
miR-135b对LZST1表达及肺癌细胞转移行为的影响
2018-07-02 01:37杜芝洋杜发旺
解放军医药杂志 2018年6期
杜芝洋,杜发旺
目前,肺癌是呼吸系统中最常见的恶性肿瘤,是医疗保险人群中排名前五的高花费肿瘤,在临床上主要通过手术或全身化疗和放疗进行治疗[1]。在免疫功能低下的老年肺癌患者中,更容易出现较高的转移风险[2]。目前,转移性肺癌的治疗效果很不理想,化疗效果也差强人意[3]。据相关报道,晚期转移肺癌的5年生存率不超过10%[4]。因此,目前需要研究参与肺癌侵袭和转移的分子靶标,以及晚期转移性肺癌新的治疗策略。
miRNA是一种小的非编码RNA分子,可以通过结合靶mRNA的3'端非翻译区来抑制翻译或诱导mRNA切割[5-6]。然而,关于肺癌中miRNA表达的临床转化成果较少[7-8]。有相关肺癌组织的miRNA芯片分析显示,肺癌组织中有9种miRNA的表达发生明显变化,其中miR-135b的表达在肺癌组织和配对正常组织中变化最大(上调8.5倍)[9]。并且miR-135b与肿瘤的生长、生存期、运动性和侵袭性有密切的关联[10],在多种其他肿瘤(乳腺癌、肺癌、结肠癌和前列腺)组织中存在明显表达差异[11-14]。在不同组织来源的恶性肿瘤中,miR-135b表现的致癌性或抑癌性,具有一定的组织特异性[15]。在头颈部,miR-135b通过激活低氧诱导因子-1α来刺激癌细胞增殖,集落形成和血管生成,可能发挥肿瘤启动子的作用[16]。亮氨酸拉链肿瘤抑制基因1(LZTS1)被描述为促肿瘤基因[17]。功能研究表明,LZTS1的一个或两个等位基因的过表达可以导致小鼠自发性肿瘤发生,并且重新导入LZTS1抑制因子可以抑制裸鼠体内的致瘤性[18]。本研究为了探讨miR-135b对LZTS1在肺癌组织中表达的影响,及其对肺癌A549细胞迁移和侵袭行为的影响,为肺癌发生和进展的机制研究提供理论参考。
1 材料与方法
1.1组织标本、细胞株和试剂 在本医院获得肺癌患者书面知情同意后,采集肺癌组织和癌旁正常组织样本,石蜡包埋相关组织用于免疫组织化学研究。本研究获得医院审查委员会的批准。人类肺癌A549细胞购自复旦大学细胞培养中心,在37℃下5% CO2培养箱中进行细胞培养。RPMI 1640细胞培养基购自美国Gibco公司,胎牛血清购自杭州四季青生物材料研究所,Transwell侵袭小室购自美国Corning,Matrigel基质胶购自美国BD公司,脂质体Lipofectamine 2000购自Santa Cruz公司。LZST1一抗购自美国Abcam公司。HOTAIR+siRNA和miR-135b-mimic购自上海吉玛生物科技有限公司。Trizol购自美国Ambion公司,逆转录试剂盒(FSQ-101)购自日本TOYOBO公司,PCR试剂盒购自美国Sigma公司。荧光素酶活性检测试剂盒购自Promega公司。荧光素酶报告载体由Promega公司合成。
1.2PCR分析 使用mirVana miRNA分离试剂盒从肺癌组织和癌旁正常组织中分离总RNA。使用Nano-2000分光光度计测定RNA浓度和纯度。miR-135b引物序列:正向5'-GGCGTGAGGCTTGAGGGCT-3',反向5'-GGCGTAGGCGATTGGGGATCG-3'。LZST1引物序列:正向5'-ATTTAAGGAGCGGATTTAGC-3',反向5'-TTTTCGAGTCGAAACACACT-3'。以总RNA为模板,逆转录cDNA,反应条件按照cDNA合成试剂盒使用说明进行设置,反应条件:95℃预变性10 min、95℃变性15 s、60℃退火32 s,循环50次后检测其溶解曲线,检测完成后,通过计算机系统自动分析各样本Ct值,采用RQ=2-ΔΔCt计算miRNA的相对表达量。实验重复3次。
1.3慢病毒转染 根据慢病毒转染说明的方案,使用10 nmol的miR-135b-inhibitor和miR-135b-mimic模拟物转染肺癌A549细胞,分为miR-135b-inhibitor组和miR-135b-mimic组;使用相应的NC模拟物进行阴性对照。使用高容量RNA-to-cDNA试剂盒和Power SYBR GreenMasterMix对转染细胞中的miR-135b mRNA表达进行qRT-PCR分析。用于miR-135b基因的PCR扩增的引物如下:正向5'-ACCTCTAGAAACCCAGAACTCA-3',反向5'-TCCAGAAGAGCCCATATCACTA-3'。使用U6作为内参照:正向5'-GCGCCCAATACGACCAA-3',反向5'-CTCTCTGCTCCTCCTGTTC-3'。使用热循环仪通过qRT-PCR扩增基因转录物。条件如下:在50℃下UNG温育2 min和95℃下聚合酶活化10 min,然后95℃ 15 s和60℃ 1 min,进行45个循环。通过计算机系统自动分析各样本Ct值,采用RQ=2-ΔΔCt计算miRNA的相对表达量。实验重复3次。
1.4Western blotting蛋白质印迹分析 用十二烷基硫酸钠-聚丙烯酰胺凝胶电泳分离细胞总蛋白,并将其转移到硝酸纤维素膜。使用5%脱脂牛奶充分封闭膜,并与一抗(LZST1浓度1∶1500),内参GAPDH(浓度1∶2000),4℃下孵育过夜。次日,TBST充分冲洗过后,使用二抗辣根过氧化物酶标记物(浓度1∶2000)孵育2 h,TBST充分冲洗过后,使用化学发光试剂ECL显影液检测目标蛋白的表达水平。实验重复3次。
1.5划痕愈合实验 将转染细胞和对照组细胞培养至对数生长期。使用200 μl无菌移液器尖端以标准方式将单层细胞划伤创建无细胞区域。将培养基吸出并用新鲜的完全培养基代替,然后将细胞在37℃孵育48 h。48 h后记录并拍照细胞的迁移情况。在每个实验条件下以5个随机间隔测量迁移角化细胞之间的迁移残余间隙,以实际迁移距离与原始擦伤宽度的百分比作为比较数据。
1.6Transwell侵袭实验 使用24孔板进行细胞侵袭测定。将转染细胞和对照组细胞培养至对数生长期,重新悬浮在无血清培养基中并以每孔1×104个细胞的密度加入到上室中,底室含有10%胎牛血清和正常浓度生长因子的培养基。培养24 h后,通过抽吸基质胶去除上部孔中的细胞并用棉签擦拭膜的顶部。使用差异快速染色试剂将膜下侧的细胞固定,进行染色。在光学显微镜下计数细胞侵袭数目,每个膜观察5个显微镜视野。实验重复3次。
1.7免疫组织化学检测 将6 μm厚组织切片在10%中性福尔马林中固定10 min,然后用PBS洗涤2次。在室温下用1%牛血清白蛋白封闭载玻片1 h,然后用1∶200稀释度的LZTS1兔多克隆抗体在室温下孵育1 h。以相同稀释度的正常兔IgG作为阴性对照。吸出一抗并在PBS中洗涤2次后,用亲和纯化的驴抗兔IgG以1∶1000的稀释度孵育LZTS1 30 min。然后洗涤载玻片并用DAPI以1∶1000的稀释度染色1 min。使用95%乙醇固定细胞2 min后在显微镜下分析LZTS1染色强度。
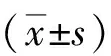
2 结果
2.1miR-135b和LZST1的表达情况 肺癌组织和癌旁正常组织中miR-135b表达水平分别为4.15±0.76和1.28±0.29,LZST1表达水平分别为6.58±0.89和1.93±0.21。肺癌组织中miR-135b和LZST1表达水平明显高于癌旁正常组织(P<0.05)。免疫组化结果显示,LZST1蛋白在肺癌组织中表达阳性,在癌旁正常组织中表达阴性。表明miR-135b和LZST1在肺癌中扮演一定的促癌作用。
2.2miR-135b对肺癌A549细胞LZST1蛋白表达的影响 miR-135b-inhibitor组、miR-135b-mimic组和阴性对照组的LZST1蛋白表达水平分别为1.03±0.29、4.03±1.02和1.86±0.76。miR-135b-mimic组的LZST1蛋白表达水平高于miR-135b-inhibitor组和阴性对照组,且阴性对照组高于miR-135b-inhibitor组(P<0.05)。结果表明过表达miR-135b后LZST1蛋白的表达上调,抑制miR-135b表达后LZST1蛋白的表达下调。见图1。
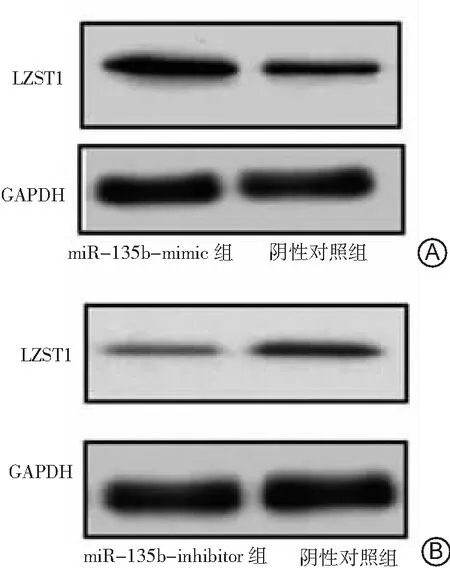
图1miR-135b对肺癌A549细胞LZST1蛋白表达的影响
A.miR-135b-mimic组和阴性对照组;B.miR-135b-inhibitor组和阴性对照组;LZST1为亮氨酸拉链肿瘤抑制基因1
2.3miR-135b对肺癌A549细胞迁移能力的影响 miR-135b-inhibitor组、miR-135b-mimic组和阴性对照组的实际迁移距离百分比分别为(9.8±1.9)%、(82.8±9.3)%和(33.6±4.2)%。miR-135b-mimic组的实际迁移距离百分比高于miR-135b-inhibitor组和阴性对照组,且阴性对照组高于miR-135b-inhibitor组(P<0.05)。结果表明过表达miR-135b后肺癌A549细胞的迁移能力增强,抑制miR-135b表达后肺癌A549细胞的迁移能力减弱。见图2。

图2miR-135b对肺癌A549细胞迁移能力的影响
2.4miR-135b对肺癌A549细胞侵袭能力的影响 miR-135b-inhibitor组、miR-135b-mimic组和阴性对照组侵袭至膜下的细胞数目分别为42.6±5.1、342.1±21.9和101.3±7.6。miR-135b-mimic组侵袭至膜下的细胞数目高于miR-135b-inhibitor组和阴性对照组,且阴性对照组高于miR-135b-inhibitor组(P<0.05)。结果表明过表达miR-135b后肺癌A549细胞的侵袭能力增强,抑制miR-135b表达后肺癌A549细胞的侵袭能力减弱。见图3。
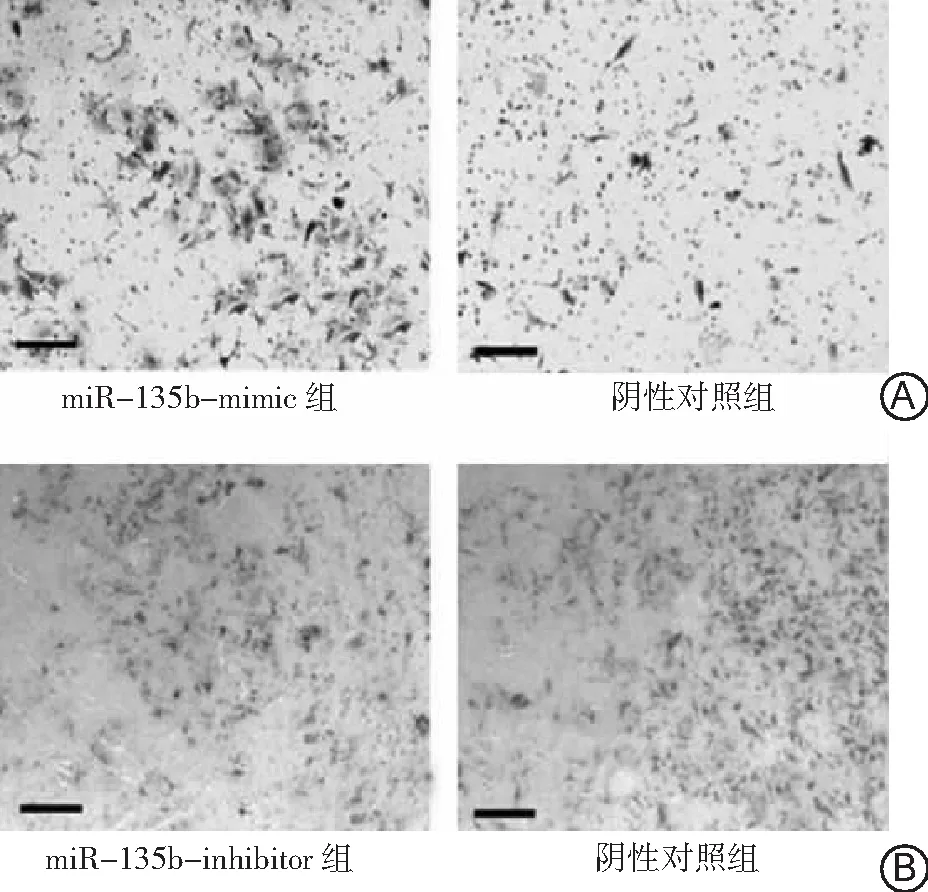
图3miR-135b对肺癌A549细胞侵袭能力的影响(×40)
A.miR-135b-mimic组和阴性对照组;B.miR-135b-inhibitor组和阴性对照组
3 讨论
最近的研究表明,部分miRNA不仅对促进实体器官肿瘤的生成以及转移瘤的发展具有一定的调控作用,而且对肿瘤的进展和转移也可能发挥重要作用[19]。全基因组测序研究表明肺癌的基因表达谱存在巨大差异,从而引起其他的分子发展机制,如影响肺癌患者的基因表达及miRNA改变[20]。
miR-135b与多种类型恶性肿瘤的进展有关[21]。本研究发现,miR-135b在肺癌组织中表达水平高于癌旁正常组织,同时可以调控下游LZST1蛋白的表达,过表达和抑制miR-135b后,LZST1蛋白水平也相应上调和下调,推测肿瘤基因LZTS1可能是miR-135b的直接靶基因[22]。进而推测miR-135b可能通过靶向调控LZTS1表达水平,而调控肺癌细胞的迁移和侵袭行为。有学者研究指出,miR-135b不直接参与肺癌A549增殖行为的直接调控,表明miR-135b是通过调控LZTS1肿瘤基因发挥功能的启动子。miR-135b表达从正常到息肉再到恶性组织的递增表明miR-135b失调是一种早期癌变因素,随着细胞分化异常逐渐表达增强[23]。因此,本实验研究的数据表明miR-135b可能通过调节肿瘤因子(如LZTS1等)干扰肺癌的进展过程。
最近研究证明,LZTS1为miR-135b的靶基因,并且影响多种肿瘤的生长,运动性和侵袭性[24]。然而,LZTS1在肺癌进展过程中的作用以及与miR-135b表达的关系未被明确探讨。LZTS1是miR-135b在肺癌中的肿瘤调控基因之一,但miR-135b/LZTS1轴在肺癌中的关系及其功能尚不清楚。本实验发现,在癌旁正常肺组织中LZTS1表达普遍较低,而在肺癌组织中LZTS1表达明显上调。这与先前报道的其他多种肿瘤类型(如乳腺癌和肾癌)中LZTS1表达降低相符合[25]。进而本研究还在蛋白水平上发现LZTS1蛋白表达水平与miR-135b表达成正相关。证明过表达miR-135b可以增加LZTS1蛋白的表达,导致肺癌细胞的迁移和侵袭能力增强。
还有研究显示,人类肿瘤基因通常作为细胞周期的调节剂起作用,通过上调miR-135b直接影响LZTS1蛋白的表达可导致肿瘤细胞M期细胞周期机制的阻断,导致遗传变化的积累,引起遗传不稳定性和细胞非整倍性的出现[26]。相关研究发现,通过敲除小鼠体内的LZTS1基因会发展出各种肿瘤,表明非整倍体表型可以驱动体内细胞恶性转化[27]。马瑞宏等[28]研究表明,抑制miR-135b不影响体外肺癌细胞的增殖能力,说明在影响恶性细胞迁移和侵袭功能的同时miR-135b的丢失不会直接干扰肿瘤生长。
综上所述,miR-135b/LZTS1信号轴在肺癌进展中起重要调控作用,并且miR-135b可能作为临床肺癌生物治疗的有效靶点,后续实验需要进一步研究miR-135b的作用机制,以逐步开发肺癌及肺癌转移瘤的新治疗方式。
[参考文献]
[1] Shi Y, Au J S, Thongprasert S,etal. A prospective, molecular epidemiology study of EGFR mutations in Asian patients with advanced non-small-cell lung cancer of adenocarcinoma histology (PIONEER)[J].J thorac Oncol, 2014,9(2):154-162.
[2] Raaschou-Nielsen O, Andersen Z J, Beelen R,etal. Air pollution and lung cancer incidence in 17 European cohorts: prospective analyses from the European Study of Cohorts for Air Pollution Effects (ESCAPE)[J].Lancet Oncol, 2013,14(9):813-822.
[3] Malvezzi M, Bertuccio P, Rosso T,etal. European cancer mortality predictions for the year 2015: does lung cancer have the highest death rate in EU women?[J].Ann Oncol, 2015,26(4):779-786.
[4] Argirion I, Weinstein S J, Mannisto S,etal. Serum insulin, glucose, indices of insulin resistance, and risk of lung cancer[J].Cancer Epidemiol Biomarkers Prev, 2017,26(10):1519-1524.
[5] Kasinski A L, Kelnar K, Stahlhut C,etal. A combinatorial microRNA therapeutics approach to suppressing non-small cell lung cancer[J].Oncogene, 2015,34(27):3547-3555.
[6] Sozzi G, Boeri M, Rossi M,etal. Clinical utility of a plasma-based miRNA signature classifier within computed tomography lung cancer screening: a correlative MILD trial study[J].J Clin Oncol, 2014,32(8):768-773.
[7] Peng Y, Dai Y, Hitchcock C,etal. Insulin growth factor signaling is regulated by microRNA-486, an underexpressed microRNA in lung cancer[J].Proc Natl Acad Sci U S A, 2013,110(37):15043-15048.
[8] Liu Z L, Wang H, Liu J,etal. MicroRNA-21 (miR-21) expression promotes growth, metastasis, and chemo-or radioresistance in non-small cell lung cancer cells by targeting PTEN[J].Mol Cell Biochem, 2013,372(1-2):35-45.
[9] Su W, Mo Y, Wu F,etal. miR-135b reverses chemoresistance of non-small cell lung cancer cells by downregulation of FZD1[J].Biomed Pharmacother, 2016,84:123-129.
[10] Nezu Y, Hagiwara K, Yamamoto Y,etal. miR-135b, a key regulator of malignancy, is linked to poor prognosis in human myxoid liposarcoma[J].Oncogene, 2016,35(48):6177-6188.
[11] Hua K, Jin J, Zhao J,etal. miR-135b, upregulated in breast cancer, promotes cell growth and disrupts the cell cycle by regulating LATS2[J].Int J Oncol, 2016,48(5):1997-2006.
[12] Valeri N, Braconi C, Gasparini P,etal. microRNA-135b promotes cancer progression acting as a downstream effector of oncogenic pathways in colon cancer[J].Cancer Cell, 2014,25(4):469-483.
[13] Xiao S, Yang Z, Lv R,etal. miR-135b contributes to the radioresistance by targeting GSK3β in human glioblastoma multiforme cells[J].PloS one, 2014,9(9):e108810.
[14] Aakula A, Leivonen S K, Hintsanen P,etal. MicroRNA-135b regulates ERα, AR and HIF1AN and affects breast and prostate cancer cell growth[J].Mol Oncol, 2015,9(7):1287-1300.
[15] Adai A, Szafransak-Schwarzbach A, Andruss B,etal. Methods and compositions involving miR-135b for distinguishing pancreatic cancer from benign pancreatic disease: U.S. Patent 9,644,241[P].2017-5-9.
[16] Umezu T, Tadokoro H, Azuma K,etal. Exosomal miR-135b shed from hypoxic multiple myeloma cells enhances angiogenesis by targeting factor-inhibiting HIF-1[J].Blood, 2014,124(25):3748-3757.
[17] Lovat F, Ishii H, Schiappacassi M,etal. LZTS1 downregulation confers paclitaxel resistance and is associated with worse prognosis in breast cancer[J].Oncotarget, 2014,5(4):970-977.
[18] Xu Z, Wang T. miR-214 promotes the proliferation and invasion of osteosarcoma cells through direct suppression of LZTS1[J].Biochem Biophys Res Commun, 2014,449(2):190-195.
[19] George S, Chaudhery V, Lu M,etal. Sensitive detection of protein and miRNA cancer biomarkers using silicon-based photonic crystals and a resonance coupling laser scanning platform[J].Lab Chip, 2013,13(20):4053-4064.
[20] Frampton G M, Fichtenholtz A, Otto G A,etal. Development and validation of a clinical cancer genomic profiling test based on massively parallel DNA sequencing[J].Nat biotechnol, 2013,31(11):1023-1031.
[21] Beltran H, Yelensky R, Frampton G M,etal. Targeted next-generation sequencing of advanced prostate cancer identifies potential therapeutic targets and disease heterogeneity[J].Eur Urol, 2013,63(5):920-926.
[22] Mori M, Triboulet R, Mohseni M,etal. Hippo signaling regulates microprocessor and links cell-density-dependent miRNA biogenesis to cancer[J].Cell, 2014,156(5):893-906.
[23] Pei H, Jin Z, Chen S,etal. MiR-135b promotes proliferation and invasion of osteosarcoma cells via targeting FOXO1[J].Mol Cell Biochem, 2015,400(1-2):245-252.
[24] Olasz E B, Seline L N, Schock A M,etal. MicroRNA-135b regulates leucine zipper tumor suppressor 1 in cutaneous squamous cell carcinoma[J].PLoS One, 2015,10(5):e0125412.
[25] Lovat F, Ishii H, Schiappacassi M,etal. LZTS1 downregulation confers paclitaxel resistance and is associated with worse prognosis in breast cancer[J].Oncotarget, 2014,5(4):970-977.
[26] Zhou W, He M R, Jiao H L,etal. The tumor-suppressor gene LZTS1 suppresses colorectal cancer proliferation through inhibition of the AKT-mTOR signaling pathway[J].Cancer Lett, 2015,360(1):68-75.
[27] He Y, Liu X. The tumor-suppressor gene LZTS1 suppresses hepatocellular carcinoma proliferation by impairing PI3K/Akt pathway[J].Biomed Pharmacother, 2015,76:141-146.
[28] 马瑞宏,黄颖秋,刘彩刚.FEZ1/LZTS1基因与原发性肝癌相关性研究[J].海南医学院学报,2013,19(2):148-151.
猜你喜欢
中国临床医学影像杂志(2022年2期)2022-05-25
中国药学药品知识仓库(2022年1期)2022-03-23
湖南畜牧兽医(2021年6期)2022-01-24
食品安全导刊(2021年21期)2021-08-30
天津医科大学学报(2019年6期)2019-08-13
中国医学影像学杂志(2018年9期)2018-10-17
中国人兽共患病学报(2018年7期)2018-07-31
中国医药指南(2017年3期)2017-11-13
医学研究杂志(2015年12期)2015-06-10
医学研究杂志(2015年12期)2015-06-10
