22
中国棉花(2018年3期)2018-04-09
今日农业(2017年1期)2017-12-22
社会科学研究(2017年5期)2017-11-01
华南师范大学学报(自然科学版)(2017年4期)2017-09-11
浙江大学学报(农业与生命科学版)(2017年2期)2017-05-19
杂草学报(2014年1期)2014-08-13
