
链霉菌30702的鉴定及其生防特性
2018-06-29 06:47丛子文焦敬华周双清黄东益吴文嫱许云夏薇张荣萍黄小龙
生物技术通报 2018年6期
丛子文 焦敬华 周双清 黄东益 吴文嫱 许云 夏薇 张荣萍 黄小龙
(海南大学热带农林学院,海口 570228)
链霉菌属(Streptomyces)是放线菌中最大的一类成员,也是产抗生素最多的放线菌菌属[1]。目前链霉菌属有效发表种823个,亚种38个(http://www.bacterio.net/streptomyces.html)。链霉菌属有效发表种的模式菌株在16S rRNA基因序列系统发育树上分散归簇于不同的进化分枝(Clade)[2],每个clade中的菌株大多具有相似的表型和基因型特征以及生物活性,但又存在种间的区别。紫黑链霉菌进化分枝(Streptomyces violaceusniger 16S rRNA gene clade)是其中一个重要的分枝,由Sembiring 等[3]推荐建立,当前包含26个有效发表种[4]。该进化分枝的放线菌普遍产生丰富的活性物质包括抗菌剂[5-11]、抗肿瘤剂[12-13]、生防制剂[14-19]、生物催化剂[20]、磷脂酶抑制剂[21]和免疫抑制剂[22],在医药和农用抗生素的开发上得到了广泛的应用。
近年来可培养及未培养的研究发现,紫黑链霉菌进化分枝放线菌在植物根际、落叶堆、海泥等具有广泛的分布[23]。Sembiring 等[3]从南洋楹(Paraserianthes falcataria)植物根际分离到大量紫黑链霉菌进化分枝放线菌新物种。Sahin等[24]发现紫黑链霉菌进化分枝放线菌在土耳其豆类植物根际具有多样性的分布。Kang等[25]从三齿蒿(Artemisia tridentate)根际分离到的紫黑链霉菌进化分枝放线菌菌株对植物病原真菌和白色念珠菌具有广谱的抗性。我们前期的研究证实,紫黑链霉菌进化分枝的放线菌菌株在海南药用植物根际也有一定的分布[26],并从药用植物海南粗榧根际分离得到一株紫黑链霉菌进化分枝的链霉菌30702,该菌株对山药炭疽病菌、芒果炭疽病菌、香蕉枯萎病菌等植物病原真菌都显示出强烈的拮抗活性[27]。本研究对其种水平的分类地位和生防特性进行深入研究,以期为该菌株在植物生防菌剂的开发与应用上提供参考。
1 材料与方法
1.1 材料
1.1.1 菌株 供试菌株30702,由本实验室前期从热带药用植物粗榧根际土壤中分离得到。供试山药炭疽病菌(Colletotrichum gloeosporioides)由本实验保存。
1.1.2 培养基 发酵培养基:可溶性淀粉25 g,黄豆粉20 g,MgSO4.7H2O 0.6 g,CaCO30.2 g,水1 000mL,pH 7.0。
1.2 方法
1.2.1 链霉菌30702的DNA提取、PCR扩增及测序 参照周双清等[28]方法进行菌株DNA 的提取。选取16S rRNA 基因和5个看家基因atpD、gyrB、recA、rpoB和trpB为实验对象。它们在放线菌的基础代谢中行使不同的功能。其中atpD 基因编码F0F1-ATP 合成酶β亚基;gyrB基因编码DNA解旋酶β亚基;recA 基因编码重组酶A;rpoB 基因编码RNA 聚合酶β亚基;trpB基因编码色氨酸合成酶β亚基。各基因序列PCR 扩增的条件参照Guo等[29]的方法,目标片段(TaKaRa试剂盒)回收、纯化后与pMD18-T载体连接,转化感受态细胞后,筛选阳性克隆送广州天一辉远基因科技有限公司测序。
1.2.2 链霉菌30702系统进化树构建及MLSA多位点序列分析 将获得的16S rRNA 基因序列登录EzTaxon 数据库中作blast 序列比对,同时下载相关相似菌株序列,用Mega6.0 软件构建系统进化树。获得的atpD、gyrB、recA、rpoB和trpB基因序列,参照Guo等[29]的方法,首尾相连拼接成多基因序列,并在GenBank数据库下载相关菌株的5个看家基因序列拼接成多基因序列,采用Mega6.0 软件构建系统进化树,利用Kimura 2-parameter(K2P)模型计算多位点序列进化距离。
1.2.3 链霉菌30702形态及培养特征分析 培养特征描述所用培养基参照国际链霉菌规划中的标准培养基[30]。将供试菌株30702分别接种到ISP2、ISP3、ISP4、ISP5、ISP6和ISP7琼脂培养基,28℃恒温培养7-21 d,观察并记录放线菌菌落形态及培养特征。
1.2.4 链霉菌30702生理生化特征 温度试验、pH试验、耐盐度测定、脲酶试验、明胶液化、硝酸盐还原、碳源利用实验和氮源利用实验参照文献[31]的方法进行。
1.2.5 链霉菌30702细胞壁化学分析 供试菌株30702的细胞壁氨基酸和糖组分分析参照Lechevalier等[32]的方法。
1.2.6 链霉菌30702的产酶特性 供试菌株30702产蛋白酶、纤维素酶、几丁质酶、β-葡聚糖酶等检测方法参照文献[33]。
1.2.7 链霉菌30702促生长特性 供试菌株30702产铁载体、解磷活性、产1-氨基环丙烷-1-羧酸(ACC)脱氨酶以及体外促进植物生长等方法参照文献[34]。其中体外促进植物生长试验具体如下:将菌株30702接种在MS 培养基上,28℃培养3 d,3 d后将拟南芥种子种植在MS培养基的另一端,距离菌株30702 5 cm,22℃,16 h光照,8 h黑暗交替培养7 d。以未接菌的MS 培养基为对照,电镜观察拟南芥幼苗的根及根毛。扫描电镜照片由海南大学测试中心完成。
1.2.8 菌株30702发酵粗提物制备 将菌株30702接种发酵培养基中(200 mL培养基/500 mL三角瓶),180 r/min,28℃恒温培养7 d。将发酵液离心(4℃,4 000 r/min,10 min),除去上清,得到菌体。向菌体中加入50 mL丙酮。摇床上振荡浸提2 h,离心收集上清液,旋蒸蒸发仪蒸干得发酵粗提物。
1.2.9 菌株30702发酵粗提物 抑制山药炭疽菌孢子萌发、菌丝生长 取10 μL山药炭疽菌的孢子悬浮液(孢子浓度为107个/mL)与 20 μL的葡萄糖溶液(1g/L),以及一定量的发酵粗提物混合均匀,发酵粗提物终浓度分别设置为 0 μg/ mL,15 μg/ mL,25 μg/mL,50 μg/ mL,75 μg/ mL 和 100 μg/ mL,滴加在无菌的血球计数板上,放置在无菌的培养皿里。28℃保湿培养24 h,观察孢子萌发和菌丝形态。
2 结果
2.1 菌株30702的菌种鉴定
2.1.1 菌株30702 的16S rRNA基因序列分析 PCR扩增菌株30702的16S rRNA基因序列,获得其序列全长为1 481 bp。在EzTaxon 数据库中进行比对检索,发现菌株30702与紫黑链霉菌进化分枝的有效发表种的序列最相似,其中相似性前三位的菌株 为 Streptomyces solisilvae HNM0141T,Streptomyces malaysiensis NBRC 16446T和Streptomyces samsunensis M1463T,相似性分别为100%、99.4%和98.8%。选取相似性较高的标准菌株序列,利用MEGA6.0 软件采用N-J 法(重复度1 000)构建系统进化树(图1)。结果表明菌株30702与S. solisilvae HNM0141T,S. malaysiensis NBRC 16446T和S. samsunensis M1463T聚在同一个大的分支上,说明菌株30702与这3株菌的亲缘关系较近,其中菌株30702与S. solisilvae HNM0141T又聚成一个小分支,表明S. solisilvae HNM0141T是菌株30702亲缘关系最近的标准菌株,因此选取这3株作为下述实验的参照菌株。
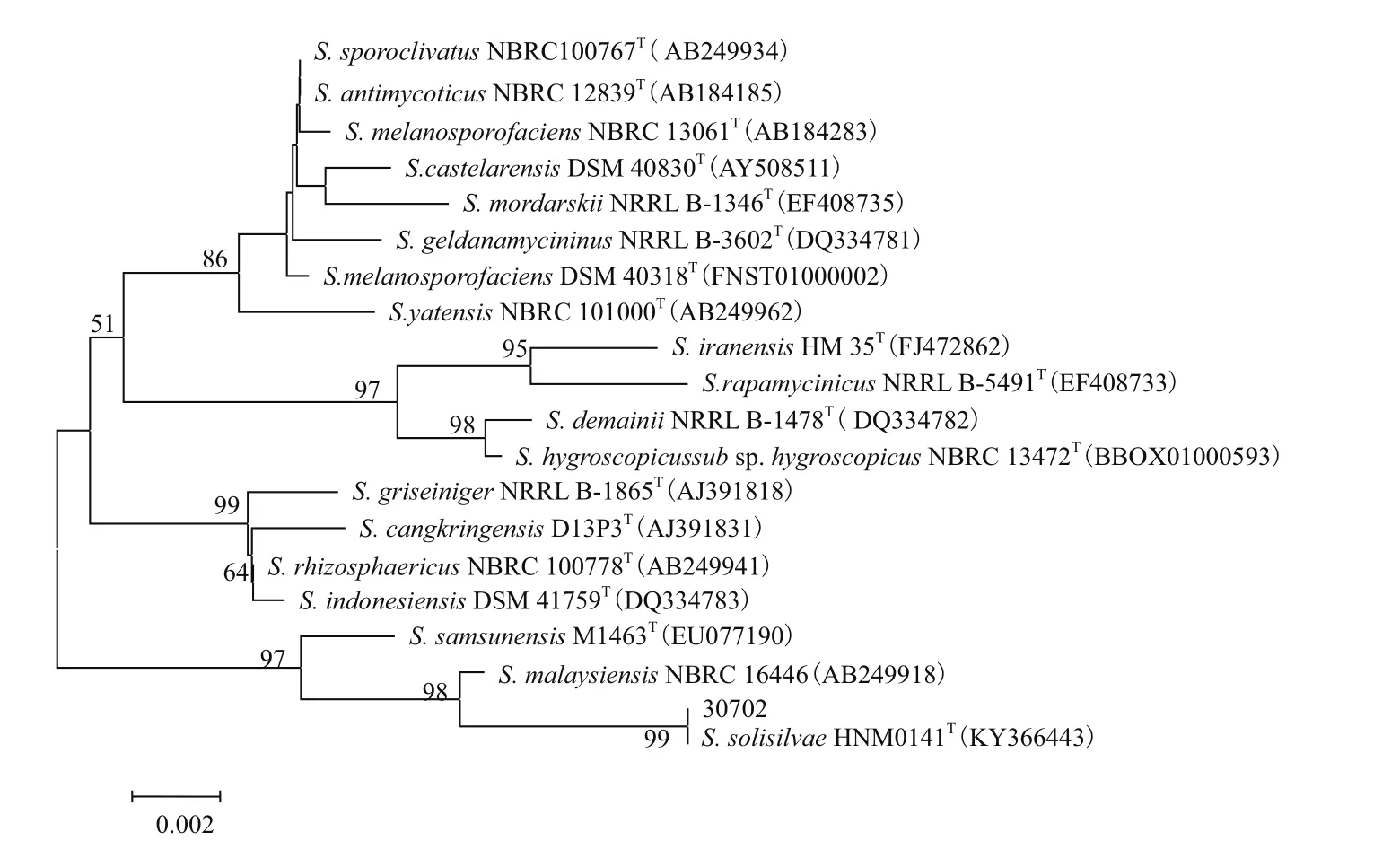
图1 菌株30702的16S rRNA基因序列系统进化树
2.1.2 菌株30702 的MLSA分析 扩增获得菌株30702的5个看家基因atpD、gyrB、recA、rpoB和trpB基因序列,并首尾相连拼接成多基因序列与菌株S. solisilvae HNM0141,S. malaysiensis NBRC 16446和S. samsunensis M1463的多基因拼接序列等构建系统进化树(图2)。菌株30702与链霉菌S.solisilvae HNM0141,S. malaysiensis NBRC 16446和S. samsunensis M1463同样聚在一单独的分支上,其中菌株30702与链霉菌S. solisilvae HNM0141又聚在同一个小分支上。遗传进化距离计算(表1)表明,菌株30702与链霉菌S. solisilvae HNM0141的多基因序列进化距离为0.004,与其他两株菌的多基因序列进化距离均大于0.007(分别为0.118和0.112)。

图2 菌株30702基于5个看家基因串联的进化树

表1 菌株30702 与参照菌株的多基因序列进化距离
2.1.3 形态及培养特征分析 菌株30702在ISP2、ISP3、ISP4、ISP5、ISP6和ISP7六种培养基上生长良好,气生菌丝为综色、粉白色或灰色,基内菌丝为深灰色、棕黄色、亮黄色和黄绿色;在所有培养基上不产生色素(表2)。菌株30702与S. solisilvae HNM0141在形态和培养特征上最相似,而与S.malaysiensis NBRC 16446和S. samsunensis M1463则存在一定差异。
2.1.4 生理生化特征 菌株30702与3株参照菌株均能利用供试的9种常见碳源(表3);菌株30702能够利用丙氨酸、亮氨酸、苯丙氨酸、精氨酸、缬氨酸、甘氨酸和赖氨酸最为惟一氮源,不能利用酪氨酸、酪蛋白和组氨酸作为氮源(表4)。与参照菌株相比,菌株30702与S. solisilvae HNM0141在氮源利用能力上更相近,与其他两株菌存在差异,如菌株30702能够利用亮氨酸和苯丙氨酸,而S.malaysiensis NBRC 16446和S. samsunensis M1463不能,菌株30702能够利用精氨酸和缬氨酸,而S.samsunensis M1463则不能。
菌株30702与所有参照菌株在明胶液化、脲酶实验、硝酸盐还原实验、淀粉水解实验均为阳性(表5),生长温度范围均为20-40℃,生长的酸度范围均为pH5-10。菌株30702在盐度为7%时仍能生长,S. samsunensis M1463和S. solisilvae HNM0141在盐度为5%时均能生长,S. malaysiensis NBRC 16446在盐度为4%时能生长,在5%时不能生长。这说明菌株30702的耐盐性更好。
2.1.5 细胞壁化学分析 菌株30702细胞壁氨基酸成分主要有甘氨酸和L-DAP;细胞壁糖类成分主要为半乳糖。表明菌株30702与链霉菌的细胞壁化学特征相符。
2.2 链霉菌30702的生物防治特性
菌株30702能够产生蛋白酶、纤维素酶、几丁质酶、β-葡聚糖酶和ACC脱氨酶以及产生铁载体,并且具有解磷的能力。同时,菌株30702能够促进拟南芥的生长,在MS培养基上表现为生长茂盛,叶片厚而绿,根毛较长且数目较多(图3-B,3-b)。而对照组叶片薄而发黄,根毛数目少,根短,植物生长弱(图3-A,3-a)。
2.3 菌株30702发酵粗提物抑制炭疽菌孢子萌发和菌丝生长
菌株30702发酵粗提物对山药炭疽菌孢子的萌发和菌丝的生长具有较好的抑制作用。在发酵粗提物浓度为15 μg/mL时,抑制率为(90.33±0.5)%;发酵粗提物浓度为25 μg/m时,抑制率为(90.53±0.8)%;发酵粗提物浓度为50 μg/m时,抑制率为(93.35±0.8)%;发酵粗提物浓度为75 μg/m时,抑制率为(96.82±0.9)%;发酵粗提物浓度为100 μg/m时,抑制率为(99.33±0.6)%。随着浓度的增加,抑制效果越来越显著。在浓度达到100 μg/mL时,抑制率几乎为100%(图4)。
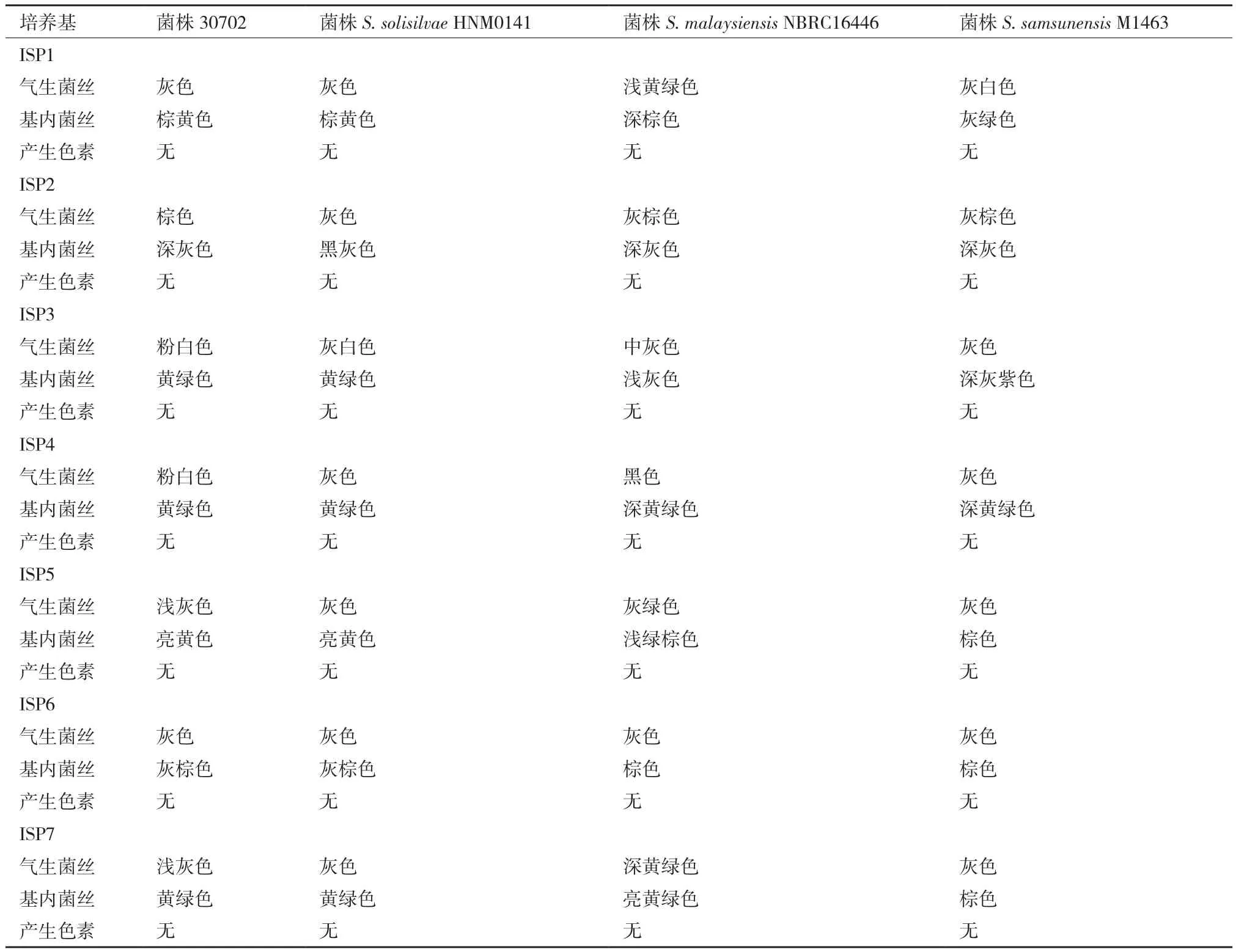
表2 菌株30702与参照菌株培养特征的比较

表3 菌株30702与参照菌株碳源利用试验
3 讨论
链霉菌是一类具有重要应用价值的生防放线菌资源,其分类地位的确认是深入研究其生防功能的前提。16S rRNA基因序列分析是快速确认放线菌分类地位最常用的方法之一[35]。但基因组内的多拷贝性致使16S rRNA 基因不能很好的区分属内密切相关的物种,尤其是链霉菌属同一clade中的种[36]。因此,一些分类学家推荐联合使用多个看家基因串联的多位点序列比较(MLSA)来鉴定链霉菌种的分类地位[36-37]。研究表明链霉菌MLSA的进化距离与DNA-DNA杂交(DDH)之间表现出很强的数据相关性,70%的DDH值匹配性于0.007的MLSA距离[38]。本实验的前期研究中,采用PCR产物直接测序的方法获得了菌株30702的16S rRNA基因部分序列,通过序列比对分析将菌株30702初步鉴定为链霉菌属紫黑链霉菌进化分枝中的放线菌[27],但其种水平的分类地位尚未确定。
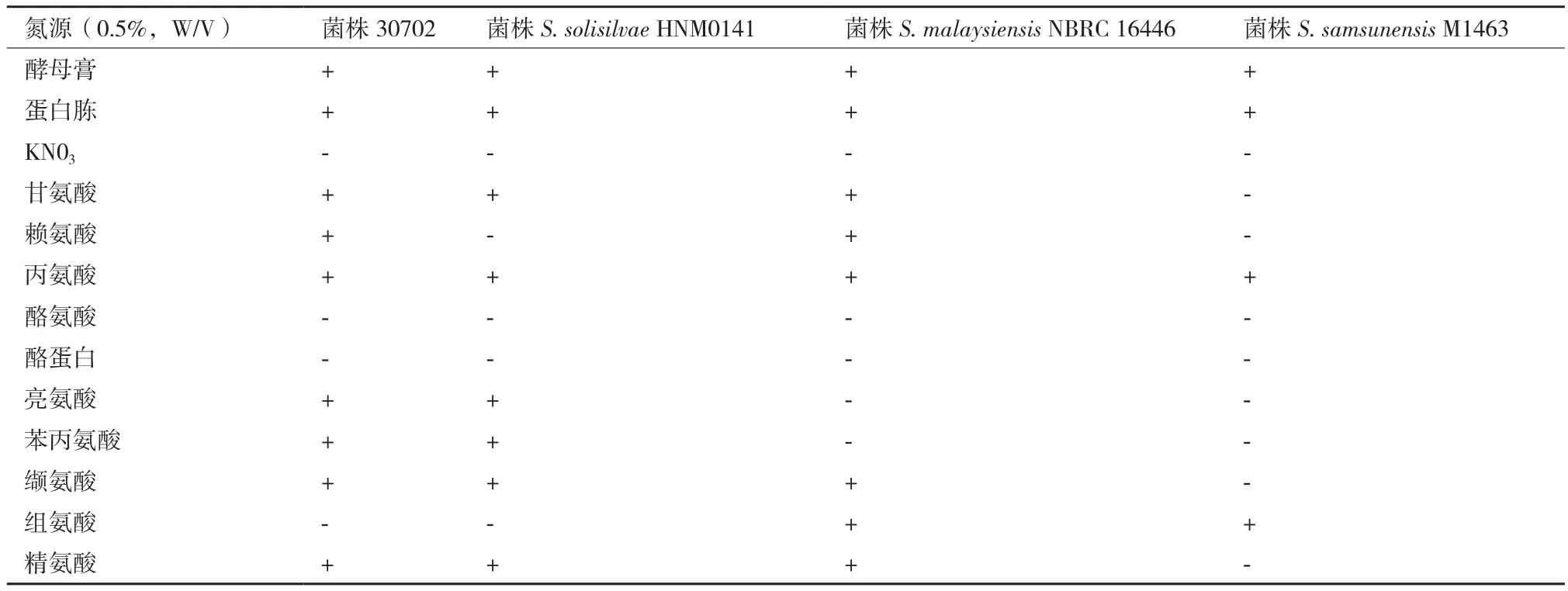
表4 菌株30702与参照菌株氮源利用试验

表5 菌株30702与参照菌株其它生化试验
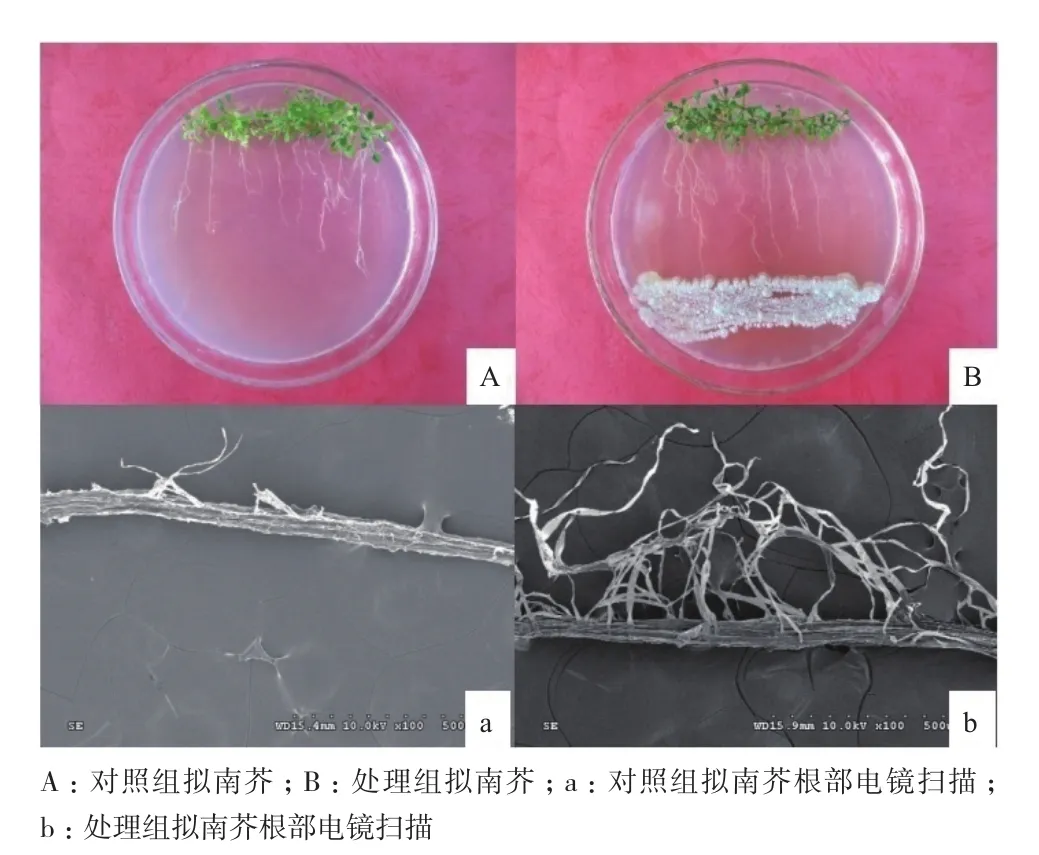
图3 菌株30702体外促进拟南芥的生长
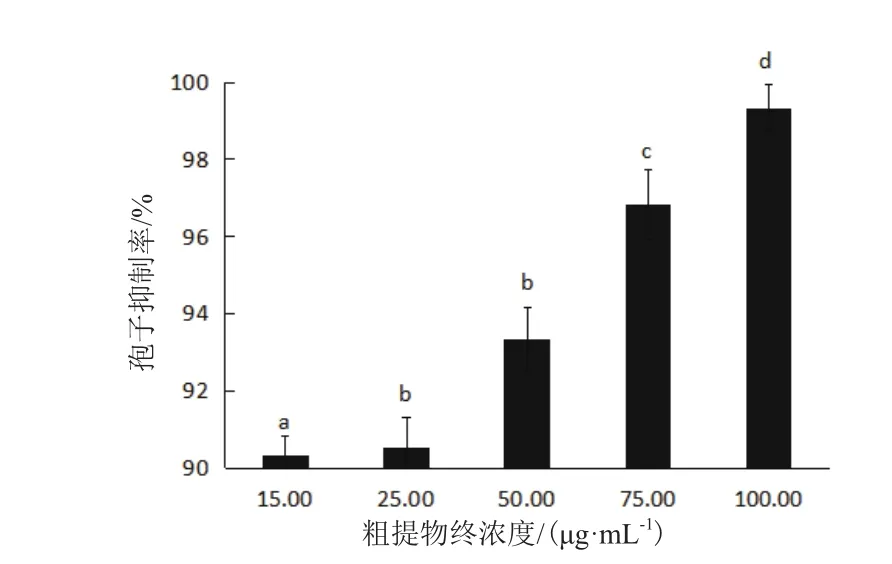
图4 菌株30702发酵粗提物对炭疽菌孢子萌发的影响
本研究通过转化克隆测序的方法获得了菌株30702的16S rRNA基因的全长序列(1481 bp),通过序列比对和16S rRNA基因系统进化树分析,确认 了 Streptomyces solisilvae HNM0141T,Streptomyces malaysiensis NBRC 16446T和Streptomyces samsunensis M1463T为该菌株亲缘关系相对较近的参照菌株。其中Streptomyces solisilvae HNM0141T与菌株30702的亲缘关系最近,相似性达到100%,因此基本推断菌株30702为Streptomyces solisilvae。为了验证这种推断,本研究将菌株30702与参照菌株进行了MLSA多位点序列分析。结果表明菌株30702与S.solisilvae HNM0141的MLSA进化距离小于0.007(种的界限),与其他参照菌株的MLSA进化距离均大于0.007。根据Rong和Huang[38]的观点,菌株30702与S. solisilvae HNM0141理应归属于同一个种,而区别于其他两株参照菌。此外,形态及生理生化特征的比较结果也支持了这种结论。因此,菌株30702最终被鉴定为紫黑链霉菌进化分枝中的Streptomyces solisilvae。
放线菌的生防特性国内外已有一定的报道,主要包括产抗生素、铁载体、水解酶(几丁质酶、β-葡聚糖苷酶、蛋白酶等)和诱导寄主抗病性等[39]。本研究的菌株30702的发酵粗提物能显著抑制山药炭疽菌孢子的萌发和孢子丝的生长,表明该菌株产生了抗真菌作用的抗生素。同时,检测到菌株30702产生蛋白酶、几丁质酶、纤维素酶和β-葡聚糖苷酶等水解酶,这些水解酶据报道都能瓦解丝状病原真菌的细胞壁,从而抑制病菌的生长发育[40]。其次,菌株30702还具有产生铁载体的能力,可以通过捕获铁离子达到抑制植物病原菌生长的目的[40]。此外,菌株30702还具有促植物生长的特性,接种的拟南芥表现为根增长,根毛数目增多,叶片厚而发绿,植株生长茂盛。这可能与菌株30702具有解磷作用和产ACC脱氨酶有关,有报道表明生防细菌通过产生激素、ACC脱氨酶以及解磷作用等改善植物根的发育,达到促进植物生长的目的[41]。因此,上述研究结果综合表明菌株30702具有优良的生防特性,预示其在植物病害的生物防治上具有较好的潜在应用前景。但其抗生素的化学结构、作用机制、发酵工艺、制剂研发、应用方式等尚需进一步的深入研究。
4 结论
本实验室从海南粗榧根际分离到一株链霉菌30702,通过16S rRNA基因序列和多位点序列分析、并结合形态和生理生化特征比较分析,将链霉菌30702鉴定为紫黑链霉菌进化分枝中的Streptomyces solisilvae。该菌株能够产生蛋白酶、纤维素酶、几丁质酶、β-葡聚糖酶、ACC脱氨酶、铁载体,具有解磷活性和促进植物生长的特性,发酵代谢产物能够抑制山药炭疽菌孢子萌发和菌丝生长。
[1]Watve MG, Tickoo R, Jog MM, et al. How many Antibiotics are produced by the genus Streptomyces[J]. Arch Microbiol, 2001,176:386-390.
[2]Goodfellow M, Kumar Y, Labeda DP, et al. The Streptomyces violaceusniger clade:a home for streptomycetes with rugose ornamented spores[J]. Antonie van Leeuwenhoek, 2007, 92 :173-199.
[3]Sembiring L, Ward AC, Goodfellow M. Selective isolation and characterisation of members of the Streptomyces violaceusniger clade associated with the roots of Paraserianthes falcataria[J]. Antonie van Leeuwenhoek, 2000, 78:353-366.
[4]Zhou S, Yang X, Huang D, et al. Streptomyces solisilvae sp. nov.,isolated from tropical forest soil[J]. International Journal of Systematic and Evolutionary Microbiology, 2017, 67(9):3553-3558.
[5]Deboer C, Meulman PA, Wnuk RJ, Peterson DH. Geldanamycin,anew antibiotic[J]. J Antibiot 1970;23:442-447.
[6]Tripathi CKM, Praveen V, Singh V, et al. Production of antibacterial and antifungal metabolites by Streptomyces violaceusniger and media optimization studies for the maximum metabolite production[J].Med Chem Res, 2004;13:790-799.
[7]El-Naggar MY. Kosinostatin, a major secondary metabolite isolated from the culture filtrate of Streptomyces violaceusniger strain HAL64[J]. J Microbiol, 2007, 45:262-267.
[8]Cheng J, Yang SH, Palaniyandi SA, et al. Azalomycin F complex is an antifungal substance produced by Streptomyces malaysiensis MJM1968 isolated from agricultural soil[J]. J Korean Soc Appl Biol Chem, 2010, 53:545-552.
[9]Kang MJ, Strap JL, Crawford DL. Isolation and characterization of potent antifungal strains of the Streptomyces violaceusniger clade active against Candida albicans[J]. J Ind Microbiol Biotechnol,2010, 7:35-41.
[10]Parthasarathi S, Sathya S, Bupesh Get al. Isolation and characterization of antimicrobial compound from marine Streptomyces hygroscopicus BDUS 49. World[J]. J Fish Mar Sci,2012, 4:268-277.
[11]Grahovac J, Grahovac M, Dodić J, et al. Optimization of cultivation medium for enhanced production of antifungal metabolites by Streptomyces hygroscopicus[J]. Crop Prot, 2014, 65 :143-152.
[12]Lam KS, Hesler GA, Mattei JM, et al. Himastatin, a new antitumor antibiotic from Streptomyces hygroscopicus. I. Taxonomy of producing organism, fermentation and biological activity[J]. J Antibiot, 1990, 43:956-960.
[13]Yuan G, Hong K, Lin H, et al. New azalomycin F analogs from mangrove Streptomyces sp. 211726 with activity against microbes and cancer cells[J]. Mar Drugs, 2013, 11 :817-829.
[14]Trejo-Estrada SR, Paszczynski A, Crawford DL. Antibiotics and enzymes produced by the biocontrol agent Streptomyces violaceusniger YCED-9[J]. J Ind Microbiol Biotechnol, 1998,21:81-90.
[15]Chamberlain K, Crawford DL. In vitro and in vivo antagonism of pathogenic turfgrass fungi by Streptomyces hygroscopicus strains YCED9 and WYE53[J]. J Ind Microbiol Biotechnol, 1999, 23 :641-646.
[16]Getha K, Vikineswary S. Antagonistic effects of Streptomyces violaceusniger strain G10 on Fusarium oxysporum f. sp. cubense race 4:indirect evidence for the role of antibiosis in the antagonistic process[J]. J Ind Microbiol Biotechnol, 2002, 28 :303-310.
[17]Getha K, Vikineswary S, Wong WH, et al. Evaluation of Streptomyces sp. strain g10 for suppression of Fusarium wilt and rhizosphere colonization in pot-grown banana plantlets[J]. J Ind Microbiol Biotechnol, 2005, 32:24-32.
[18]Shekhar N, Bhattacharya D, Kumar D, et al. Biocontrol of woodrotting fungi with Streptomyces violaceusniger XL-2[J]. Can J Microbiol, 2006, 52:805-808.
[19]Palaniyandi SA, Yang SH, Suh JW. Foliar application of extract from an azalomycin-producing Streptomyces malaysiensis strain MJM1968 suppresses yam anthracnose caused by Colletotrichum gloeosporioides[J]. J Microbiol Biotechnol, 2016, 26 :1103-1108.
[20]Molinari F, Romano D, Gandolfi R, et al. Newly isolated Streptomyces spp. as enantioselective biocatalysts:hydrolysis of 1,2-O-isopropylidene glycerol racemic esters[J]. J Appl Microbiol,2005, 99:960-967.
[21]Yoshimura S, Otsuka T, Tsurumi Y, et al. WA8242A1, A2 and B, novel secretory phospholipase A2 inhibitors produced by Streptomyces violaceusniger[J]. J Antibiot, 1997, 51 :1-6.
[22]Kino T, Hatanaka H, Hashimoto M, et al. FK- 506, a novel immunosuppressant isolated from a Streptomyces. I. Fermentation,isolation, and physico-chemical and biological characteristics[J].J Antibiot, 1987, 40:1249-1255.
[23]Kumar Y, Aiemsum-ang P, Ward AC, et al. Diversity and geographical distribution of members of the Streptomyces violaceusniger 16S rRNA gene clade detected by clade-specific PCR primers[J]. FEMS Microbiology Ecology, 2007, 62(1):54-63.
[24]Sahin N, Sazak A, Güven K, et al. Diversity of members of the Streptomyces violaceusniger 16S rRNA gene clade in the legumes rhizosphere in Turkey[J]. Annals of Microbiology, 2010, 60(4):669-684.
[25]Kang MJ, Strap JL, Crawford DL. Isolation and characterization of potent antifungal strains of the Streptomyces violaceusniger clade active against Candida albicans[J]. Journal of Industrial Microbiology & Biotechnology, 2010, 37(1):35-41.
[26]黄小龙, 陈吉良, 洪葵, 等. 热带药用植物根际放线菌的分离、鉴定及生物活性分析[J]. 生物技术通报, 2012(2):122-127.
[27]焦敬华, 黄东益, 黄小龙, 等. 山药炭疽病菌拮抗放线菌30702菌株的初步鉴定及发酵培养基优化[J]. 热带作物学报,2016, 3(4):775-783.
[28]周双清, 黄小龙, 黄东益, 等. Chelex-100快速提取放线菌DNA作为PCR扩增模板[J]. 生物技术通报, 2010(2):123-125.
[29]Guo X, Zhang L, Li X, et al. Streptomyces rubrisoli sp. nov.,neutrotolerant acidophilic actinomycetes isolated from red soil[J]. International Journal of Systematic and Evolutionary Microbiology, 2015, 65(9):3103-3108.
[30]Shirling EB, Gottlieb D. Methods for characterization of Streptomyces species[J]. Int J Syst Bacteriol, 1966, 16, 313-340.
[31]Williams ST, Goodfellow M, Alderson G, et al. Numerical classification of Streptomyces and related genera[J].Microbiology, 1983, 129(6):1743-1813.
[32]Lechevalier MP, Lechevalier HA. The chemotaxonomy of actinomycetes //[M]Dietz A, Thayer DW Actinomycete Taxonomy. Arlington, VA:Society of Industrial Microbiology, 1980:227-291.
[33]Palaniyandi SA, Yang SH, Cheng JH, et al. Biological control of anthracnose(Colletotrichum gloeosporioides)in yam by Streptomyces sp. MJM5763[J]. Journal of Applied Microbiology,2011, 111(2):443-455.
[34]Palaniyandi SA, Yang SH, Damodharan K, et al. Genetic and functional characterization of culturable plant-beneficial actinobacteria associated with yam rhizosphere[J]. Journal of Basic Microbiology, 2013, 53(12):985-995.
[35]阮继生、黄英. 放线菌快速鉴定与系统分类[M]. 北京:科学出版社, 2011.
[36]Guo Y, Zheng W, Rong X, et al. A multilocus phylogeny of the Streptomyces griseus 16S rRNA gene clade:use of multilocus sequence analysis for streptomycete systematics[J]. International Journal of Systematic and Evolutionary Microbiology, 2008, 58(1):149-159.
[37]姜钊, 职晓阳, 李文均. 分离自印度洋深海沉积物的部分链霉菌分离株多位点序列分析[J]. 微生物学通报, 2016, 43(5):927-934.
[38]Rong X, Huang Y. Taxonomic evaluation of the Streptomyces griseus clade using multilocus sequence analysis and DNA-DNA hybridization, with proposal to combine 29 species and three subspecies as 11 genomic species[J]. International Journal of Systematic and Evolutionary Microbiology, 2010, 60(3):696-703.
[39]Palaniyandi SA, Yang SH, Zhang L, et al. Effects of actinobacteria on plant disease suppression and growth promotion[J]. Applied Microbiology and Biotechnology, 2013, 97(22):9621-9636.
[40]Pal KK, Gardener BMS. Biological control of plant pathogens[R].The plant health instructor, 2006, PHI-A-2006-1117-02.
[41]Wang S, Wu H, Qiao J, et al. Molecular mechanism of plant growth promotion and induced systemic resistance to tobacco mosaic virus by Bacillus spp.[J]. J Microbiol Biotechnol, 2009, 19(10):1250-1258.
猜你喜欢
中国食品学报(2022年8期)2022-09-07
江苏农业学报(2022年4期)2022-09-07
中国果业信息(2021年5期)2021-12-05
语文周报·教研版(2021年28期)2021-08-19
国际呼吸杂志(2019年22期)2019-12-09
亚热带农业研究(2019年1期)2019-06-11
陕西农业科学(2019年4期)2019-05-13
新疆农业科技(2016年6期)2016-02-19
应用海洋学学报(2014年4期)2014-11-22
塔里木大学学报(2014年1期)2014-06-21
