
不同遮阴条件下麦冬光合和叶绿素荧光特性研究
2018-06-29 06:47刘丽娟高辉
生物技术通报 2018年6期
刘丽娟 高辉,2
(1. 西华师范大学环境科学与工程学院,南充 637002;2. 西藏大学农牧学院,林芝 860000)
麦 冬(Ophiopogon japonicus(L.F.)Ker-Gawl)是百合科沿阶草属多年生常绿草本植物,又名沿阶草、书带草、麦门冬,在我国很多地区都有野生分布和栽培种植,这些年常常被用作园林绿化植物,具有养阴生津、润肺清心等功效,主治肺燥干咳,津伤口渴等症[1]。目前对于麦冬的研究多集中在繁殖栽培技术和耐阴性的研究[1-2],对其光合生理特性方面的研究报道较少[3]。植物的光合作用是其能量吸收固定、物质分配转化、水碳循环等重要的生理生态和复杂的生物化学过程,是所有物质和能量代谢的物质基础,同化合成作物中90%-95% 的有机物质,与植物的生长密切相关[4]。光合作用除了受植物本身的遗传特性影响外,还受包括湿度、温度、光照强度、二氧化碳浓度和矿物质营养等诸多环境因素影响,其中光照强度是调节植物生长发育的一项基本措施,不同的遮阴条件会直接影响植物的光照强度,还会影响植株的生理代谢,使光合作用发生相应的变化,进而影响植物的生长情况[5]。本研究通过对麦冬植株进行不同的遮阴处理,并且采用Li-6400XT便携式光合分析仪拟探其光合和生理作用,探究最优的种植栽培方案,为其大规模栽培条件的探索提供理论依据。
1 材料与方法
1.1 材料
西 华 师 范 大 学 实 验 地(106°03′46.7′′E,30°48′54.1′′N),海拔 288 m。土壤以紫色土为主[6],供土的基本理化性质具体见表1。属亚热带季风湿润气候区,年均温度15.8℃-17.8℃,年降水量在1 020-1 250 mm,年均相对湿度78%-85%,多年平均日照仅 152.73 h[6-8]。

表1 供试土壤基本理化性状
试验材料采集于2016年9月,采集地为四川省南充市西山风景区,采集后移栽至西华师范大学实验地内。驯化3个月后,2016年12月,选择生长良好、大小基本一致的麦冬盆栽苗,利用不同层数的遮阳网对其进行遮阴处理:共设5个处理,控制相对光强为全光照的95%、72%、48%、24%和8%。
1.2 方法
每种遮阴处理选择60株,待遮阴驯化6个月左右,2017年7月取受光方向一致,且发育时期相同的叶片测定各项指标,每一指标重复5次。
1.2.1 叶绿素含量的测定 分光光度法:利用分光光度计测定叶绿素提取液在最大吸收波长下的吸光值,即可用朗伯—比尔定律计算出提取液中各色素的含量。叶绿素a和叶绿素b在645和663 nm处有最大吸收,且两吸收曲线相交于652 nm处。因此测定提取液在645、663和652 nm波长的吸光值,并根据经验公式分别计算出叶绿素a和叶绿素b的量。
1.2.2 光响应曲线的测定 用Li-6400XT便携式光合仪,叶室选取红蓝光源,空气CO2浓度设定为恒定400 μmol/mol,温度为25℃,光合有效辐射(Photosynthetically available radiation)从最大值(2 000 μmol/m2s)逐渐降低至最小值(0 μmol/m2s),测定在不同光合有效辐射下的麦冬叶片的净光合速率,绘制光响应曲线。非直角双曲线拟合,根据拟合曲线计算光饱和点和光补偿点[5]。非直角双曲线公式为:
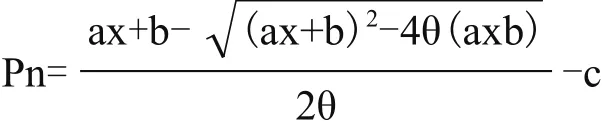
其中,Pn为净光合,x为光强,c为暗呼吸强度,b为最大光合速率,a为直线部分的斜率,θ为曲线形状参数。重复5次。
1.2.3 CO2响应曲线的测定 采用Li-6400XT CO2响应自动曲线测定,光源设定为1 200 μmol/m2s,温度为25℃,将CO2浓度从较大浓度(1 000 μmol/mol)逐渐减少(20 μmol/mol),根据非直角双曲线拟合,计算出CO2补偿点、CO2饱和点、最大净光合效率等重要参数;重复5次。
1.2.4 光合速率日变化的测定 选择晴天,从8:00到20:00,每隔2 h测定自然状态下的麦冬叶片光合速率与蒸腾速率,共测定5株。
1.2.5 荧光参数的测定 采用英国Technologica公司的CF Imager叶绿素荧光仪测定各种处理麦冬植株分成的叶绿素荧光参数。并计算PSⅡ最大光能转换效率(Fv/Fm)作用光下实际的PSⅡ光化学效率ΦPSⅡ;PSⅡ有效光化学量子效率(Fv′/Fm′)、电子传递速率(Electron transport rate,ETR)、光化学淬灭系数(Photochemical quenching,qP),非光化学淬灭系数(Non-photochemical quenching,NPQ)。叶绿素荧光参数Fv/Fm、Fv/Fo是研究植物生理生化的重要参数指标,Fv/Fm是指开放的PSⅡ反映中心捕获激发能的效率,即内禀光化学效率。光反应中心PSⅡ的潜在活性主要用Fv/Fo来表示。遮阴后能够激发麦冬叶片的光能利用效率,不过当弱光达到一定程度时,会对PSⅡ的功能造成一定的影响,使PSⅡ实际的光能利用效率、原初光能捕获效率以及光合电子传递效率下降。qP反映PSⅡ反应中心中开放程度,1-qP则反映了反应中心关闭程度,反映了 QA 的 还 原 程 度。qP=(Fm′-Fs)/(Fm′-F0′);NPQ可以用来表示植物热耗散的能力变化,一般情 况 下 数 值 范 围 为 0-n(1,2,3,4,5……)。NPQ=(Fm-Fm′)/Fm′=Fm/Fm′-1;qN 与 NPQ 相同,现在使用较少。数值范围为0-1。qN=(Fm-Fm′)/(Fm-F0′);ETR =PPFD×ΦPS Ⅱ ×0.84×0.5。
2 结果
2.1 遮阴对麦冬叶绿素含量的影响
由表2可知,麦冬在不同遮阴处理下叶绿素含量不同,表现在先增加后减小的趋势,48%遮阴处理和72%遮阴处理差异不显著,当遮阴度达到95%时叶绿素比例差异显著,表明光照太强或者太弱对麦冬叶绿素含量影响较大。也间接说明麦冬在是当遮阴处理下可以更好的利用光能进行光合作用,更适合在弱光的环境下生长。
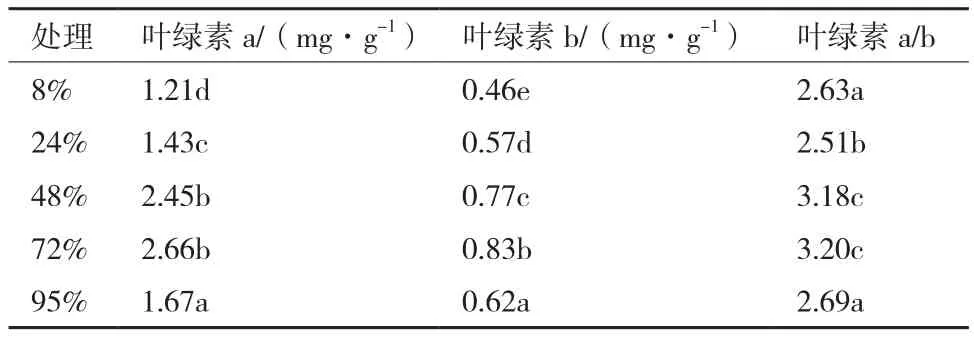
表2 不同遮阴下麦冬叶片叶绿素含量及比例
2.2 光强对麦冬光合参数的影响
麦冬饱和点和光补偿点都比较低,适当的遮阴处理对暗呼吸速率(Dark respiration rate)、最大净光合速率(Maximum net photosynthetic rate)、光饱和点(Light saturation point,LSP)和光补偿点(Light compensation point,LCP)都有明显的影响(表3)。
麦冬的LCP在48%光照强度下呈现最大值,在24%光照处理下呈现最小值不同光照处理下差异显著,整体看来呈现先增大后减小的趋势;光饱和点在48%光照处理下表现出最大值;但表观量子效率(Apparent quantum efficiency,AQE)随着光照强度的增大表现出先升高后降低。暗呼吸速率和最大净光合速率均呈现出先降低后增高的特点。反映麦冬在相对比较弱的光照强度下,能够大幅度的提高光饱和点,降低光补偿点,从而最大程度利用弱光,降低自身的呼吸消耗,用来保证植株的正常生长,说明麦冬适合在遮阴环境下生长。
为了进一步探讨麦冬对弱光的利用能力,对不同处理下的麦冬植株的净光合速率、气孔导度、胞间CO2浓度、蒸腾速率和水分利用效率作了多元相关性分析(表4)。发现净光合速率随着光照强度的升高出现先降低后升高的趋势,胞间CO2浓度在不同处理下基本恒定;蒸腾速率先降低后升高,对水分的利用能力表现出先增大后减小的趋势。

表3 不同遮荫处理对麦冬光合参数的影响
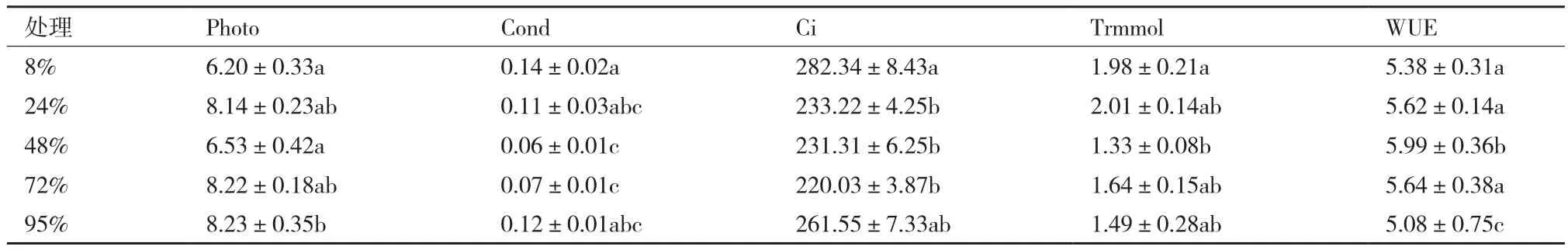
表4 不同遮阴处理对麦冬叶片光合参数的影响
2.3 遮阴对麦冬荧光参数的影响
随相对光强的升高,Fv/Fm逐渐降低,与对照组相比,差异比较明显;ΦPSⅡ、ETR和qP均在72%遮荫处理下的值最大,在8%处理下最小;Fv′/Fm′试验组的值均比对照组全光照条件下大(表5)。随光强的减弱,相对光强8%-24%时,PS以热耗散形式消耗的光能增加,而用于光合电子传递的光能减少,qP先升高后降低,证明在相对光强为48%-72%时,PSⅡ用于光合电子传递的光能增加,而以热耗散形式消耗的光能减少。
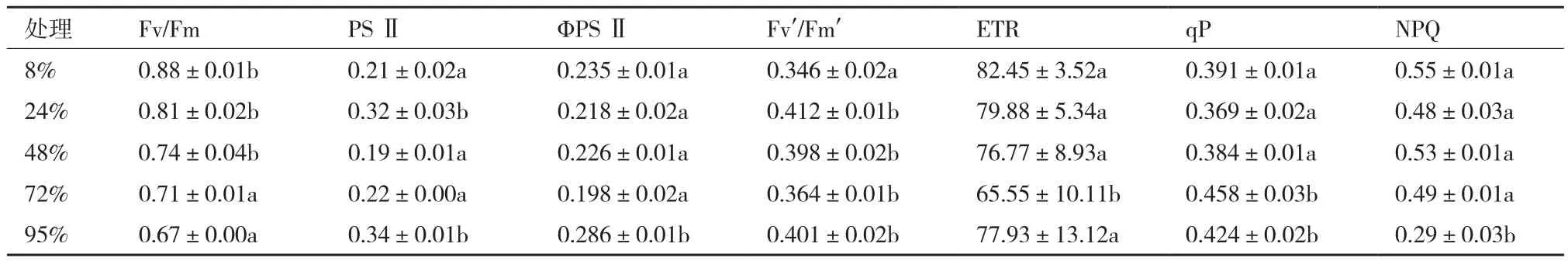
表5 不同遮阴对麦冬叶片荧光参数的影响
2.4 光合速率和蒸腾速率日变化
从8点开始,随着光照强度的增强,麦冬的净光合速率迅速增强(图1)。在阳光充足的条件下,不同遮阴处理下的麦冬净光合速率(Pn)的日变化呈现明显的差异。从图1可以看出麦冬光合速率呈现出明显的双峰曲线;随着时间的增长,麦冬净光合速率在12:00点附近出现第一个极峰,随后出现下降趋势,在14:00附近出现低值(峰谷),根据生态学原理可知麦冬植株应该是出现了植物特有的“午休情况”。在下午15:30左右,麦冬的净光合速率出现第二次峰值,随后一直降至1点附近。从图1可以看出麦冬的净光合速率在72%的试验控制条件下可以达到最大值,8%光照条件下净光合速率值较低,说明适当遮阴理有利于麦冬的光合作用,有利于有机物质的积累和合成。麦冬在全天各个时间的光合速率与光合有效辐射及气温变化相关性不大。

图1 不同遮荫条件下麦冬净光合速率(Pn)日变化
3 讨论
遮阴条件是麦冬生长的主要限制因子,适当遮阴或者合适的种植密度可以很好地改善麦冬群体结构,可以提高对光能的利用率。而光照不足则会限制麦冬的光合作用,导致碳获取的减少,使生长受阻。因此植物光合作用的效率决定着其生产力水平,其光合性能可以通过叶绿素含量的比例变化、光合速率得到衡量[7-8],而叶绿素荧光作为光合作用机理研究的有力探针,可以更直接地反映不同光照条件下麦冬植株光合作用反应中心的开放程度及植物热耗散等情况,是一种反应灵敏、操作简便且对植物无伤害的方法。遮阴对光合作用的影响,因作物品种、生育期和遮阴时间的不同,会有很大差异,许多研究表明,遮阴通常可以降低单叶的净光合速率[9]。
植物的耐阴性一般情况下可以用植物光补偿点来衡量。植物的光补偿点越低,在弱光条件下光合作用能力就越强,植物的有机物质开始迅速积累;光饱和点越低,植物光合作用速率随光量子通量的增大就可以很快达到最大效率。麦冬的光补偿点在适度遮阴的条件下可以适当降低,说明它对光强辐射强度要求的下限降低,并且通过增加表观量子效率来适应外界光强的降低,对弱光具有一定的适应能力[10]。也可以直接证明植物在外界环境改变的时候,可以通过自身的调节来使自己尽快的适应下逆境环境。当光强降至自然光的24%以下时,麦冬植株的表观量子效率开始下降,表明光合体系已经不能够适应更弱的光照环境,许多试验表明,遮阴条件可以降低植物单叶的净光合速率[11-12]。
植物在一定范围内对光环境有很强的自我适应与调节能力,一些植物在弱光下会通过合成大量的叶绿素,从而去捕获更多的光能,但不同光反应类型的品种之间也会存在差异[13-14]。本试验研究结果与梁镇林等[15]的研究结果一致,即随着遮阴度的增加,麦冬叶片的叶绿素含量均显著增加。光是影响叶绿素形成的主要条件,从原叶绿素酸酯转变为叶绿酸酯需要光,而光过强,叶绿素又会受光氧化而破坏。黑暗中生长的幼苗呈黄白色,遮光或埋在土中的茎叶也呈黄白色,这种因缺乏某些条件而影响叶绿素形成,使叶子发黄的现象。
麦冬叶片的荧光参数Fv /Fm随光强的减弱而增加,但是 ΦPSⅡ、Fv ′/Fm ′、ETR和 qP在光强为72%光照处理条件下可以达到最大值,反映出在一定的弱光范围内ΦPSⅡ的光合能力提高,且能够将吸收的光能更多的分配给光化学途径。从试验数据看出当光强小于24%时,ΦPSⅡ等参数开始下降,证明上述引起光合速率降低的次要因素即ΦPSⅡ的光合活性并非未受到影响,这与迟伟等对草莓的研究结果相吻合,也可能与光合色素含量的降低有关[14]。由于在阴生的环境下的麦冬植株,需要对植株中保障能够正常进行生长的叶绿素荧光参数进行维持,从而会捕获大量的光量子来支撑植株的光合作用,并且可以大幅的降低呼吸消耗。Fv/Fm也就是所说的内禀光化学效率,主要指开放回路中的PSⅡ反应中心所捕获激发能进行的效率。Fv/Fo表示光反应中心PSⅡ的潜在活性。一般对于植物来说,最大的光能转化效率并不代表着最大的光合速率。PSⅡ反应中心可以将天线色素吸收的过量的光能以热耗散的形式消耗从而避免对光合机构造成失活或破坏。
随着光照强度的增加,Pn数值显著增强,但是光照达到一定程度会对植物造成伤害[17],导致光合作用中氧化副产物增加。这些终产物不是植物生长发育必需的。所以在光合生理上表现为ETR、Y、qP和Pn数值的降低(说明PSⅡ活性降低),而qN值增高[18],会引起光合作用的光抑制,过剩的能量会形成各种活性氧分子,最终可能导致膜质过氧化,植物体光合色素分解即产生光氧化伤害,这种伤害是不可逆的,此时叶子形态会发生病变。光抑制同时也会降低植物的光合生产力,限制植物在自然界的适应和生长。
4 结论
麦冬为阴生植物,适当的遮阴处理有利于麦冬叶绿素的积累,栽培麦冬植株的过程中应调整种植密度或者进行遮阳网遮阴处理,使相对光强控制在自然光的72%左右为宜。
[1]韩蕾, 李成, 赵五一, 等. 山麦冬繁殖与栽培[J]. 林业实用技术, 2007, 8(6):43-57.
[2]汪发缵, 唐进. 中国植物志(第十五卷)[M]. 北京:科学出版社,1978.
[3]王晓玲, 石雷孙, 吉雄. 遮荫对山麦冬生长特性和生物量分配的影响[J]. 植物研究, 2002, 12(2):225-228.
[4]张文标, 金则新, 柯世省, 等. 木荷光合特性日变化及其与环境因子相关性分析[J]. 广西植物, 2006, 26(5):492-498.
[5]Arvidsson J. Nutrient uptake and growth of barley as affected bysoil compaction[J]. Plant and Soil, 1999, 208(1):9-19.
[6]段浩, 杨军, 苏智先. 等. 南充市水土流失成因分析及防治对策[J]. 灾害学 , 2004, 19(3):34-37.
[7]Hu YB, Sun GY, Wang XC. Induction characteristics and response of photos-ynthetic quantum to changes in ir-radiance in mulberry plants[J]. Jouranl of Plant Physiology, 2006, 164(5):959-968.
[8]闫瑞瑞, 卫智军, 运向军, 等. 放牧制度对短花针茅荒漠草原主要植物种光合特性日变化影响的研究[J]. 草业学报, 2009,18(5):160-167.
[9]刘贤赵, 康绍忠, 周吉福. 遮阴对作物生长影响的研究进展[J].干旱地区农业研究, 2001, 19(4):65-73.
[10]周兴元, 曹福亮. 遮荫对假俭草抗氧化酶系统及光合作用的影响[J]. 南京林业大学学报:自然科学版, 2006, 30(3):35-43.
[11]张往祥, 曹福亮, 吴家胜, 等. 遮荫对银杏光合性能及其叶片产量的影响[J]. 南京林业大学学报, 2000, 24(4):11-18.
[12]Gregoriou K, Pontikis K, Vemmos S. Effects of reduced irradi2 ance on leafmorphology, photosynthetic capacity, and fruit yieldin olive(Olea europaea L.)[J]. Photosynthetica, 2007, 45(2):172-181.
[13]Farquhar GD, Sharkey TD. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Physiology, 1982, 33(6):317-326.
[14]许岳飞, 金晶炜, 陈浩, 等. 草坪植物耐弱光机理研究进展[J].草地学报, 2011, 19(6):1064-1069.
[15]梁镇林, 梁慕勤, 潘世元, 等. 大豆耐阴性研究X:不同耐阴性大豆叶片叶绿素含量和比叶重研究[J]. 贵州农学院学报,1992, 11(2):16-22.
[16]张守仁. 叶绿素荧光动力学参数的意义及讨论[J]. 植物学通报, 1999, 16(4):444-457.
[17]Melis A. Photosystem Ⅱ damage and repair cycle in chloroplasts:What modulates the rate of photo damage in vivo[J]. Trends in Plant Science, 1999, 21(4):130-136.
[18]Dai YJ, Shen ZG, Liu Y, et al. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence and chlorophyll content of Tetrastigma hemsleyanum Diels et Gilg[J].Environmental and Experimental Botany, 2009, 65(2):177-182.
猜你喜欢
江苏农业学报(2018年6期)2019-01-04
特别健康(2018年4期)2018-07-03
现代园艺(2017年23期)2018-01-18
新课程·下旬(2017年7期)2017-08-14
中成药(2017年4期)2017-05-17
中成药(2016年4期)2016-05-17
动物营养学报(2015年10期)2015-12-01
西北林学院学报(2015年5期)2015-01-03
中国中医药现代远程教育(2014年16期)2014-03-01
植物营养与肥料学报(2013年1期)2013-10-26
