
麦长管蚜羧酸酯酶基因cDNA片段克隆及吡虫啉胁迫对其表达的影响
2018-06-29 06:47白微微高海峰张航杨安沛李广阔
生物技术通报 2018年6期
白微微 高海峰 张航 杨安沛 李广阔
(新疆农业科学院植物保护研究所 农业部西北荒漠绿洲作物有害生物综合治理重点实验室,乌鲁木齐 830091)
麦长管蚜Sitobion avenae(Fabricius)是新疆麦区的常发性害虫,长期以来主要使用化学杀虫剂进行防治,随着防治次数和用药量的不断上升,麦长管蚜对杀虫剂产生了不同程度的抗药性[1]。昆虫对杀虫剂产生抗性的原因之一是体内解毒酶活性提高,昆虫解毒能力增强[2]。羧酸酯酶是昆虫解毒酶系之一,在昆虫头部、中肠、马氏管等组织中均有存在,参与激素代谢,神经递质降解及农药解毒等[3-4]。随着昆虫抗药性研究的不断深入[5],现已阐明解毒酶参与昆虫抗药性分子机制主要包括基因增强表达与基因内部氨基酸突变两部分[6]。有研究证明棉铃虫Helicoverpa armigera(Hübner)羧酸酯酶的mRNA过量表达对拟除虫菊酯的抗性起重要作用[7];Demaeght[8]等发现二斑叶螨 Tetranychus urticae(Koch)抗螺螨酯品系中一个酯酶基因和两个细胞色素P450氧化酶基因的相对表达量升高;致倦库蚊Culex quinquefasciatus中有机磷类杀虫剂抗性品系的酯酶基因拷贝数量较之敏感品系多达250倍[9];通过RNAi技术证实羧酸酯酶基因参与飞蝗Locusta migratoria manilensis对马拉硫磷等杀虫剂的解毒过程[10-11],这些结果均显示昆虫酯酶在解毒代谢与抗药性中的重要作用。
吡虫啉是蚜虫防治中广泛使用的杀虫剂,具有高效、低毒,与常用杀虫剂无交互抗性的特点,对麦长管蚜有良好防治效果[12]。然而由于吡虫啉长期、频繁的使用,其抗性问题日益引起关注。目前,吡虫啉对麦长管蚜解毒酶基因分子机理层面的影响还尚未明确。结合昆虫解毒酶基因与抗药性的相关研究报道,本研究通过克隆得到羧酸酯酶基因序列,利用实时荧光定量PCR对吡虫啉胁迫下的麦长管蚜羧酸酯酶基因表达进行了初步探索,旨在研究吡虫啉对麦长管蚜羧酸酯酶基因表达的影响,揭示羧酸酯酶基因在麦长管蚜解毒代谢中发挥的作用,为开展麦长管蚜抗性分子机制研究奠定一定基础,同时该研究对于延长现有杀虫剂使用寿命,降低防治成本也具有重要意义。
1 材料与方法
1.1 材料
供试虫源:麦长管蚜采自新疆昌吉军户农场,室内用新鲜麦苗饲养繁殖。饲养条件为:温度(23± 1)℃,光周期14:10(L:D),相对湿度RH 60%-70%。选取大小基本一致的健康无翅成蚜个体为实验材料。
主要试剂:97.4%吡虫啉原药为江苏扬农化工生产;TRNzol Reagent(Total RNA提取试剂)为Invitrogen公司产品,FastQuant cDNA 第一链合成试剂盒、PCR Master Mix、PCR产物纯化试剂盒、SuperReal PreMix(SYBR Green)均为北京天根生化公司产品,pMD 19-T Vector Cloning Kit为Promega公司产品。
1.2 方法
1.2.1 毒力测定及不同浓度吡虫啉对麦长管蚜的处理 吡虫啉毒力测定及后续处理均采用带虫浸叶法,将吡虫啉原药用丙酮稀释为母液,配制成不同浓度梯度,将麦长管蚜试虫连带小麦叶片浸入吡虫啉药液中10 s后取出放于滤纸上晾干,置于放有滤纸的圆形塑料培养皿中,用纱布覆口后将培养皿放于麦长管蚜常规培养箱中,对照试虫使用丙酮处理,24 h后检查并统计蚜虫死亡率(轻触虫体时不动计为死亡),对照死亡率小于10%数据可用。每处理均为3次重复,每重复45-60头蚜虫,利用SPSS 19.0计算毒力回归方程、LC50及95%置信区间。根据毒力测定结果,用吡虫啉LC5、LC15、LC30、LC50和LC80将室内饲养的无翅成蚜按照毒力测定方法处理,24 h后挑取存活成蚜置于-80℃储存备用。
1.2.2 总RNA的提取及第一链cDNA的合成 取40头蚜虫于1.5 mL Eppendorf管中低温匀浆,用TRNzol Reagent 进行总RNA提取后溶于DEPC水,电泳检测核酸质量并测浓度。按照FastQuant cDNA第一链合成试剂盒的试剂说明书合成第一链cDNA,合成cDNA产物置于-20℃储存备用。
1.2.3 麦长管蚜羧酸酯酶基因克隆及序列分析 利用Primer Premier 5.0软件设计引物(表1)。以反转录的第一链cDNA为模板,克隆麦长管蚜羧酸酯酶基因。PCR 扩增体系(50 μL):0.25 μL Ex Taq(5 U/μL),10 μL Ex Taq Buffer,4 μL dNTP Mixture(2.5 mmol/L),cDNA 模板1μL,上、下游引物(10μmol/L)各 0.5 μL,加入 RNase-free water 补足至 50 μL。PCR反应条件为:94℃预变性3 min;94℃变性30 s,55℃退火30 s,72℃延伸30 s,35循环;72℃终延伸10 min。扩增完成后使用1%琼脂糖凝胶电泳检测后将目标产物使用OMEGA的Gel Extraction试剂盒割胶纯化。将纯化后产物与pMD19-T vector连接转化至JM 109感受态细胞中,菌落PCR检测后挑选阳性克隆测序。测序结果及后续分析利用NCBI网站(https://www.ncbi.nlm.nih.gov/)和 DNAMAN 8、ExPASy、MAGA 7.0、GeneDoc等软件进行。
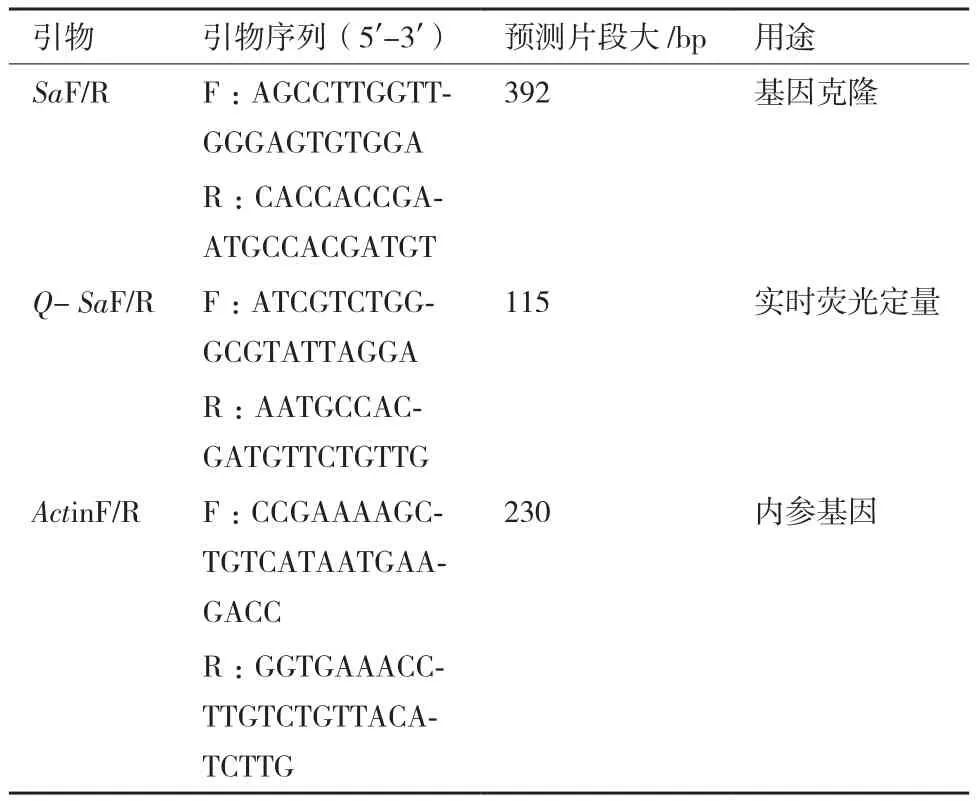
表1 实验引物序列
1.2.4 实时荧光定量PCR(qRT-PCR) 采用qRTPCR分析麦长管蚜羧酸酯酶基因在不同吡虫啉处理浓度下的表达差异,对照为使用丙酮处理后相同条件下饲养的试虫。内参基因Actin[13],所用引物见表1,qRT-PCR 扩增体系(20 μL):8 μL RealMaster Mix,9.8μLRNase-free water,20×SYBR Solution 1 μL,cDNA模板(500 ng/μL)1 μL,上、下游引物(10 μmol/L)各 0.1 μL。反应条件:95℃ 30 s;95℃ 5 s,50℃ 30 s,72℃ 30 s,40个循环。荧光信号采集自动完成,溶解曲线根据荧光定量PCR仪程序进行,每样品3个生物学重复。不同浓度吡虫啉处理下的羧酸酯酶基因相对表达量采用 2-ΔΔCt[14]计算,采用 GraphPad软件对数据多重比较及差异显著性分析,数值均以平均值±SE进行表示。
2 结果
2.1 毒力测定
通过毒力测定结果计算吡虫啉对小麦长管蚜的致死中浓度LC50为13.038 mg/L(95%置信区间为11.951-14.917 mg/L),毒力回归方程为y=-1.148+1.030 x,根据毒力回归方程计算吡虫啉对麦长管蚜的 LC5、LC15、LC30和 LC80值分别为 0.329、1.284、4.035和85.636 mg/L。

图1 麦长管蚜羧酸酯酶SaEST 3(GenBank登录号:KY 441614)推导的氨基酸序列与其他昆虫羧酸酯酶的相似性比较
2.2 羧酸酯酶基因片段的克隆及序列分析
以麦长管蚜cDNA为模板,PCR扩增得到一条约390 bp大小的清晰条带,与预测片段大小基本一致。纯化后连接至pMD 19-T vector中,选取阳性克隆进行序列测定,该序列长度为392 bp,编码130个氨基酸,推导的蛋白质分子量为14 kD,等电点为4.93。将其推导的氨基酸序列与其它昆虫进行序列多重比对,结果(图1)显示其与豌豆长管蚜Acyrthosiphon pisum(GenBank登录号:XP 003248391)、麦双尾蚜Diuraphis noxia(GenBank登录号:XP 015367392)、夹竹桃蚜 Aphis nerii(GenBank登录号:AAF 89552)、桃蚜Myzus persicae(GenBank登录号:P 35502)的酯酶氨基酸序列相似性较高,分别达94%、85%、80%和80%。通过 NCBI 网站Conserved Domain Database保守结构预测(图2)表明,该基因蛋白质序列1-129位均包含在酯酶家族的保守结构域内,属于β酯酶亚族。综合以上结果推测所得片段为麦长管蚜羧酸酯酶基因部分片段,将其命名为SaEST 3,GenBank登录号KY 441614。为进一步明确麦长管SaEST 3进化过程,对和麦长管蚜SaEST 3氨基酸序列相似性较高的昆虫在MEGA 7.0中以N-J算法(Bootstrap 1 000)进行了系统进化树构建(图3),发现SaEST 3与豌豆长管蚜的酯酶亲缘关系最近。
2.3 羧酸酯酶基因的诱导表达
为研究吡虫啉对麦长管蚜羧酸酯酶基因的影响,使用不同浓度吡虫啉(LC5、LC15、LC30、LC50和LC80)处理无翅成蚜24 h后,采用2-ΔΔCt法对羧酸酯酶基因SaEST 3的mRNA相对表达进行处理分析,其中对照组羧酸酯酶基因相对表达量为1。结果(图4)表明吡虫啉对羧酸酯酶基因SaEST 3的表达有一定影响,LC5、LC50、LC80处理下SaEST 3基因相对表达量均达到1.5以上。从LC5到LC80,随着吡虫啉处理浓度增加,SaEST 3基因的转录水平有不同程度的增加,其中吡虫啉LC5处理对羧酸酯酶SaEST 3 mRNA表达的所产生的诱导影响较其它亚致死剂量高,与LC15和LC30的差异显著(P<0.05),与LC50差异不显著(P>0.05)。LC80处理后麦长管蚜在吡虫啉胁迫下羧酸酯酶SaEST 3 mRNA表达水平升高,与LC15和LC30相比较差异显著(P<0.05),与LC5和LC50相比则未达到差异显著性水平(P>0.05)。

图2 麦长管蚜羧酸酯酶核酸序列及推导的氨基酸序列
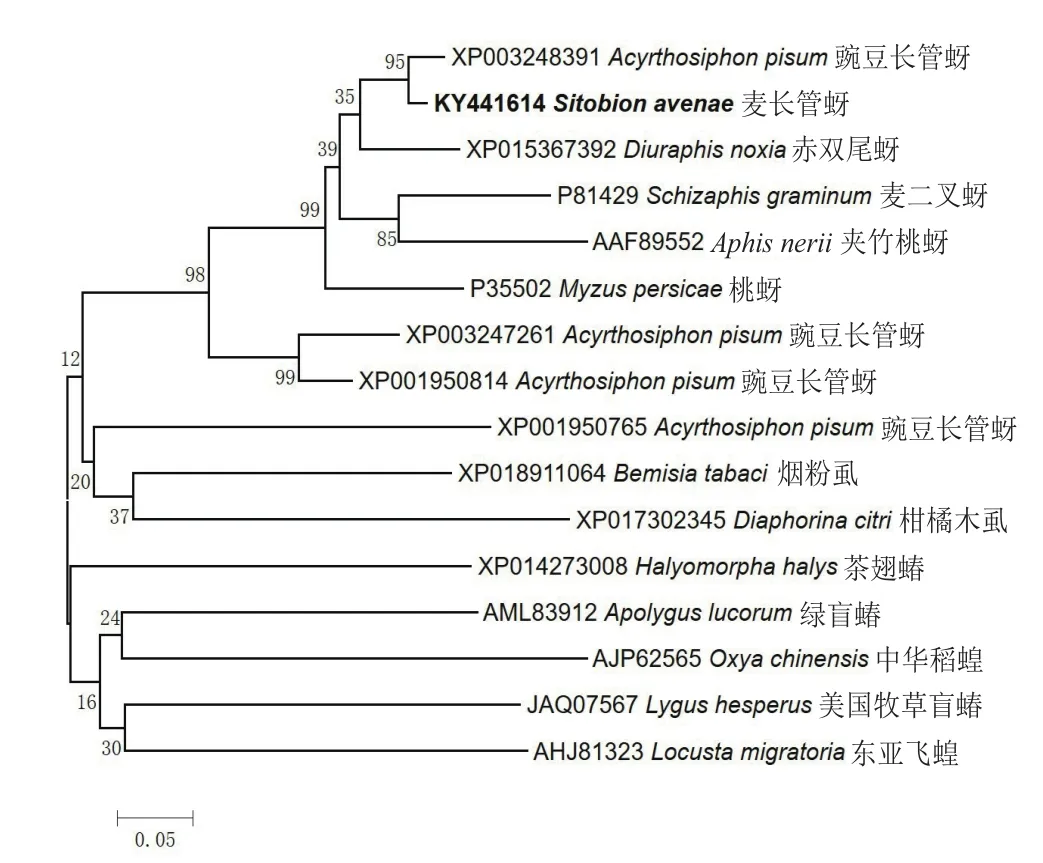
图3 麦长管蚜与其它相关物种羧酸酯酶氨基酸的系统进化树
3 讨论
羧酸酯酶广泛存在与昆虫体内,其对外源性化合物的水解作用增强是昆虫对杀虫剂产生抗性的重要原因之一[15]。目前已通过克隆技术得到多种昆虫的羧酸酯酶基因序列[16-17],本研究利用同源克隆方法得到1个麦长管蚜羧酸酯酶基因片段SaEST 3。通过序列比对和生物信息学分析表明SaEST 3与豌豆长管蚜等昆虫的酯酶有较高相似性,所编码氨基酸与其他几种昆虫酯酶氨基酸序列有较高相似性,定位于酯酶家族的保守区域内,该基因片段为进一步获得麦长管蚜羧酸酯酶基因全长,了解结构功能具有重要意义。
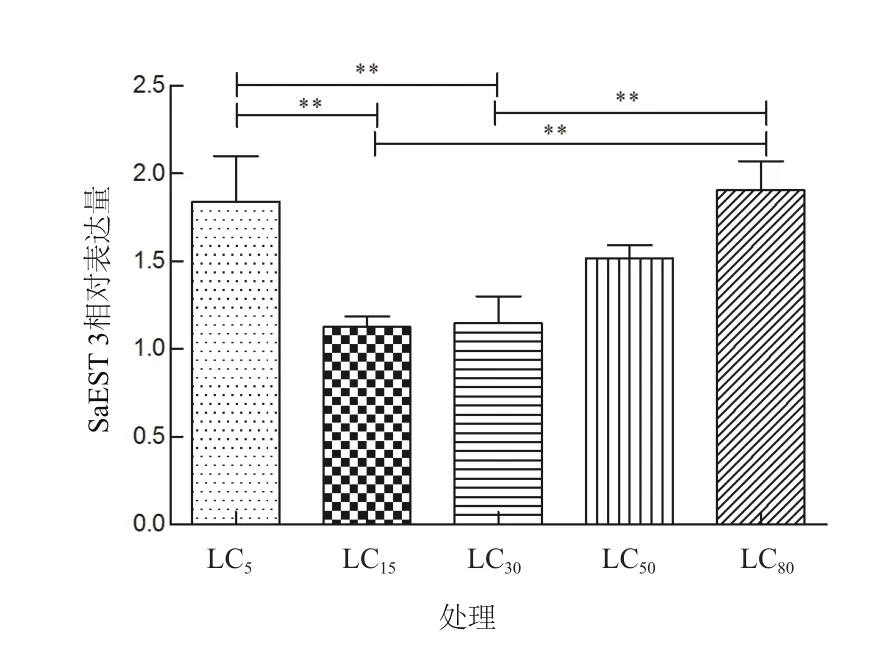
图4 不同浓度吡虫啉处理下麦长管蚜羧酸酯酶基因的表达水平
研究表明羧酸酯酶介导昆虫对有机磷类、氨基甲酸酯类、拟除虫菊酯类及氯虫苯甲酰胺类等多种杀虫剂的抗药发展[18]。羧酸酯酶主要通过提高对杀虫剂的解毒代谢能力、阻隔杀虫剂能力或改变酶对底物亲和力发挥致使昆虫产生抗性的功能[19-20]。用亚致死剂量的氯虫苯甲酰胺连续处理小菜蛾Plutella xyllostella(Linnaeus)5代后处理组羧酸酯酶活性显著高于对照组,表明羧酸酯酶可能参与了小菜蛾对氯虫苯甲酰胺的抗性[21]。解毒酶基因表达量变化跟抗药性密切相关,昆虫解毒代谢酶具有可诱导性,根据这一特征可推测其参与杀虫剂解毒过程的可能性[22],张建琴等[23]对飞蝗经过溴氰菊酯处理后发现羧酸酯酶基因表现为可诱导表达,说明其可能参与飞蝗对溴氰菊酯的代谢解毒和抗性产生,谢佳燕等[24]研究证明吡虫啉不同处理剂量和时间可影响麦二叉蚜Schizaphis graminum(Rondani)启动酯酶蛋白表达,使虫体内酯酶活性有显著性差异,黄水金等[5]发现羧酸酯酶基因在抗性斜纹夜蛾Spodoptera litura(Fabricius)中的转录水平与敏感品系相比高达46.85倍,由此推测羧酸酯酶基因转录水平升高与斜纹夜蛾对溴氰菊酯的抗药性密切相关。
昆虫在药剂胁迫后可能会刺激体内相应的防御解毒体系来不同程度的降低毒害[25]。本研究结果说明吡虫啉对麦长管蚜酯酶基因的表达具有一定影响,据此推测该羧酸酯酶基因SaEST 3可能与麦长管蚜对吡虫啉的解毒代谢有关。羧酸酯酶基因在LC5处理下表达量升高,随着吡虫啉剂量增加呈现先降低后增加的表达水平,表明该基因的相对表达水平受药剂处理浓度的影响,然而对羧酸酯酶基因在吡虫啉处理下的具体表达模式目前并未明确。因此,为了阐明羧酸酯酶基因在麦长管蚜对杀虫剂代谢解毒中发挥的作用,今后的研究工作将围绕羧酸酯酶的序列结构分析和表达模式分析展开,通过对基因结构的进一步探索、转录调控及RNA干扰等技术研究羧酸酯酶的结构特征和验证麦长管蚜羧酸酯酶基因的功能,为探讨麦长管蚜的抗性产生及解毒代谢机制奠定基础。
4 结论
本研究所克隆麦长管蚜羧酸酯酶基因片段大小为392 bp,共编码130个氨基酸,推导的蛋白质分子量为14 kD,等电点为4.93。序列比对和生物信息学分析显示该基因片段与其他几种昆虫酯酶基因有较高相似性,命名为SaEST 3(GenBank登录号:KY 441614)。使用荧光定量技术进一步分析了SaEST3在吡虫啉不同剂量处理下mRNA相对表达水平,结果证明吡虫啉对麦长管蚜羧酸酯酶基因SaEST3表达有一定影响。
[1]张乃芹, 于凌春, 王明友, 等. 小麦蚜虫抗药性现状及综合治理策略[J]. 江西农业学报, 2007, 19(8):50-52.
[2]周斌芬, 唐振华, 高菊芳. 昆虫代谢抗性的研究进展[J]. 农药,2008, 47(5):313-315.
[3]Kontogiannatos D, Michail X, Kourti A. Molecular characterization of an ecdysteroid inducible carboxylesterases with GQSCG motif in the corn borer[J]. J Insect Physiol, 2011, 57(7):1000-1009.
[4]Nardini L, Christian RN, Coetzer N, et al. Detoxification enzymes associated with insecticide resistance in laboratory strains of Anopheles arabiensis of different geographic origin[J]. Parasites&Vectors, 2012, 5:113.
[5]黄水金, 秦文婧, 陈琼. 斜纹夜蛾羧酸酯酶基因的克隆、序列分析及表达水平[J]. 昆虫学报, 2010, 53(1):29-37.
[6]李飞, 黄水金, 韩召军. 害虫抗药性分子检测技术[J]. 生命的化学, 2003, 23(5), 392-395.
[7]袁国瑞. 小菜蛾GABA受体基因克隆及棉铃虫羧酸酯酶的功能表达[D]. 南京:南京农业大学, 2011.
[8]Demaeght P, Dermauw W, Tsakireli D, et al. Molecular analysis of resistance to acaricidal spirocyclic tetronic acids in Tetranychus urticae:CYP392E10 metabolizes spirodiclofen, but not its corresponding enol[J]. Insect Biochem Mol Biol, 2013, 43(6):544-554.
[9]Claude M, Pasteur N, Berge JB, et al. Amplification of an esterase gene is responsible for insecticide resistance in a California Culex Mosquito[J]. Science, 1986, 233(4765):778-780.
[10]Zhang JZ, Zhang JQ, Yang ML, et al. Genomics-based approaches to screening carboxylesterase genes potentially involved in malathion resistance in oriental migratory locust(Locusta migratoria manilensis[J]. Pest Manag Sci, 2011, 67(2):183-190.
[11]Zhang JQ, Li DQ, Ge PT, et al. RNA interference revealed the roles of two carboxylesterase genes in insecticide detoxification in Locusta migratoria[J]. Chemosphere, 2013, 93(6):1207-1215.
[12]刘爱芝, 贾艳丽, 韩松, 等. 高毒替代农药对小麦蚜虫的防治效果[J]. 河南农业科学, 2007, (11):58-60.
[13]Zhang M, Zhou YW, Wang H, et al. Identifying potential RNAi targets in grain aphid(Sitobion avenae F.)besed on transcriptome profiling of its alimentary canal after feeding on wheat plants[J].BMC Genomics, 2013, 14:560.
[14]Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR[J]. Nucleic Acids Res, 2001, 29(9):2002-2007.
[15]Cygler M, Schrag J D, Sussman J L, et al. Relationship between sequence conservation and three-dimensional structure in a large family of esterases, lipases, and related proteins[J]. Protein Science, 2008, 2(3):366-382.
[16]Field LM, Williamson MS, Moores GD, et al. Cloning and analysis of the esterase genes conferring insecticide resistance in the peachpotato aphid, Myzus persicae(Sulzer)[J]. Biochemical Journal,1993, 294(Pt2)(2):569-74.
[17]陈茂华, 蓝家样, 韩召军, 等. 禾谷缢管蚜羧酸酯酶基因cDNA片段的克隆与序列分析[J]. 麦类作物学报, 2006, 26(4):145-148.
[18]任娜娜, 谢苗, 尤燕春, 等. 羧酸酯酶及其介导昆虫抗药性的研究进展[J]. 福建农林大学学报:自然科学版, 2014, 43(4):337-344.
[19]唐茂华. 昆虫抗药性及其治理[M]. 北京:农业出版社,1993.
[20]Ranson H, N’guessan R, Lines J, et al. Pyrethroid resistance in African anopheline mosquitoes:what are the implications for malaria control?[J]. Trends Parasitol, 2011, 27(2):91-98.
[21]邢静, 梁沛, 高希武. 亚致死浓度氯虫苯甲酰胺对小菜蛾药剂敏感度和解毒酶活性的影响[J]. 农药学学报, 2011, 13(5):464-470.
[22]Poupardin R, Reynaud S, Strode C, et al. Cross-induction of detoxification genes by environmental xenobiotics and insecticides in the mosquito Aedes aegypti:Impact on larval tolerance to chemical insecticides[J]. Insect Biochem Mol Biol, 2008, 38(5):540-551.
[23]张建琴, 王燕, 李大琪, 等. 溴氰菊酯对飞蝗羧酸酯酶基因表达的影响[J]. 应用昆虫学报, 2014, 51(2):418-424.
[24]谢佳燕, 林佳. 吡虫啉胁迫时间和剂量对麦二叉蚜酯酶活性的影响[J]. 江苏农业科学, 2015, 43(3):116-118.
[25]Chevillon CH, Raymond MI, Guillemaud TH, et al. Population genetics of insecticide resistance in the mosquito Culex pipiens[J].Biological Journal of the Linnean Society, 1999, 68(1/2):147-157.
猜你喜欢
河北果树(2021年4期)2021-12-02
建材发展导向(2021年7期)2021-07-16
四川蚕业(2021年4期)2021-03-08
建材发展导向(2021年24期)2021-02-12
核农学报(2021年2期)2021-01-28
世界热带农业信息(2018年3期)2018-09-26
天然产物研究与开发(2018年8期)2018-09-10
天然产物研究与开发(2018年4期)2018-05-07
分析化学(2017年12期)2017-12-25
中国塑料(2015年2期)2015-10-14
