
Polyphasic examination on Merismopedia tenuissima CHAB 7021 from Ganjiang River, China revealed the polyphyly of the genus Merismopedia (Cyanobacteria)*
2018-08-02 02:50:42SHENLiqin沈丽琴MASuchao马苏超CAIFangfang蔡芳芳YUGongliang虞功亮LIShouchun李守淳LIRenhui李仁辉
关键词:马苏
SHEN Liqin (沈丽琴) , MA Suchao (马苏超) , CAI Fangfang (蔡芳芳) ,YU Gongliang (虞功亮) LI Shouchun (李守淳) , , LI Renhui(李仁辉)
1 College of Life Science, Jiangxi Normal University, Nanchang 330022, China
2 Key Lab of Algal Biology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China
3 University of Chinese Academy of Sciences, Beijing 100049, China
Abstract Species in the cyanobacterial genus Merismopedia are present in freshwaters at different trophic levels, with some species even as the components of cyanobacterial blooms. However, species diversity in this genus was not fully veri fied by molecular investigation and polyphasic taxonomic studies.In this study, Merismopedia-like strain tenuissima CHAB 7021 was isolated from Ganjiang River in Jiangxi Province, China, and polyphasic characterization of this strain was performed by morphological observation,ultrastructural examination, chemical detection of pigments and phylogenetic analysis based on 16S rRNA gene sequences. Morphological identi fication of the strain was supported by the ultrastructural features,as the tiny species Merismopedia tenuissima Lemmermann. The phylogeny based on 16S rRNA gene sequences revealed at least three clades formed by the strains of Merismopedia. The three M. tenuissima strains including M. tenuissima CHAB 7021 was gathered in clade III with distant relationship to the clade I formed by the six Merismopedia strains including the type species M. punctata, and such a genetic distance may propose Merismopedia tenuissima to separate from Merismopedia genus. However intermixture relationship in between strains of M. punctate and M. glauca in the phylogenetic tree still complicated the taxonomic status in the genus Merismopedia. The process for taxonomic revision in the Merismopedia genus still await for examination and further information on more strains of type species M. punctata.
Keyword: cyanobacteria; Merismopedia; polyphasic; phylogeny; taxonomic revision
1 INTRODUCTION
Cyanobacteria are the most diverse group of prokaryotes, morphologically ranging from simple unicells, colonies of coccid cells and simple filaments without differentiated cells to filaments with differentiated cells and branches (Castenholz et al.,2001). Cyanobacterial organisms had been traditionally classi fied based on morphological and ecological characters, but such a taxonomic system has been continuously revised since the advent of DNA sequence information. The polyphasic approach,mainly using DNA sequences in addition to morphology and habitat preference, has brought a resolution to the cyanobacterial taxonomic problem,therefore this approach has been commonly used in characterizing cyanobacterial taxa and taxonomic revision. Along the revision process in cyanobacterial taxonomy, filamentous cyanobacteria, generally including oscillatorean and nostocalean groups, have largely led to a considerable change. In contrast,chroococcaceaen cyanobacteria, represented as unicellular or unicellular colonial group, have been less revised, especially at both genus and species levels.
The genus Merismopedia was established by Meyen in 1939 with Merismopedia punctata as type species, and the genus is characterized as a coccoid,unicellular cyanobacterium forming flat cell plates of regularly arranged cells by synchronous division in two planes (Geitler, 1932; Meyen, 1839; Komárek and Anagnostidis, 1986). Species in the genus Merismopedia are present in a range of environments covering both oligotrophic lakes with low alkalinity and eutrophic water reservoirs found in Brazilian semi-arid region (Hörnström, 1999; Dantas et al.,2011), with some species even as the components of cyanobacterial blooms. Species level classi fication within the genus Merismopedia was mainly based on morphological characters differing in the size of individual cells, in the maximal number of cells within aggregates, and in the size of cell aggregates(Geitler, 1932; Desikachary, 1959; Kaas et al., 1985).There are currently 40 taxonomically accepted species in the genus Merismopedia, based on the description in Algaebase (Guiry and Guiry, 2018), and such a species number re flects a relatively higher diversity in the genus. However this species diversity was not fully veri fied by molecular investigation and polyphasic taxonomic studies. Palinska et al. (1996)investigated six Merismopedia strains assigned into three species as M. punctata, M. glauca, and M.elegans, based on polyphasic characterization, and they found that these differently phenotypic strains had the identical genotypes based on DNA sequences of 16S rRNA gene. This finding seemed to us that the genus Merismopedia may represent a monophylic group. There had been no more DNA sequences available from Merismopedia species for quite a long time. Castiglioni et al. (2004) deposited a 16S rRNA gene sequence of Merismopedia tenuissima 0BB46S01 in Genbank, but did not give any characteristic description on this strain. Rajaniemi-Wacklin et al. (2006) reported that M. glauca strains 0BB39S01 isolated from Bubano Basin, Italy, and this strain was rather distantly related to the Merismopedia strains published by Palinska et al.(1996) (93.5% 16S rRNA gene sequence similarity).All the Merismopedia strain did not form a monophyletic cluster in any of the phylogenetic trees,indicating that these Merismopedia strains are genetically more heterogeneous and might even belong to different genera. Furtado et al. (2009)reported Merismopedia sp. CENA106 from a Brazilian facultative wastewater stabilization pond based one morphological and molecular characterization, and this strain was shown to have a high homology in 16S rDNA sequence to Merismopedia tenuissima 0BB46S01, but both Merismopedia sp. CENA106 and Merismopedia tenuissima 0BB46S01 were not forming a close cluster with any Merismopedia strains in the studies of Palinska et al. (1996) and Rajaniemi-Wacklin et al.(2006). However, Thomazeau et al. (2010) con firmed the monophyly of the Merismopedia genus based on phylogeny formed by the 16S rDNA sequences of 144 cyanobacterial strains including only three Merismopedia. Therefore previous studies on diversity, taxonomy and phylogeny on the genus Merismopedia brought quite con flicting results,indicating currently confused status on taxonomy within the genus of Merismopedia. To solve this problem, polyphasic characterization on more Merismopedia strains from more regions should be performed.
In China, 11 species in the genus Merismopedia were described in 1991 (Zhu, 1991) and no new species have been described since then and no taxonomic studies based cultivated strains have been performed. In the present study, a Merismopedia-like strain was isolated from Ganjiang River in Jiangxi Province, China. The strain was taxonomically and phylogenetically evaluated by investigating several characteristics, including morphological features,cellular ultrastructures, pigment composition, and 16S rRNA gene sequences. The results of this study provided insights into the taxonomy and diversity and phylogeny of Merismopedia.
2 MATERIAL AND METHOD
2.1 Strain isolation and culture
A strain was isolated by using classical micropipette method (Rippka, 1988) from the sample collected in the middle reach of Ganjiang River in Jiangxi Province, China, in December, 2016. The unialgal strain, named as Merismopedia-like tenuissima strain CHAB 7021 was cultured in liquid CT medium(Ichimura, 1979) with constant white light intensity of 30 μmol/(m2·s) under a 12 h:12 h L/D cycle at 25±1°C. This strain was stored in the Harmful Algae Biology Laboratory, Institute of Hydrobiology,Chinese Academy of Sciences.
2.2 Morphological analysis
Morphological observation by using Nikon eclipse 80i light microscope with the external DS-Ril digital camera (Nikon, Japan) connected to the desktop computer through the converter. Digital photo and data measurement through NIS-Elements D 3.2. Cell size was measured totally at least 50 measurements each strain. The transmission electron microscope(TEM) procedure had made appropriate adjustments(Gkelis et al., 2005; Shao et al., 2011). Cyanobacterial culture solution during the exponential growth phase was centrifuged leaved a moderate sludge in 1.5 mL centrifuge tube, then added 2.5% glutaraldehyde to fixed 2 h at room temperature, centrifuged precipitation to collecting cells. Additionally, cells double fixed with glutaraldehyde in phosphate buffer PBS(0.1 mol/L, pH 7.2). After dehydrated, penetrated,embedded, the sample repaired to the right size and shape and sliced to about 60–100 nm thickness in the Leica ultra-thin slicing machine (EM UC7, Germany),then double staining with lead and uranium. The ultrastructure features of the sample were observed and photographed by TEM (Tecnai G220 TWIN,USA) with iTEM FEI software to selecting appropriate images at 200 kV.
2.3 Spectral measure of pigments
The absorption spectra of Merismopedia tenuissima CHAB 7021 was measured by the dual beam spectrophotometer with the simplistic quantitative filter technique (Mitchell, 1990). Distilled water was added into 10 mL culture solution in logarithmic phase for diluting to 250 mL uniform solution. Then the 250 mL uniform solution was filtered through the Whatman GF/FTMfilter made cells evenly distributed.In addition, 250 mL distilled water was filtered through Whatman GF/FTMfilter as blank to provide a spectral background. Then the filter samples were measured by the spectrophotometer (SHIMADZU,UV2550) from 300 nm to 800 nm.
2.4 Molecular and phylogenetic analysis
Total genomic DNA was extracted by reference to the cetyltriethylammnonium bromide (CTAB)method (Neilan et al., 1995) with actual situation adjustment. A pair of primers, PA (5′-AGAGTTTTGATCCTGGCTCAG-3′) (Neilan et al., 1997) and B23S (5′-CTTCGCCTCTGTGTGCC-TAGGT-3′)(Taton et al., 2003), were used to amplify 16S rRNA gene and internal transcribed spacer (ITS) between 16S rRNA and 23S rRNA gene sequences. Primers and 2×PCR Master Mix were provided from Qingke Biotechnology Company (Qingke, Beijing, China).PCR reaction system is a 50-μL volume reaction mixture containing 25 μL 2×PCR Master Mix, each positive and negative primer 10 pmol, DNA 1 μL and sterile water finally added to 50 μL. The thermal cycle reaction was set with an initial denaturation step at 95°C for 3 min, 35 cycles of DNA denaturation at 95°C for 30s, primer annealing at 58°C for 30s, strand extension at 72°C for 1 min 10 s, and final extension step at 72°C for 5 min.
The ampli fied products were used on 1% agarose gel electrophoresis, and ampli fication strips were detected at 300 nm of ultraviolet (Molecular Imager Gel Doc EX System, Newbio Industry). Then ampli fied products were puri fied and recycled with Biospin gel recycling kit (Omega, USA). Clone experiment was carried out according to the manufacturer’s protocol. The puri fied products were connected to the PMD18-T carriers and then transferred to the competent cells E. coli, following by spreading plate and selecting the positive single clone to be commercially sequenced (Qingke, Beijing, China).
DNA sequences, both examined in this study and obtained from GenBank, were aligned using CLUSTALW integrated into the BioEdit (Hall, 1999;Tamura et al., 2007). The most appropriate models of DNA sequence evolution were selected for both datasets by a hierarchical likelihood ratio test using MODELTEST version 3.7 (Posada and Crandall,1998). The neighbor-joining (NJ) tree and the maximum-parismony (MP) were constructed in MEGA 7.0.2 (Kumar et al., 2016). NJ phylogenetic tree was constructed by using the distance matrix calculated according to alignments under Kimura-2 substitution model with uniform nucleotide substitution rates both sites and lineages. Maximumlikelihood (ML) trees was constructed in PhyML 3.0(Guindon et al., 2010). The most appropriate models of DNA sequence evolution were selected by Smart Model Selection using heuristic strategies to avoid testing all models and options. (Lefort et al., 2017).Bootstrap analysis was performed 1 000 times for estimating degree of con fidence about each branch node. Escherichia coli LW1655F was used as the out group. The obtained 16S rRNA gene and ITS sequences were deposit in Genbank as MF680039 and MF 680076 respectively.
The primers set as mcyE-R4 (5′-AATTCTAAAGCCCAAAGACG-3′) and mcyE-F2 (5′-GAAATTTGTGTAGAAGGTGC-3′) (Rantala et al., 2004) were used for the detection of microcystin synthesis related gene.
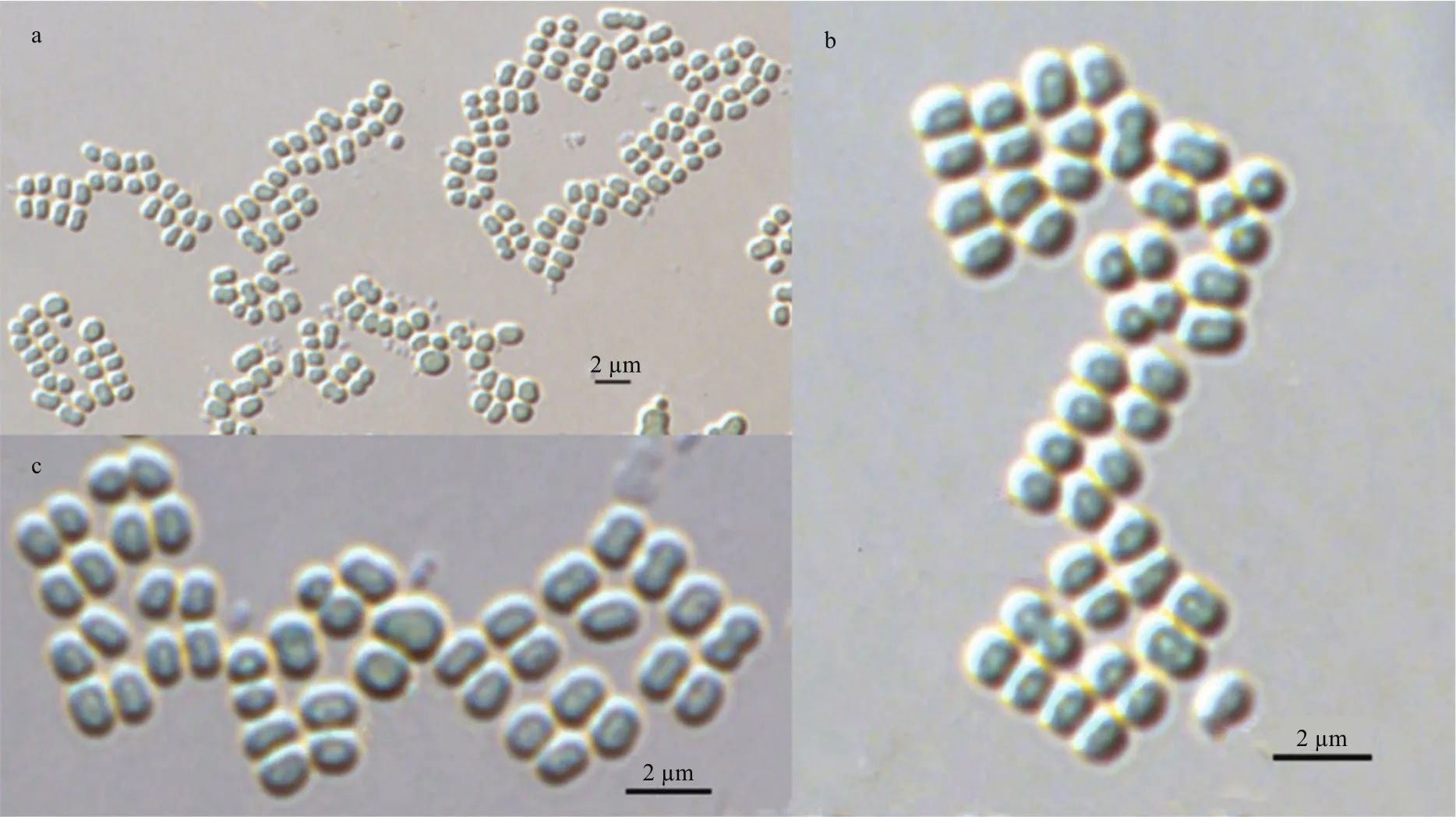
Fig.1 The light microscopic image of M. tenuissima CHAB 7021
3 RESULT
3.1 Morphological characteristics
According to the description of Komárek and Anagnostidis (1999), strain CHAB 7021 was morphologically identi fied as Merismopedia tenuissima (Figs.1, 2). Morphological characteristics of Merismopedia tenuissima CHAB 7021 were as follows: Cells blue-green, elliptical, spherical or hemispherical, with 0.82–0.93 μm wide and 0.97–1.28 μm long. Protoplast was uniform without gas vesicles. Usually four cells up to many cells, arranged regularly, aggregated regular group, like a layer of flat shape. Mucilaginous envelopes colourless, transparent and fragile outside groups. Proliferation was carried out in binary fission, commonly had two splitting± perpendicular planes.
Through the TEM, ultrastructural features of Merismopedia tenuissima CHAB 7021 can be observed as followings: cells have at least three parallel parietal thylakoids, these thylakoids form concentric circles surrounded in the protoplast. No gas vesicle was found in the protoplast and mucilaginous envelopes were transparent. Cellular division plane was obviously shown along the cell division (Fig.2).
3.2 Spectral analysis
As shown in Fig.3, absorption spectrum showed a peak at approximately 632 nm indicating as the presence of phycocyanin (PC). There are two obvious absorption peaks for chlorophyll a (Chl a) at 437 nm and 680 nm respectively.
3.3 DNA sequences and phylogenetic analysis
The 16S rDNA sequences of Merismopedia tenuissima CHAB 7021 (MF680039) have the size of 1 478 bp. NJ/MP/ML phylogenetic trees based on 16S rDNA sequences were constructed (Fig.4). Other cyanobacterial strains belonging to Chroococcales were mostly forming a cluster, while Merismopedia were obviously divided into three clades:Merismopedia tenuissima CHAB 7021, Merismopedia tenuissima OBB46S01 and Merismopedia sp.CENA106 were gathered into clade III, and clade I mainly included strains of M. punctata and M. glauca,and two other strains of Merismopedia sp. were gathered into clade II. The DNA similarities between clade I and II and between clade I and III were both below 90%.
The ITS sequence of Merismopedia tenuissima CHAB 7021 (MF680076) was obtained with 700 bp in length. ITS sequence of the M. tenuissima CHAB 7021 strain had less than 85% sequence similarity with any others ITS sequences existing in Genbank,and it contained two different tRNA: tRNAIleand tRNAAla.
No microcystin synthesis gene ( mcy) was molecularly detected in this study.
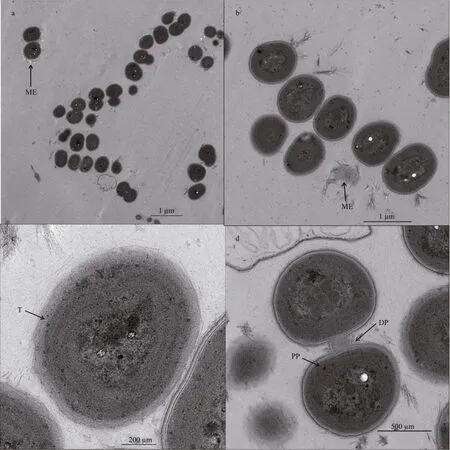
Fig.2 Transmission electron microscopy (TEM) images of M. tenuissima CHAB 7021
4 DISCUSSION
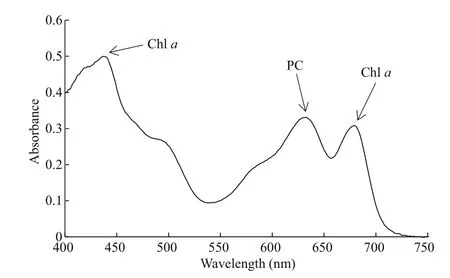
Fig.3 Pigment absorption spectrum of M. tenuissima CHAB 7021
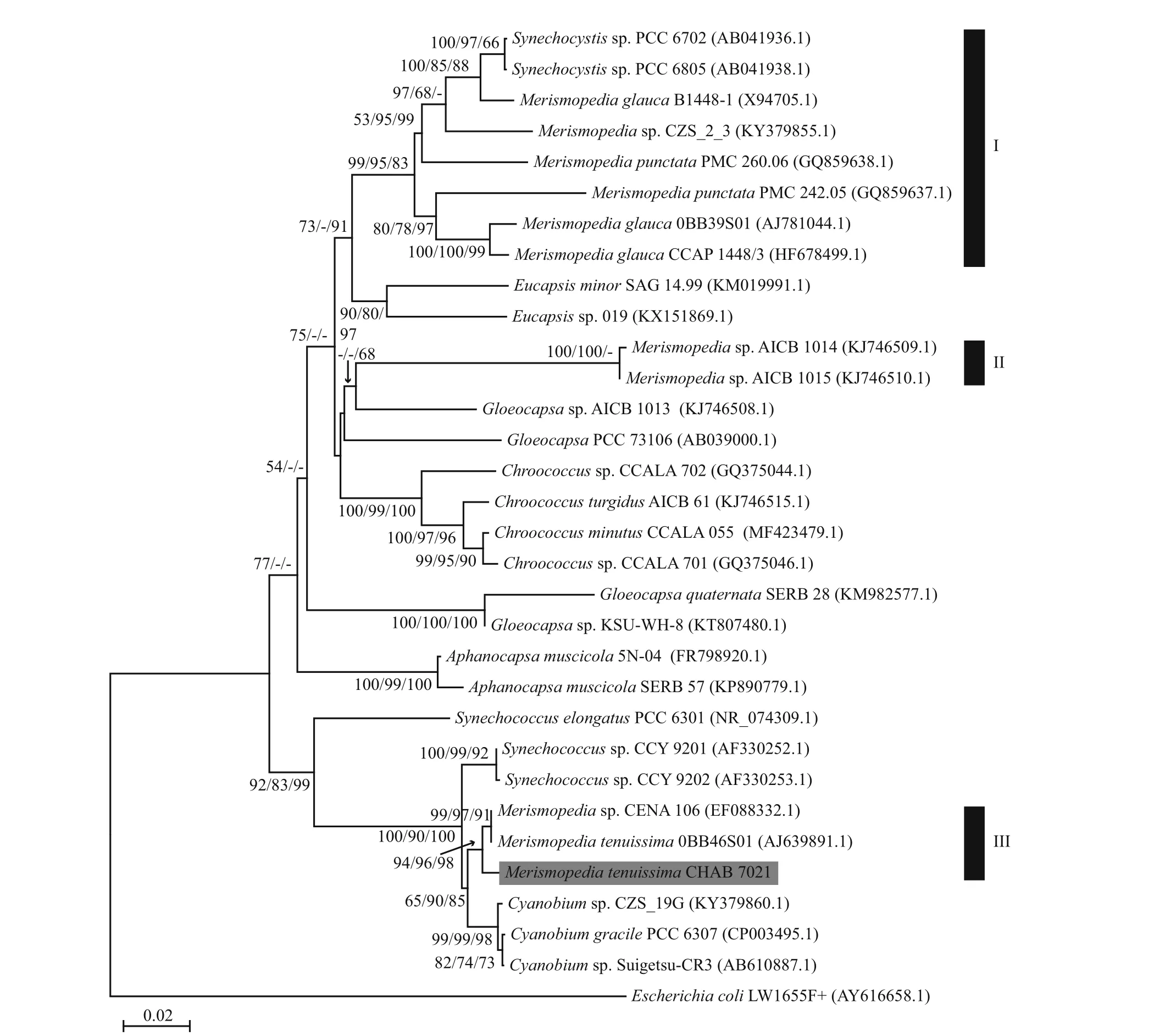
Fig.4 NJ/MP/ML phylogenetic tree constructed based on 16S rRNA gene sequences of M. tenuissima CHAB 7021 from this study and the others from GenBank
Availability of huge number of DNA sequences and usage of polyphasic characterization have led to extensive restructuring and revision of whole taxonomic system in cyanobacteria (species, genera,families, orders) and further changes are expected in the future (Komárek et al., 2014). Since 2000, more than sixty new cyanobacterial genera have been described by elucidating evolutionary process in cyanobacterial taxonomy based on the molecular phylogenetic analyses (Komáreket et al., 2014),especially in the filamentous (both heterocystous and non-heterocystous) cyanobacterial groups. In contrast,unicellular or unicellular colonial group (formerly named as Chroococcales) have been much less studied for their taxonomic revision at genus and species levels except Microcystis and Synechococcus. The genus Merismopedia is a case with many species but less information for the taxonomic evaluation, genetic diversity and phylogenetic relationship within the genus. Only 12 longer (>1 200 bp) 16S rRNA gene sequences of Merismopedia strains are currently available in Genbank, greatly hampering the taxonomic evaluation and better understanding for this genus. As described above, con flicting results on molecular phylogeny based on 16S rDNA sequences of the genus Merismopedia were gained from different strains by different researchers. In the first molecular phylogenetic study in the genus Merismopedia,Palinska et al. (1996) found 100% similarity in 16S rRNA gene sequences from the six strains morphologically corresponding to different Merismopedia species, indicating the phenotype variability of identical genotypes in the genus.Regarding this finding, Komárek and Anagnostidis(1999) suggested that stability of different phenotypes,ecotypes and variability of various species need the further study and con firmation. Later studies dealing with Merismopedia strains from different regions gave a controversy result, indicating the genotype variability of the same morphology (Rajaniemi-Wacklin et al., 2006; Thomazeau et al., 2010).Therefore, the current taxonomic state of the Merismopedia genus remains unsatisfactory. This problem may be attributed to no assignment of the type strain of the type species ( M. punctata) for the genus, and con flicting results from previous studies on molecular characterization of several Merismopedia strains led to further confusion. It is necessary to perform taxonomic and phylogenetic reevaluation on the Merismopedia genus, with starting at the type species— M. punctata.
This study is the first to examine the polyphasic characterization of the Merismopedia genus using cultivated strain from China. Our first task was to polyphasically characterize the Merismopedia tenuissima CHAB 7021 in order to accumulate the basic knowledge and information for the Merismopedia strains in China. The second task was to evaluate the molecular phylogeny on the Merismopedia genus using all the Merismopedia strains with enough length of 16S rRNA gene sequences (>1 200 bp) currently available in Genbank. From the 16S rDNA sequences based phylogenetic tree, 12 Merismopedia strains used were divided into three clades, indicating the polyphyly of the genus. The clade I, composed of six strains including type species M. punctata and M.glauca and two strains of Synechocystis, shared the similarity ranging from 91.3%–98.4% in 16S rRNA gene sequences. Clade II contained two nearly identical genotype strains isolated from the same site, but more characteristic description of these two strains were not provided, thus these two strains need further con firmation. In clade III, only strains of M.tenuissima were included. Merismopedia sp.CENA106 exhibited more than 99.1% similarity in 16S rRNA gene sequences with the two strains of M.tenuissima, plus description about the characteristic of Merismopedia sp. CENA106 from Furtado et al.(2009). Therefore, Merismopedia sp. CENA106 should be identi fied as M. tenuissima. Current morphological taxonomic system in cyanobacteria leads to difficulty in identifying the tiny species of the genus Merismopedia, since there exist several species with highly similar morphological characters especially at the narrow range of cell size (Komárek and Anagnostidis, 1999; Joosten, 2006). The identi fication of the strain CHAB 7021 as M.tenuissima in this study was based on the combined characters such as cellular arrangement in the colony,cell size and niche.
The obtained phylogenetic relationship in this study apparently indicated that the Merismopedia species in the current cyanobacterial taxonomic system are divided into several groups, implicating the need for taxonomic revision and division in the genus. According to the type species decisive criteria in cyanobacterial taxonomic revision at species level, the cluster I containing type species M. punctata represents the real Merismopedia species and some Synechocystis strains. The close relationship between Merismopedia and Synechocystis was also revealed in the study by Palinska et al. (1996). The species in cluster II and III with distant relationship to the cluster I should separate from the Merismopedia genus and may become new cyanobacterial genera, at least for the cluster III with three strains of M. tenuissima.However, larger genetic distance of 16S rRNA gene sequences existing in Merismopedia strains in the cluster I still confused the taxonomic status within Merismopedia genus, and therefore caused taxonomic revision difficult. It is hard to de fine which strain represent the real type species/strain since the two strains assigned as M. punctata showed 93.5%–95.4% similarity, even intermixed with the strains of M. glauca in the phylogenetic tree (Fig.4).M. glauca B1448-1 was distantly related to the other Merismopedia strains in the cluster I, and this freshwater strain was even found to have the identical 16S rRNA gene sequences with saline strains of M. elegans and M. punctata (Palinska et al., 1996), further complicating the determination of type species in this genus. The process for taxonomic revision in the Merismopedia genus still await for examination and further information on more strains of type species M. punctata.
5 CONCLUSION
The Merismopedia species are commonly found in aquatic environments with different trophic states,and some species were found as the dominant species in the eutrophic water. The taxonomic and phylogenetic evaluation on the Merismopedia genus in this study, by polyphasic charcterization on the M.tenuissima strain from China and all strains in the Genbank, brought some addition to the general view and basic knowledge for the classi fication and systematics of the genus Merismopedia, and further provided the future direction for the process of taxonomic revision in the genus.
6 DATA AVAILABILITY STATEMENT
The resulting for 16S rRNA gene sequence and ITS sequence were deposited in GenBank. The accession numbers of 16S rRNA gene sequence and ITS sequence were MF680039 and MF680076,respectively.
猜你喜欢
中国水产(2023年8期)2023-09-09 08:58:30
科学与财富(2019年11期)2019-08-06 19:20:34
南都娱乐周刊(2018年9期)2018-05-11 03:21:18
南都娱乐周刊(2018年8期)2018-04-26 05:54:00
恋爱婚姻家庭(2016年28期)2016-10-11 09:33:49
恋爱婚姻家庭(2016年10期)2016-10-10 21:04:32
北京电视周刊(2015年52期)2015-05-30 10:48:04
南都娱乐周刊(2014年39期)2014-11-05 09:11:49
意林(2011年12期)2011-05-14 06:40:44
 Journal of Oceanology and Limnology2018年4期
Journal of Oceanology and Limnology2018年4期
- Journal of Oceanology and Limnology的其它文章
- Editorial Statement
- Effects of seawater acidi fication on the early development of sea urchin Glyptocidaris crenularis*
- Dietary effects of A zolla pinnata combined with exogenous digestive enzyme (Digestin™) on growth and nutrients utilization of freshwater prawn, Macrobrachium rosenbergii(de Man 1879)
- Preliminarily study on the maximum handling size, prey size and species selectivity of growth hormone transgenic and non-transgenic common carp Cyprinus carpio when foraging on gastropods*
- Hydrodynamic characteristics of the double-winged otter board in the deep waters of the Mauritanian Sea*
- De novo transcriptome sequencing reveals candidate genes involved in orange shell coloration of bay scallop Argopecten irradians*