
虚拟仿真场景中威胁性视觉刺激搜索的注意偏向效应*
2018-06-22 01:49:10袁小钧崔晓霞曹正操汪亚珉
心理学报 2018年6期
袁小钧 崔晓霞 曹正操 阚 红 王 晓 汪亚珉
(首都师范大学心理学院, 北京市“学习与认知”重点实验室, 北京 100048)
1 引言
威胁性刺激引起注意偏向一直备受心理学家关注。根据注意的认知资源理论, 认知系统内存在着一个负责资源分配的机制, 这一机制是受意识控制的, 可以把认知资源灵活地分配到重要的刺激上。同时, 人类的认知系统内还存在着一个评价环境重要性的系统, 重要的刺激会优先进入到这个系统, 获得进一步加工(彭聃龄, 2012)。因此, 这些占有认知资源比较多、优先加工的刺激会引起注意偏向。
进化心理学认为, 对威胁性刺激的偏向加工是进化而来的。人类已经进化了对远古危险物的害怕倾向, 形成了一些生存适应器, 这些适应器会处理危及我们祖先生命的各种问题。当我们在一个开阔的地方四处张望时, 我们的信息加工机制会让我们觉察到“暗藏的敌人”, 我们的注意会自动地偏向这些危险物。与现代环境中的危险物相比, 人们更容易对远古环境中就存在的危险物表现出威胁反应。比如说, 人大多害怕蛇, 但很少害怕汽车, 尽管汽车每年夺走很多人的生命。对此, 进化心理学的解释是蛇是人类进化过程中一直存在的危险物(Buss,1999)。这种威胁不仅反应在情绪上, 还反映在注意偏向上。因此, 威胁性刺激更容易引起注意偏向。
在已有的威胁性刺激导致注意偏向的研究中,最常用的研究范式主要有 5种:视觉搜索范式(Öhman, Flykt, & Esteves, 2001)、情绪 Stroop 范式(Janczyk, Ausgst, & Kunde, 2014)、线索−靶子范式(Stormark, Nordly, & Hugdahl, 1995)、点探测范式(Mogg, Bradley, & Hallowell, 1994)、情绪条件反射范式(Stormark & Hugdahl, 1996), 其中视觉搜索范式是使用最为广泛的一种范式。在早期 Öhman和Flykt等(2001)的威胁性刺激搜索实验中, 研究者以蛇、蜘蛛、蘑菇、花的图片作为实验材料, 要求被试在一些与威胁无关的图片(花、蘑菇)中搜索与威胁相关的图片(蛇、蜘蛛), 或在一些与威胁相关的图片(蛇、蜘蛛)中搜索与威胁无关的图片(花、蘑菇)。结果发现在与威胁无关物体中搜索与威胁相关的目标的速度要显著快于在与威胁相关的物体中搜索与威胁无关的目标, 并且不受目标物所在位置以及分心物数目的影响。此后, Mogg和Bradley(2006)使用视觉搜索范式研究了不同高低水平蜘蛛威胁在不同暴露时间下的注意偏向。实验时给被试成对呈现蜘蛛和猫的图片, 呈现时间分 3种, 分别为200 ms、500 ms、2000 ms。研究结果发现, 当呈现时间为 200 ms时, 高水平蜘蛛威胁组对蜘蛛的注意偏向比低水平组要大。随着暴露时间的增加,高水平组注意偏向降低。在更长暴露时间时, 这种注意偏向差异则不再显著。这一研究表明, 威胁性刺激对注意的捕获效应主要发生在早期的快速视觉信息加工阶段。王福兴、李文静、颜志强、段朝辉和李卉(2015)使用眼动技术来研究幼儿对威胁性刺激物——蛇的注意偏向, 结果也发现, 无论是幼儿还是成人对蛇的首次注视时间都比注视花朵的时间更短。LoBue (2010)使用视觉搜索范式研究了儿童和成人对蜘蛛的注意偏向。该实验要求被试在蘑菇或蟑螂的图片中搜索蜘蛛, 或者是反过来进行搜索。研究结果表明, 无论是儿童还是成人, 搜索蜘蛛的速度都比其他两种要快, 并且搜索蘑菇和蟑螂差异不显著, 后续的电生理研究也得到了类似的结果(LoBue & Rakison, 2013; Thrasher & LoBue,2016)。Dudeney, Sharpe和Hunt (2015)对焦虑儿童的研究也发现, 与正常儿童相比, 焦虑儿童在注意搜索中能更快地搜索到威胁性刺激。
除了蛇与蜘蛛外, 负性情绪面孔也常被用作威胁性刺激, 研究者们以面孔为刺激材料的研究得到了同样的结论。Hansen和Hansen (1988)最早以面孔图片为刺激材料来研究情绪刺激的注意偏向。该实验要求被试在一个2 × 2的矩阵中搜索面孔, 结果表明, 被试在快乐面孔中搜索愤怒面孔较快, 而在愤怒面孔里搜索快乐面孔较慢。他认为愤怒面孔属于威胁性刺激, 这种刺激能够捕获注意, 因而在快乐面孔中的愤怒面孔更容易找到, 而快乐面孔很容易被忽视。他们把这种现象称为凸显效应(popout effect)。Öhman, Lundqvist和 Esteves (2001)使用与 Hansen不同的面孔图片来研究情绪刺激对注意的影响。Öhman和Lundqvist等的实验采用的是图示面孔, 而不是真实面孔。他认为在Hansen的实验中, 被试之所以搜索愤怒面孔快, 是因为愤怒面孔的异质性更大, 而图示面孔是从真实面孔中抽取出来的, 同质性更高。实验结果表明, 在快乐面孔中搜索愤怒面孔要显著地快于在愤怒面孔中搜索快乐面孔, 这和Hansen的实验结果是一致的。也有一些对焦虑个体的研究, 得到了类似的结果。与搜索微笑面孔相比, 高特质焦虑个体搜索愤怒面孔要更快, 而且识别愤怒面孔也快于微笑面孔(Matsumoto,2010)。此外, 有关威胁性刺激的认知神经机制研究也表明, 威胁性信息导致注意偏向主要发生在视觉信息加工的早期阶段, 这一阶段的加工是一种自动化的加工, 主要依赖于低频轮廓信息(LeDoux, 2000;Vuilleumier, Armony, Driver, & Dolan, 2003; van Le et al., 2013)。
现有研究在不同被试身上以不同的刺激材料重复出了同样的结果, 对这一结果, 研究者们采用环境适应这一进化心理学的观点予以解释(Öhman& Mineka, 2001; Öhman, 2009; Öhman, Soares, Juth,Lindström, & Esteves, 2012; Soares, Lindström,Estevens, & Öhman, 2014)。然而, 要证实威胁性刺激导致注意偏向的进化论解释还有一个非常重要的问题没有解决, 那就是已有实验的生态效度不高。既然人对威胁性刺激的注意偏向加工是适应环境的结果, 那么环境信息与搜索时的光流信息就很重要。现有实验都是通过威胁性刺激图片的呈现来设计的, 没有提供早期人类搜索蛇或蜘蛛时的必要环境信息。如果不提供必要的环境信息, 现有的注意偏向结果也可能解释为新异刺激导致的效应(Peira,Golkar, Öhman, Anders, & Wiens, 2012)。另外, 真实环境中人在搜索威胁性刺激时是一种身体不断接近的过程, 威胁物就算是蛰伏的, 相应的外形颜色特征也会随着接近过程中的光流信息的变化而获得监测。在知觉理论中, 吉布森早就提出并强调光流(optic flow)信息在视知觉加工中的重要作用,他认为利用环境提供的光学分布、结构级差、光流模式、功能承受性和共振等信息可以解释视知觉(Gibson, 1979; Lee & Kalmus, 1980; Sun, Carey, &Goodale, 1992)。在真实的视觉搜索中, 物体远近及角度变化带来的动态信息对理解视觉搜索加工的认知机制很重要, 而威胁性刺激呈现的情境信息(比如远古人类面对的丛林草地环境)对检验已有研究的生态效度同样重要。显然, 无论是环境信息还是光流信息, 已有研究都没有提供。
受过去的实验技术限制, 已有的视觉搜索研究中几乎找不到生态效度好的实验数据。随着计算机信息技术的进步, 新近快速发展起来的虚拟现实技术为解决这一问题提供了可能。虚拟现实技术是利用计算机创设和体验虚拟环境的一种仿真系统, 这一系统的最大特征是沉浸感强, 能够给使用者带来身临其境的感觉。就光流信息而言, 虚拟现实场景中的漫游就是典型的模拟了现实场景中相对运动而导致的物体的表面、边缘等处的光流信息, 这种光流信息本质上体现的是一种似动运动信息。利用虚拟现实技术, 心理学实验不仅可以有仿真的刺激情境, 还可以提供远近角度上的光流动态变化信息,还原进化心理学家假设的远古人类在丛林或草丛中随时搜索发现蛇与蜘蛛的真实场景。
基于以上的分析, 本研究利用虚拟现实技术,模拟早期人类生存的丛林场景, 让被试以浸入式体验的方式执行Öhman和Flykt等(2001)设计的威胁性刺激的视觉搜索任务, 探讨威胁性刺激引起注意偏向的认知机制。将虚拟现实技术应用于视觉搜索实验范式的优势在于, 一是提供刺激的仿真性情境信息, 二是提供刺激的光流信息, 三是提供反应时之外的反应距离信息。反应时之外的反应距离信息在二维呈现的传统实验里几乎无法测试, 但是在虚拟仿真的三维场景中就可以很容易地观测被试做搜索反应时的空间距离。这种反应距离的测量既是反应时或正确率数据的有力补充, 又是进一步了解注意偏向的认知机制的新的有效指标。根据已有二维呈现的实验研究结论, 我们预期在三维虚拟仿真场景中威胁性刺激将同样导致注意偏向反应, 同时反应距离信息能进一步证实这种注意偏向反应。由此, 我们假设, 相比于非威胁性刺激, 被试会在更远的距离下做出对威胁性刺激的搜索确认反应。
此外, 越来越多的研究表明, 动物的搜索要比植物的搜索快(Soares, Esteves, Lundqvist, & Öhman,2009; Soares, 2012)。在 Öhman 和 Flykt等(2001)的经典研究中, 威胁性刺激是蜘蛛与蛇, 而非威胁性刺激是蘑菇与花, 因此在验证威胁性刺激注意偏向时有必要排除威胁性刺激与非威胁性刺激在类型属性上的这种差异。为此, 本研究进一步在虚拟现实情境中比较了威胁性刺激蛇和蜘蛛与非威胁性刺激松鼠和蝉在丛林搜索中的差异, 从而进一步验证威胁性刺激的注意偏向现象, 为威胁性刺激注意加工的进化心理学解释提供最重要的实验证据。
为验证上述观点, 本研究设计了两个实验, 实验1是设计构建野外丛林虚拟仿真环境, 以在较好的生态效度情况下重复Öhman和Flykt等(2001)的经典实验。实验2则是在实验1的基础上将非威胁性刺激替换成松鼠和蝉, 再次验证威胁性刺激的注意偏向加工。
2 方法
实验1将探讨被试在虚拟仿真的丛林搜索实验中, 搜索威胁性刺激是否快于非威胁性刺激, 以演示经典的威胁性刺激注意偏向加工现象。实验2拟将进一步探讨同属动物的威胁性与非威胁性刺激是否仍然存在威胁性刺激的注意偏向加工现象。实验1与实验2的结果一起为威胁性刺激的注意偏向加工的进化心理学解释提供证据支持。
2.1 被试
招募大学生被试共89名(男45名)。实验1共有59人参与。其中10名(男5名)参加了刺激材料的评定实验, 24名(男12名)填写了虚拟现实场景的体验问卷, 20名(男 10名)参加了视觉搜索实验, 5名(男3名)参加了数据剔除标准的评定测验。实验2共有30人参与。其中10名(男5名)参加了刺激材料的评定实验, 20名(男10名)参加了视觉搜索实验。所有被试视力或矫正视力正常, 均为右利手,身体心理健康, 无精神病史, 无任何恐惧症。被试自愿参加实验, 实验完成后获得一定报酬。
2.2 材料
2.2.1 实验用虚拟仿真场景
实验场景包括三部分材料, 第一部分为实验刺激材料, 包括威胁性刺激(蛇与蜘蛛)与非威胁性刺激(蘑菇、花、松鼠、蝉)的仿真模型; 第二部分为丛林草地自然场景, 包括多种草、花、树、石头等;第三部分为实验交互控制材料, 包括进入丛林入口处的一扇门以及第一人称虚拟角色(在实验中这一角色是隐藏的, 被试不可见)。丛林草地场景与门如图1所示。
2.2.2 视觉搜索实验中的刺激物
搜索实验所用刺激物分别为蛇、蜘蛛、花、蘑菇、松鼠、蝉六种。对于刺激物的大小, 由于很难找到精确的尺度来衡量, 所以我们采用了比较相对体积的方法, 这种方法确保了这6种目标物的尺寸的大小接近。对于刺激物的颜色, 这6种模型采用了同样的颜色贴图。6种刺激物的大小及颜色对比如图2所示。
2.2.3 丛林草地中间的小径设置

图1 视觉搜索实验中所使用的仿真丛林场景

图2 搜索实验中所用刺激物模型及颜色对比
为了保证被试以相同的条件进行视觉搜索, 我们在丛林草地中间设置了一条人为踩踏出的略呈“S”型的小道, 被试通过摄像机以直线在小路上行走, 沿小道两边布置目标刺激物(威胁性或非威胁性目标刺激)放置点, 同时在草地上点缀多种野花、杂树、石头等必要自然物。为了方便记录, 路的右边记为位置1到位置7, 路的左边记为位置11到位置17, 如图3左侧所示。需要说明的是, 路径两边的各点分布并不是等距离选取的, 这是防止被试出现搜索猜测。
2.2.4 实验前的评定实验所用材料
评定的材料是从创设的虚拟仿真场景中对4种目标模型的截图, 显示器分辨率为1024 × 768。我们分别从3个不同距离进行截图, 一个是肯定可以看到目标物时, 截图大小为 65 × 65像素; 一个是基本可以看到目标物时, 截图大小为50 × 50像素;一个是隐约可以看到目标物时, 截图大小为 30 ×30像素。其中肯定可以看到目标物时对所有位置进行截图, 共有 56张图片; 基本可以看到目标物时从位置3开始截图, 共有40张图片; 隐约可以看到目标物时从位置5开始截图, 共有24张图片, 3种截图如图3右侧所示。
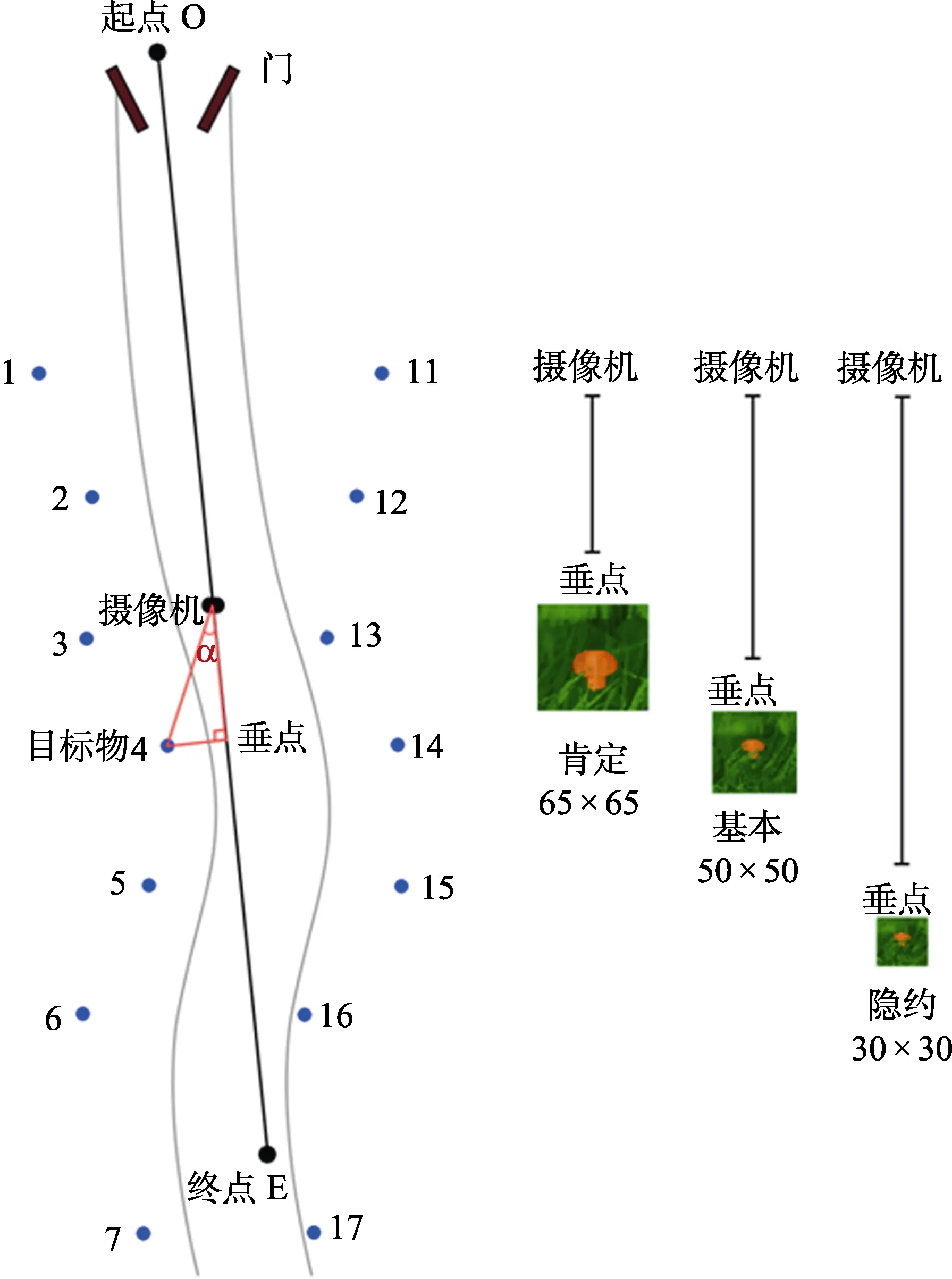
图3 丛林草地上的路径及对所搜索目标物(蘑菇)进行评定时的3种条件
2.2.5 实验中所用材料的制作软件信息
实验中所用的图形场景制作软件主要有3个:3dsmax9, Adobe Photoshop CS 5及 Virtools 5.0。3dsmax9 (Autodesk, San Rafael, California, U.S.)主要用于制作和修改模型, 为了使后期计算机运行起来更流畅, 在制作和修改模型时, 在保证逼真的前提下, 尽量减少模型的面数。Adobe Photoshop CS5(Adobe Systems Inc, SAN jose, California, U.S.)主要用于对模型贴图的修改。Virtools 5.0虚拟现实游戏引擎(Dassault Systèmes, Vélizy, Villacoublay Cedex,France)用于创设野外丛林场景。我们将3dsmax中做成的模型导入进来, 在Virtools 5.0中整合成完整场景, 然后进行交互设计和编程, 实现被试能够在场景中进行固定路径的视觉搜索。
2.3 设计
实验分为实验 1与实验 2, 每个实验包括两个部分, 第一部分为虚拟现实实验场景的真实感体验评定和视觉搜索目标物物理属性差异的评定, 第二部分是正式虚拟现实场景中的视觉搜索实验。
虚拟现实场景的体验评定采用国外较为常用的IGROUP问卷(Schubert, Friedmann, & Regenbrecht,2001; Felnhofer et al., 2015)。视觉搜索目标物的物理属性差异评定是通过一个简单的选择反应时实验来完成的。物理属性差异评定采用 4(刺激物模型:蛇、蜘蛛、花、蘑菇) × 14(位置:虚拟场景中的 14个位置)被试内设计, 刺激物采用电脑图片呈现方式, 记录被试的反应时和错误率。
正式的视觉搜索实验为2(刺激类型:威胁性刺激、非威胁性刺激) × 14(位置:虚拟场景中的 14个位置)被试内设计, 记录反应时、错误率以及被试做反应时的反应距离。反应时为从门开到被试按键反应的时间, 错误率为反应错误试次所占该条件总试次数的百分比。对于反应距离的定义, 我们将反应距离与传统实验的反应时(Response Time)相对应。具体做法为, 首先, 我们将被试在三维场景中做反应时摄像机的位置(即反应位置)距离目标位置的连线, 然后, 将反应位置距离目标位置对摄像机行走路径做垂点的连线, 最后, 两条连线做夹角,夹角的正切值即为反应距离(Response Distance),如图4中的tanα。这样一来, 反应距离包含反应位置与目标位置的距离信息及测量视角, 故而是一种更为全面的指标。图4示意了反应距离的计算方式。在 Virtools软件中, 固定路径漫游中的某时刻点的坐标无法获取, 程序是封装模块, 除非进行非常底层的二次开发。实际上, 固定路径漫游是匀速运动,我们根据被试的反应时进行事后计算, 相应的位置坐标同样可以得到。因此, 本研究中就以某个目标位置(比如位置 4)搜索时的平均反应时 RT为基础(即摄像机行走S(图4中的2D子图)所用的时间)进行计算。程序中最初设定的路径起点O到终点E的总时间为T, 将RT与T相比就可以得到A点的坐标。图中B点为该目标位置向摄像机路径做垂线的垂点, 目标位置、摄像机位置、垂点构成三角形, 计算现tanα值, 以此值作为RD的指标。
特别要说明, 实验中采用了我们实验室开发的计时器来完成的, 它是基于CPU时钟来记录的, 比较精确。如果采用Virtools软件本身提供的记时器,由于它是基于刷新率来记录时间的, 所以在加载高清场景时, 刷新率不容易稳定, 记时精度也不够高,值得注意。
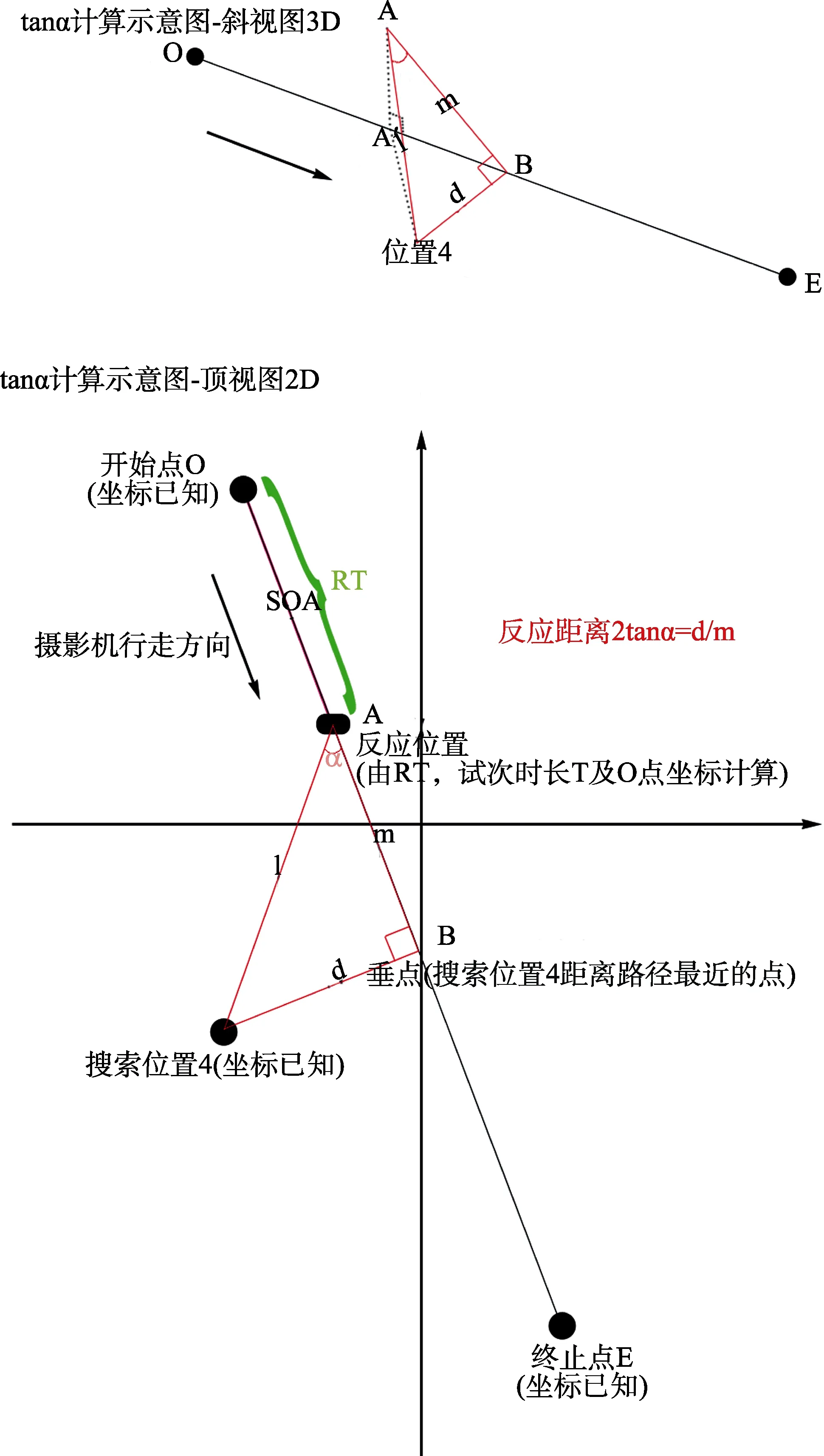
图4 反应距离计算示意图
我们在设计时也考虑了光流信息。光流信息包含了被观察物体的运动信息, 也包含有关景物三维结构的丰富信息, 我们在设计时通过虚拟现实技术展现了搜索物体相对运动的信息。摄像机在场景中移动, 搜索的物体距离摄像机越来越近, 其相对摄像机向摄像机反方向移动。也包含了搜索场景的信息。前人因技术限制没有考虑光流信息, 我们通过该技术提供了视觉搜索的光流信息, 更好地为威胁场景中刺激搜索提供了更完善的证据支持。
因实验1的威胁性刺激模型为动物(蛇与蜘蛛),非威胁性刺激模型为植物(花与蘑菇), 为排除此动植物两种自变量混淆对实验结果的影响, 我们进行了实验2。实验2将刺激物模型全部更换为动物,分别是:蛇、蜘蛛、松鼠、蝉。其余与实验1保持一致。
2.3.1 虚拟现实场景体验评定
尽管当前的Oculus DK2完全浸入式的虚拟现实呈现设备的临场感已经非常明显, 但为了准确评价所建场景是否有效提高了实验任务的生态效度,我们对实验1的场景进行了评定。被试进行8次搜索, 每个模型搜索两次, 然后填写IGROUP问卷。该问卷包括14个问题, 每个问题都采用0~6评分,标记从无真实感到非常真实的感觉。目前我们研究组已经对该问卷进行了中国化的修订, 相关研究报告正在投稿中。
24名被试填写的IGROUP问卷结果如表1所示。从评价的3个指标的均值上来看, 所建野外丛林场景的存在感、沉浸感与真实感都在中间值3以上。体验感的评定与国外两款虚拟游戏场景(Tomb Raider, Half Life)的评价结果相近(Schubert, Friedmann,& Regenbrecht, 2001; Felnhofer et al., 2015), 实验场景生态效度较好。
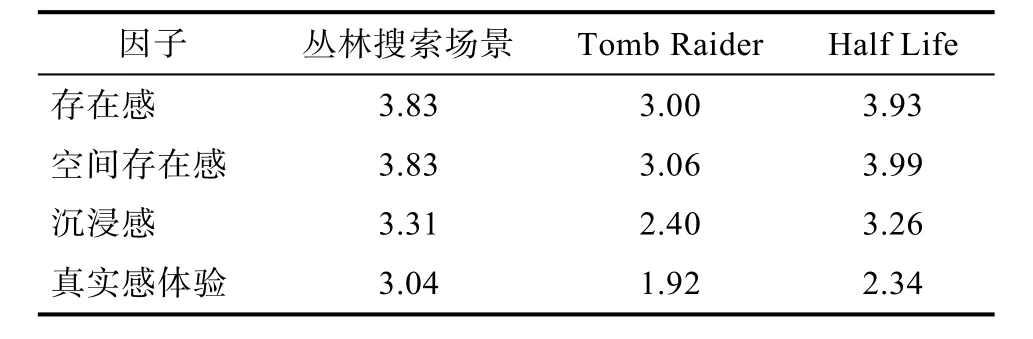
表1 丛林搜索场景与国外虚拟现实游戏场景IGROUP评定结果比较
2.3.2 目标物评定实验
采用传统的选择反应时实验范式进行刺激物的评定, 实验程序使用 E-prime 2.0 (Psychology Software Tools, Sharpsburg, Pennsylvania, U.S.)软件编程实现。实验分为练习和正式实验两阶段, 正式的评定实验分为3个组块(block)。第一个组块的图片全部是肯定可以看到目标物时的图片, 每个位置的图片呈现 2次, 所以第一个组块共 112个试次(trial)。第二个组块的图片是基本可以看到目标物时的图片, 每个位置的图片呈现 2次, 所以第二个组块共有 80个试次。第三个组块的图片是隐约可以看到目标物时的图片, 每个位置的图片呈现2次,所以第三个组块共有48个试次。总共3个组块有240个试次。实验流程是先呈现 500 ms的注视点,接着呈现图片, 被试按键反应后呈现 800 ms的空屏, 实验进入到下一个试次。实验过程中, 被试双手食指分别放在“F”键和“J”键上, 实验要求如果被试看到的是蛇和蜘蛛, 按“F”键进行反应; 如果看到的是花和蘑菇或者松鼠和蝉(分别对应实验 1与实验2), 按“J”键进行反应。实验流程如图5所示。
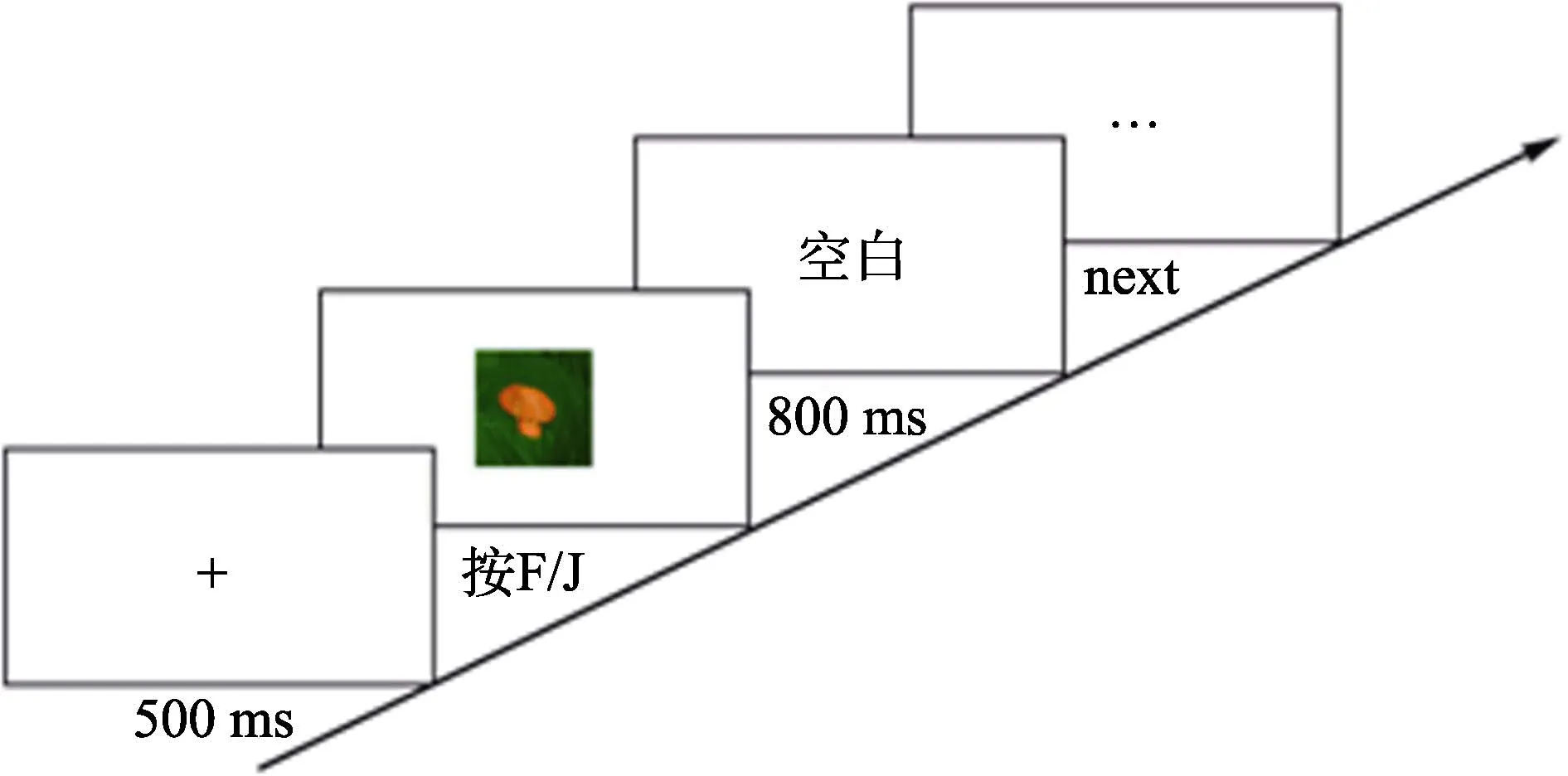
图5 刺激评定实验流程图
2.3.3 物理显著性分析
采用Itti和Koch (2000)文章中的方法进行物理显著性分析。我们使用该文章中所用的 MATLAB程序对材料图片进行评定处理, 得到 Saliency和Winner Take All两幅图, 如图6所示。SaliencyMap是一幅清晰的二维地图, 包含对视觉环境中物体的显著性信息的编码。其具体方法是, 第一步, 将输入的图片中的每一个特征进行计算, 并转换为图标地形特征地图; 第二步, 将这些特征地图合并成为一幅地图, 这幅地图包含任意一个位置相对其他位置的显著性; 第三步, 找出显著地图中的最大值,在指定的时间点, 最大值所在的位置就是下一个注意视觉区。Winner-take-all (WTA)是一种算法, 这种算法由 Tsotsos等人(1995)提出, 用于 SaliencyMap分析方法的第三步, 即找出显著地图中的最大值。得到每种条件下这两幅图后, 我们对 Winner Take All图中心红框内的白色方形区域进行明度提取。我们首先使用Photoshop截取每种条件下的白色方形区域, 然后使用 MATLAB程序对白色方形区域按HSV颜色分类方式提取明度值(V), 最后提取与SaliencyMap中心白色区域相等大小区域, 即为边长64个像素正方形的区域内4096个点的明度值。我们对4种条件下的明度值进行单因素方差分析。
2.3.4 视觉搜索实验
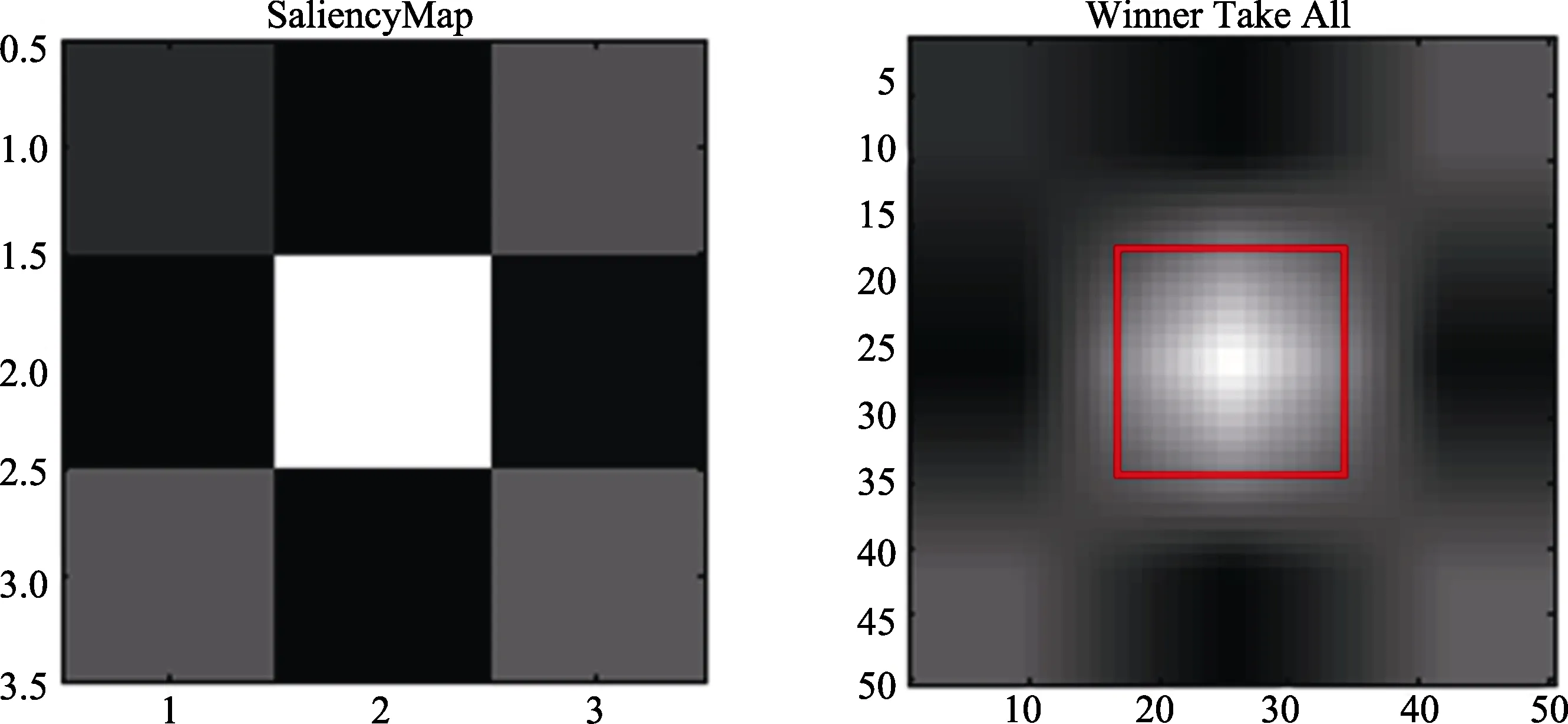
图6 物理显著性分析示意图
正式的视觉搜索实验采用与Öhman和Flykt等人(2001)一样的实验范式, 但实验材料与情境均采用前述提到的三维仿真模型。实验程序在虚拟现实软件平台Virtools 5.0上运行。正式实验分为8个组块, 每个组块中目标物在14个位置出现2次共28个试次, 由于第一个试次的计时器计时可能不准确,因此在每个组块前加了一个缓冲试次, 所以每个组块实际上有 29个试次。我们对实验顺序进行了拉丁方平衡。根据Öhman和Flykt等的实验范式, 蛇和蜘蛛属于威胁性刺激, 花、蘑菇、松鼠与蝉属于非威胁性刺激。在每一个试次中, 目标物为威胁性刺激或非威胁性刺激。
实验时, 被试坐在电脑旁, 戴上头盔显示器。实验开始后, 摄像机镜头会自动地沿着预先设定好了的路径移动。实验要求被试仔细观察整个场景,如果被试找到了所要搜索的目标, 则按游戏手柄的“LB”键, 如果一直到镜头停止移动都没有找到所要搜索的目标, 则按游戏手柄的“RB”键。按键反应后, 再按游戏手柄的“A”键即可以进入到下一个试次。在每个场景中, 整个森林的场景是不变的, 改变的只是分心物和目标物。
2.3.5 数据剔除标准测试
整个实验是在一个虚拟的三维仿真场景中进行, 目前还没有关于虚拟现实场景中反应时实验的报告, 为了能够更好地查看虚拟仿真场景中反应时测量, 我们特别增加设计了一个空场景中搜索刺激物的测试, 并以此作为后续反应时实验中反应记录的一个参考标准。并且设置这一测试也同时考虑到被试会对多个位置点的目标物进行搜索反应, 因此获得一个不同位置点简单反应时值也是一个不错的参考指标。就虚拟现实场景渲染距离来看, 本研究中的实验场景均没有出现远处物体不能渲染的问题。为此, 我们招募 5名被试进行测试, 求得可以看到刺激物的最小反应时。考虑到场景开门时,位置1~4和位置11~14可以看清楚的, 而位置 5、位置6、位置7、位置15、位置16、位置17在开门后并不能真正看清楚。但是, 在虚拟现实场景漫游的动态情形下完成搜索实验, 由于有很多类似颜色的花草干扰, 被试不认真时很容易误认而做出按键反应。这正说明虚拟现实情境实验在提高生态效度的同时也增加了被试错误辨认的机率。搜索的模型为蛇、蜘蛛、花、蘑菇, 结果如表2所示。正式实验时, 当被试的反应时小于测试的反应时最小值,则予以剔除。
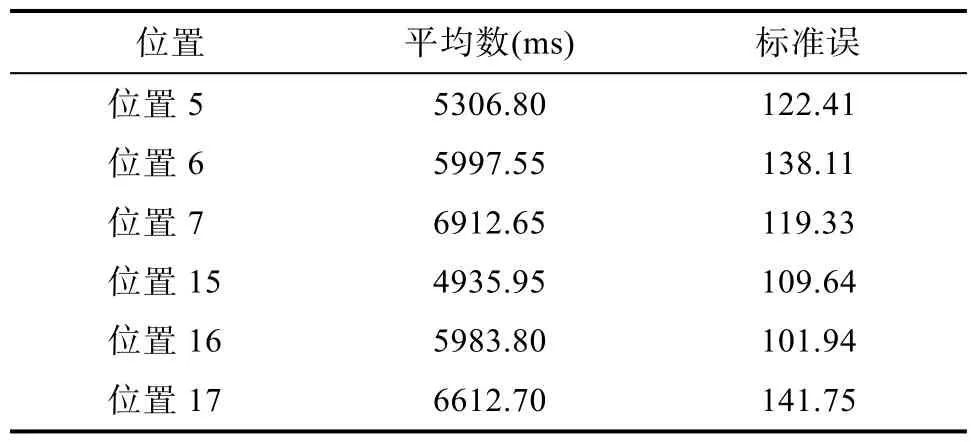
表2 不同位置反应时最小值(平均数、标准误)
3 结果
3.1 实验1结果
3.1.1 刺激材料评定实验
为了防止极端值以及错误反应对实验结果的影响, 剔除平均数在3个标准差之外的数据以及错误反应的数据, 共剔除极端值和错误反应数据 72个, 剔除比例为3%。使用SPSS 22.0进行统计分析。
对3种不同距离的错误率分别进行单因素重复测量方差分析。肯定可以看到目标物时模型种类的主效应不显著,
F
(3, 27) = 1.91,p
> 0.05。基本可以看到目标物时模型种类的主效应不显著,F
(3, 27) =0.61,p
> 0.05。隐约可以看到目标物时模型种类的主效应不显著,F
(3, 27) = 1.48,p
> 0.05。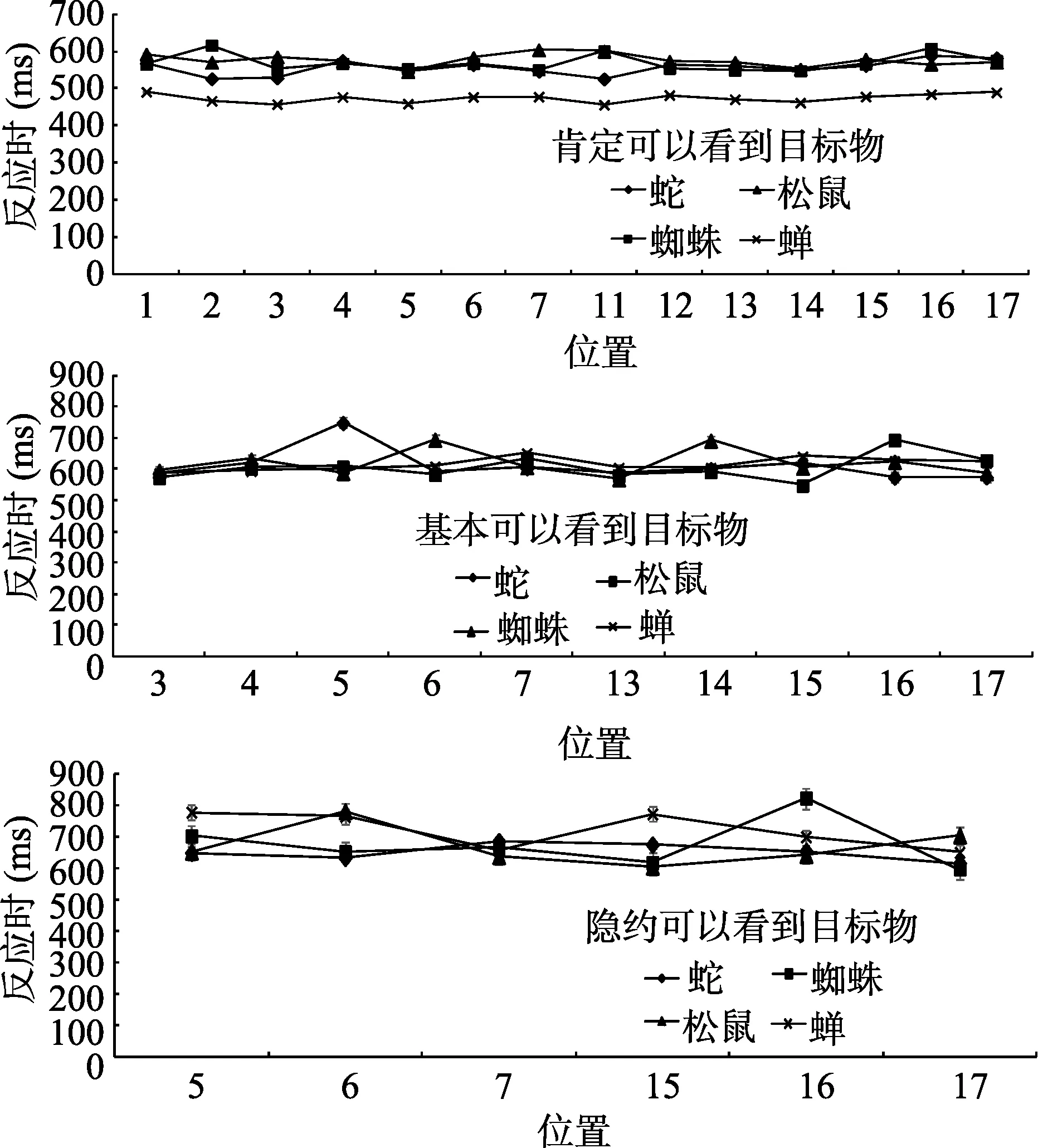
图7 实验1刺激材料评定实验反应时结果
对3种不同距离的反应时进行重复测量方差分析。肯定可以看到目标物时, 模型种类的主效应不显著,
F
(3, 27) = 0.47,p
>0.05。位置的主效应不显著,
F
(13, 117) = 0.43,p
> 0.05。模型种类与位置的交互作用也不显著,F
(39, 351) = 0.97,p >
0.05。基本可以看到目标物时, 模型种类的主效应不显著,
F
(3, 27) = 0.16,p
> 0.05。位置的主效应显著,F
(9, 81) = 2.02,p
< 0.05, η= 0.18。进行事后检验发现, 对位置进行两两比较表明位置3和位置5差异显著,p
< 0.05。模型种类与位置的交互作用为边缘显著,F
(27, 243) = 2.45,p
< 0.05, η= 0.21。简单效应分析表明, 位置 15时, 蛇和蜘蛛差异显著,p
<0.05。位置16时, 蛇和蜘蛛差异显著,p
< 0.01。其他位置时, 两两刺激之间差异均不显著。虽然位置15和 16时, 蛇和蜘蛛的差异显著, 但蛇与蜘蛛都属于威胁性刺激, 而且不存在蛇中找蜘蛛和蜘蛛中找蛇的组合, 因此它们之间的知觉辨别力差异对实验结果没有影响。隐约可以看到目标物时, 模型种类的主效应不显著,
F
(3, 27) = 0.88,p
> 0.05。位置的主效应不显著,F
(5, 45) = 1.36,p
> 0.05。模型种类与位置的交互作用也不显著,F
(15, 135) = 1.66,p
> 0.05。在3种不同距离时, 无论是反应时还是正确率,模型种类的主效应均不显著, 表明4种模型在知觉辨别力上无显著差异, 排除了后面搜索过程中由于模型本身引起的变量混淆。
3.1.2 视觉搜索实验
所有被试的正确率较高。被试各个条件下错误率、反应时、反应距离如图8所示, 其中反应距离的平均正切值威胁性刺激(
M
= 0.61,SD
= 0.08)比非威胁性刺激(M
= 0.73,SD
= 0.10)小, 以具体场景中的反应距离计算, 威胁性刺激(11.79米,SD
=0.77)也同样比非威胁性刺激(10.60米,SD
= 0.73)为远。场景中呈现的物体大小(0.75米 × 0.75米), 因为场景较大, 所以物体适当放大了以使看起来是正常的比例。在威胁刺激条件下, 物体的平均视角为3.65°, 而非威胁性刺激条件下, 物体的平均视角为4.05°。为了防止极端值以及错误反应对实验结果的影响, 剔除平均数在3个标准差之外的数据、错误反应的数据以及位置5、位置6、位置7、位置15、位置16、位置17小于各自最小值的数据。共剔除81个数据, 剔除比例为1.8%。使用SPSS 22.0进行统计分析。我们还进一步地把反应时实验数据与前述剔除测试的反应时标准进行了对比, 结果发现, 前述各位置点上的简单反应时都落在反应时实验中的反应时数据的3个标准差之外。这说明, 虚拟仿真场景中的搜索辨别反应时的记录与传统实验是一致的。
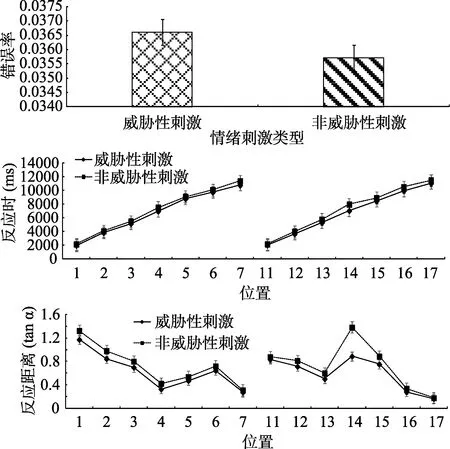
图8 实验 1视觉搜索实验被试错误率、反应时、反应距离(平均数±标准误)结果
对错误率进行配对样本
t
检验, 结果表明刺激类型差异不显著,t
(19)= 0.25,
p
> 0.05。对反应时进行2(刺激类型:威胁性刺激、非威胁性刺激)×14(位置:虚拟场景中的14个位置)重复测量方差分析。刺激类型主效应显著,
F
(1, 19) =301.72,p
< 0.01, η= 0.94。位置的主效应显著,F
(13,247) = 449.89,p
< 0.01, η= 0.96。刺激类型和位置的交互作用显著,F
(13, 247) = 2.60,p
< 0.05, η=0.12。简单效应分析表明, 威胁性刺激与非威胁性刺激的差异均显著(p
s < 0.05), 搜索威胁性刺激显著快于非威胁性刺激。对反应距离进行2(刺激类型:威胁性刺激、非威胁性刺激)×14(位置:虚拟场景中的 14个位置)重复测量方差分析。刺激类型主效应显著,
F
(1, 19) =301.72,p
< 0.001, η= 0.94。位置的主效应显著,F
(13, 247) = 202.84,p
< 0.001, η= 0.997。刺激类型和位置的交互作用显著,F
(13, 247) = 3.87,p
< 0.05,η= 0.88。简单效应分析表明, 威胁性刺激与非威胁性刺激的差异均显著(p
s < 0.05), 搜索威胁性刺激的反应距离比搜索非威胁性刺激的反应距离小。我们对反应距离和反应时进行了皮尔逊积差相关分析, 发现负相关显著,
r
= −0.67,p
< 0.01。相关分析表明, 反应时越长, 反应距离越短。上述结果表明, 在提高了实验生态效度的情况下, 威胁性刺激的注意偏向加工依然存在, 支持了经典的威胁性刺激优先加工假设(Cisler, Bacon, & Williams,2009; Cisler & Koster, 2010; Yiend, 2010; 张禹, 罗禹, 赵守盈, 陈维, 李红, 2014)。如前言中所讨论到的, 近期有不少研究揭示, 动物的搜索要比植物的搜索快(Soares et al., 2009; Soares, 2012)。而Öhman和 Flykt等(2001)的经典研究所使用的非威胁性刺激是植物(蘑菇与花)威胁性刺激是动物(蛇与蜘蛛)。因此在验证威胁性刺激注意偏向时有必要排除威胁性刺激与非威胁性刺激在类型属性上的差异。为此, 本研究在实验1的基础上进一步比较了蛇和蜘蛛同动物类非威胁性刺激(松鼠和蝉)的比较。3.2 实验2结果
3.2.1 刺激材料评定实验
为了防止极端值以及错误反应对实验结果的影响, 剔除平均数在3个标准差之外的数据以及错误反应的数据。共剔除76个数据, 剔除比例为3%。使用SPSS 22.0进行统计分析。
对3种不同距离时的错误率分别进行单因素重复测量方差。肯定可以看到目标物时模型种类的主效应不显著,
F
(3, 27) = 1.80,p
> 0.05。基本可以看到目标物时模型种类的主效应不显著,F
(3, 27) =3.72,p
> 0.05。隐约可以看到目标物时模型种类的主效应不显著,F
(3, 27) = 1.12,p
> 0.05。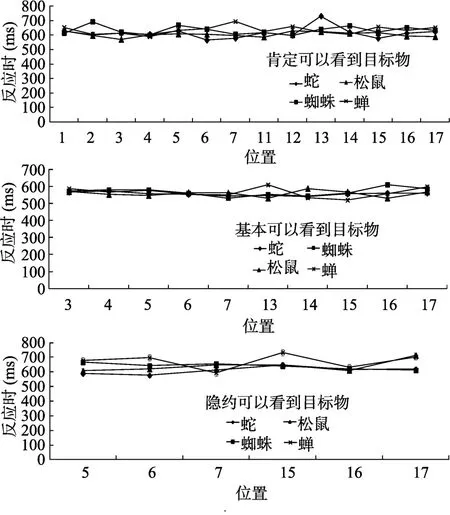
图9 实验2刺激材料评定实验反应时结果
对3种不同距离的反应时进行重复测量方差分析。肯定可以看到目标物时, 模型种类的主效应不显著,
F
(3, 27) = 1.82,p
> 0.05。位置的主效应不显著,F
(13, 117) = 0.76,p
> 0.05。模型种类与位置的交互作用也不显著,F
(39, 351) = 1.03,p
> 0.05。基本可以看到目标物时, 模型种类的主效应不显著,F
(3, 27) = 0.56,p
> 0.05。位置的主效应不显著,F
(9,81) = 0.64,p
> 0.05。模型种类与位置的交互作用也不显著,F
(39, 351) = 1.41,p
> 0.05。隐约可以看到目标物时, 模型种类的主效应不显著,F
(3, 27) =0.89,p
> 0.05。位置的主效应不显著,F
(5, 45) = 0.97,p
> 0.05。模型种类与位置的交互作用也不显著,F
(15, 135) = 0.76,p
> 0.05。在3种不同距离时, 无论是反应时还是正确率,模型种类的主效应均不显著, 表明4种模型在知觉辨别力上无显著, 排除了后面搜索过程中由于模型本身引起的变量混淆。
3.2.2 视觉搜索实验
各个条件下错误率、反应时、反应距离如图10所示, 其中反应距离的平均正切值威胁性刺激(
M
=0.57,SD
= 0.08)比非威胁性刺激(M
= 0.71,SD
=0.10)小, 以具体反应距离计算, 威胁性刺激(12.47米,SD
= 0.59)也同样比非威胁性刺激(10.69米,SD
=0.59)为远。因为场景较大, 物体的大小也同实验 1有所放大(0.75米 × 0.75米)。在威胁性刺激条件下,物体的平均视角为 3.46°, 在非威胁性刺激条件下物体的平均视角为 4.02°。为了防止极端值以及错误反应对实验结果的影响, 剔除平均数在3个标准差之外的数据、错误反应的数据以及位置5、位置6、位置7、位置15、位置16、位置17小于各自最小值的数据, 共剔除68个数据, 剔除比例为1.5%。使用SPSS 22.0进行统计分析。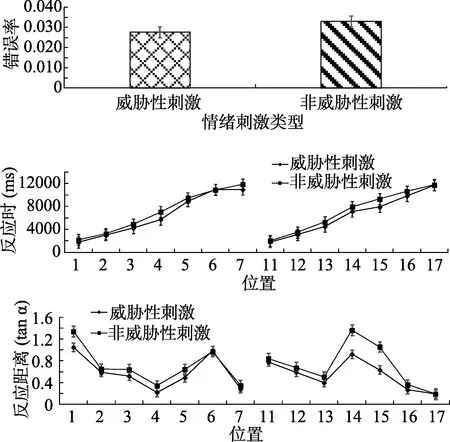
图10 实验2视觉搜索实验被试错误率、反应时、反应距离(平均数±标准误)结果
对错误率进行配对样本 t检验, 结果表明刺激类型差异不显著,
t
(19)= −0.92,p
> 0.05。对反应时进行2(刺激类型:威胁性刺激、非威胁性刺激) × 14(位置:虚拟场景中的14个位置)重复测量方差分析。刺激类型主效应显著,
F
(1, 19) =59.99,p
< 0.01, η= 0.76。位置的主效应显著,F
(13,247) = 1462.51,p
< 0.01, η= 1.00。刺激类型和位置的交互作用显著,F
(13, 247) = 9.04,p
< 0.01, η=0.94。简单效应分析表明, 除位置6与位置17之外,其余位置威胁性刺激与非威胁性刺激的差异均显著(p
s < 0.05), 搜索威胁性刺激显著快于搜索非威胁性刺激。位置6与位置17差异不显著的原因是:在摄像机的行驶过程中, 被试最先会看到前 6个位置, 行驶一段距离后, 会看到中间4个位置, 最后看到位于最后方的 4个位置, 这三部分每部分内部出现的时间差异较小, 因此会有差异不显著的可能性。对反应距离进行2(刺激类型:威胁性刺激、非威胁性刺激) × 14(位置:虚拟场景中的14个位置)重复测量方差分析。刺激类型主效应显著,
F
(1, 19) =59.99,p
< 0.001, η= 0.76。位置的主效应显著,F
(13,247) = 1462.51,p
< 0.001, η= 1.00。刺激类型和位置的交互作用显著,F
(13, 247) = 9.04,p
< 0.01, η=0.94。简单效应分析表明, 除位置6与位置17差异不显著之外, 其余位置威胁性刺激与非威胁性刺激的差异均显著(p
s < 0.05), 搜索威胁性刺激的反应距离比搜索非威胁性刺激的反应距离小。对反应距离和反应时进行了皮尔逊积差相关分析, 负相关边缘显著,
r
= −0.33,p
= 0.08。同实验1的结果, 反应时越长, 反应距离越短。上述结果表明, 在把威胁性刺激替换成同样的动物后, 蛇与蜘蛛的搜索反应仍然显著高于非威胁刺激, 说明威胁性刺激的注意偏向加工现象是独立于刺激类型和动植物属性影响的。考虑到模型显著性的可能影响,我们采用前人的方法对模型在搜索中的显著性进行了分析(Itti & Koch, 2000)。根据上述搜索实验的结果判定, 实际搜索到的目标物均在“基本可见”条件下发生, 因此我们对此条件下的模型在场景中的截图在 MATLAB中进行了显著性分析, 结果如图11所示。从显著性分析结果图上可以看出, 无论是注意点还是显著性分布图均较一致, 未见明显差异。为了更好地比较搜索不同目标物在显著性上的差异, 我们对显著性结果做了进一步的量化分析。选取显著性分析明度分布图的中心区域(64像素×64像素), 计算各像素点的明度值(V), 然后对不同目标物的明度值进行单因素方差分析, 结果表明,实验1中明度主效应不显著,F
(3, 16380) = 0.48,p
>0.05。实验2中的明度主效应不显著,F
(3, 16380) =1.51,p
> 0.05, 各目标物的平均明度值见表3。这些结果表明, 在实际搜索场景中, 各目标物本身的显著性彼此之间并无差异。4 讨论
本研究的目的是要在提高实验生态效度的前提下, 进一步验证威胁性刺激注意偏向加工现象,从而检验威胁性刺激注意加工偏向现象的进化心理学解释的可靠性。研究采用完全浸入式的虚拟仿真技术, 首次抽象模拟了人类祖先生存的野外丛林环境, 为视觉搜索威胁性刺激提供了必要的野外丛林环境与接近刺激时的光流动态信息, 最终实现了提高传统行为实验生态效度的目的。研究中收集的环境体验数据表明, 实验中建构的仿真环境能够给人带来较好的真实体验, 能够保证实验生态效度提高的有效性。实验1与实验2的结果表明, 在提高了生态效度的情况下, 人们在视觉搜索威胁性刺激时确实要比搜索非威胁性刺激快, 即使把非威胁性刺激由蘑菇和花替换成松鼠和蝉后, 被试仍然是搜索蛇与蜘蛛要比搜索松鼠和蝉快, 验证了威胁性刺激的注意加工偏向现象, 夯实了进化心理学解释的实验基础。除此之外, 本研究还首次发现了注意偏向加工在空间距离尺度上的存在证据, 即在视觉搜索中, 人对威胁性刺激搜索反应的距离显著远于对非威胁性刺激搜索反应的距离。反应距离的数据与反应时的数据负相关显著, 两个指标可互相补充,趋势一致。无论是反应时数据还是反应距离数据都稳定一致地支持威胁性刺激的注意偏向加工效应,支持进化心理学的解释。
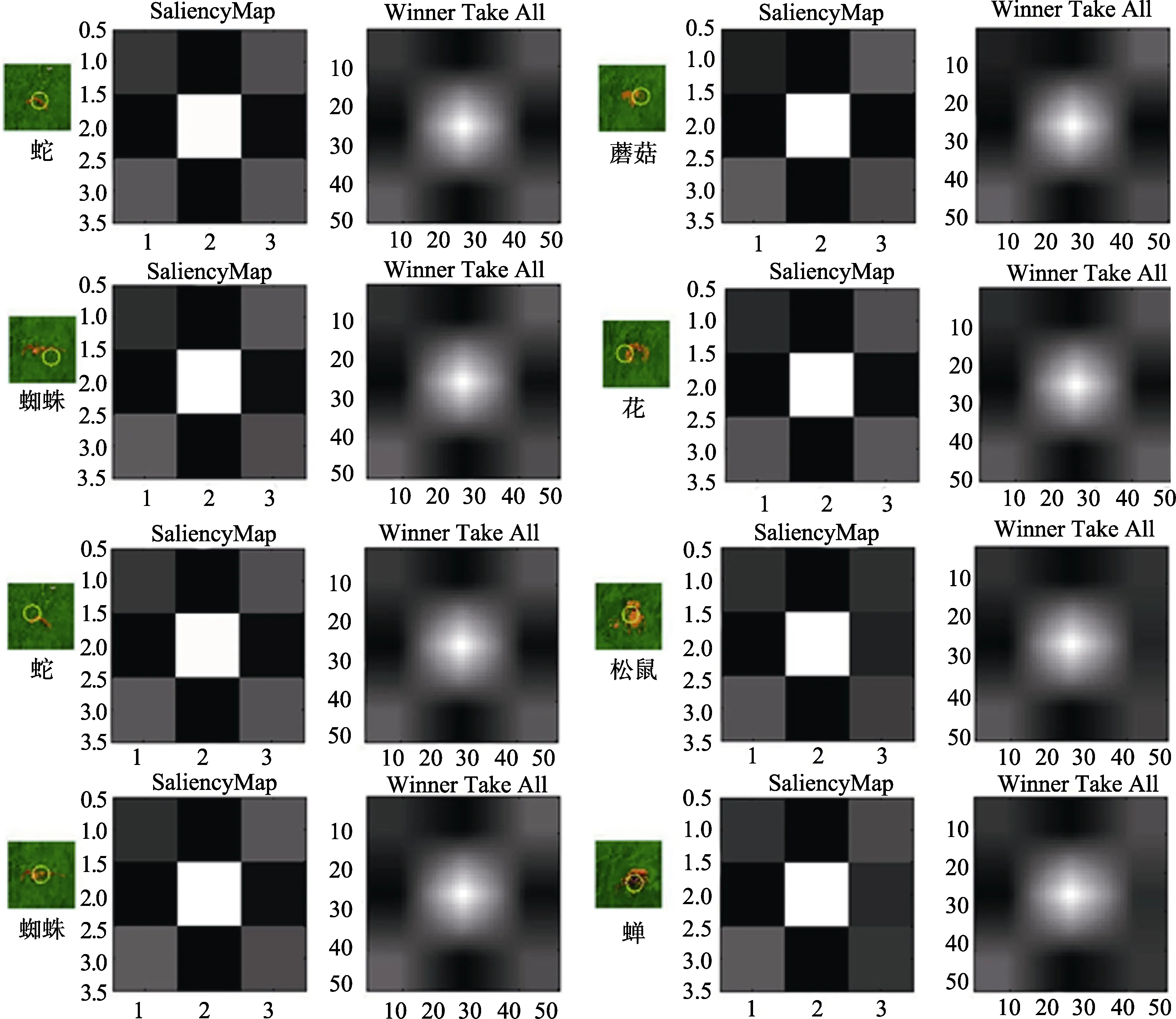
图11 实验中所用模型在搜索局部场景中的显著性

表3 图片物理显著性的明度值平均数(括号内为标准误)
实验中的反应距离信息有重要的验证性价值。传统的二维图片呈现实验只是从反应时指标上揭示这种威胁性刺激的优先反应, 尚不能从空间距离上模拟动态的搜索过程, 以更好的生态效度来呈现这一结果。本研究的空间距离信息首次弥补了这方面的信息。与传统二维图片实验结果一样, 威胁性刺激优先加工与已有认知神经科学的研究预期相一致。Vuilleumier等(2003)的研究揭示人对威胁性刺激的加工更多地是存在于视觉信息加工的早期阶段, 这一阶段的加工是一种自动化的加工, 主要依赖于低频轮廓信息。本研究通过反应距离的计算证实, 威胁性刺激(3.5°视角处)相比于非威胁性刺激(4.0°视角)在更远的距离上就被检测确认。当然因为是动态的搜索, 被试在看到搜索物到按下键还有一段时间, 这个时间会导致实际记录的视角变得更大。从刺激信息上看, 距离越远越只能依赖于低频轮廓信息进行加工。Vuilleumier等(2003)的研究是通过人为地操纵刺激物的空间频率信息来验证威胁性刺激的早期自动加工的, 而本研究则是通过自然的仿真情境观察直接得到这一结果。虽然我们从反应时数据上完全可以推测出反应距离的结果, 但是这种推测仍然是建立在可以测量搜索过程中的空间距离这一条件之下的, 这与传统实验中固定刺激物与被试眼睛之间的距离有着很大的不同, 这种不同正是本研究提高实验生态效度的结果。因此,反应距离信息在视觉搜索实验中可以作为反应时数据之外的一个重要补偿性指标。同时, 它具有进化心理学的重要意义。反应距离数据提示我们, 搜索威胁性刺激更可能是一种低频自动化加工。从进化论的角度来看, 一种心理行为如果是适应环境的结果, 那么它更可能是情绪性的反应, 是快速的自动化反应(Tooby & Cosmides, 1990)。反应距离数据恰好告诉我们, 威胁性刺激的搜索反应是快速的自动化反应, 符合适应进化的特征。
实验中的情境信息很重要。在已有研究中, 有少数研究并没有重复出威胁性刺激导致注意偏向的结果, 比如快乐面孔的搜索要比愤怒面孔快。Juth, Lundqvist, Karlsson和Öhman (2005)采用了视觉搜索的范式让被试在中性面孔中寻找快乐、愤怒和恐惧目标面孔。结果表明, 探测快乐面孔要显著地快于探测愤怒和恐惧面孔。Savage, Lipp, Craig,Becker和 Horstmann (2013)使用来自 Ekman和Friesen面孔图片库的面孔图片作为实验材料, 结果也表明, 在中性面孔中搜索快乐面孔要快于搜索愤怒面孔。现在看来, 这些相反的结果很可能是由实验的生态效度不够导致的。就一般的社会刺激情境而言, 威胁性刺激并不一定会启动人类的生存适应器, 而如果提供了一些必要的情境信息这些结果就可能反转。那么什么样的信息才是关键的情境信息?本研究是通过概念释义的方式来确定的, 即根据 Öhman (2009)的人类祖先的野外生存适应假说,我们将这种必要的情境具体化为野外丛林, 草中的蛇与蜘蛛, 那么对于威胁性的面孔来说, 可能就要用更为社会化的情境来解释。
如何理解对威胁性刺激的注意偏向加工的认知机制还需要进一步的研究证据的支持。注意偏向是优先捕获, 还是特异性特征的加工, 目前尚无法确认。知觉过程包含两种加工:自上而下的加工和自下而上的加工。自上而下的加工是由有关知觉对象的一般知识开始的加工, 而自下而上的加工是由外部刺激开始的加工。Lindsay和Norman (1977)将自上而下的加工称为概念驱动加工, 而将自下而上的加工称为数据驱动加工。在此基础上, Torralba,Oliva, Castelhano和Henderson (2006)提出背景引导模型, 认为观察者根据从场景中获得的信息, 结合个体的知识经验, 从而注视场景中最可能存在目标物的位置。同时, 该模型认为对目标物的搜索依赖于自上而下的知识经验, 并结合自下而上的场景信息。Theeuwes (2010)认为人类对威胁自己生存的信息始终保持着注意, 这可以看作始终存在的自上而下的“预设”任务目标。从本研究的结果来看, 对威胁性刺激的早期预警反应可能是开始于某种特异性特征的驱动, 继而启动自上而下加工的确认所导致的。
本研究使用了虚拟现实技术, 大大提高了实验结果的生态效度, 但也存在着一些问题。首先, 虚拟现实环境中的远景渲染与实际环境中不太一样,考虑到虚拟现实场景一般都比较大, 相应的硬件性能还不够强, 一般对远处场景及物件的渲染都有一个限定距离, 这可能会给实验带来一些影响。好在已有研究表明, 威胁性刺激的情绪反应主要是在意识层面(Peira et al., 2012)。本研究为了避免这个问题的影响, 专门进行了最小反应时的测定, 并据此剔除那些低于这一反应时数值的反应。其次, 虚拟仿真场景目前受限于硬件性能, 场景的逼真度还不够, 尤其是头盔的刷新率及分辨率与真实场景比较还有很大的距离, 这可能也会影响被试的浸入感。本研究为了弱化这一影响, 对场景中的外围模型进行了面数优化, 保留了搜索范围内的物件模型的高精度, 并且在性能较高的工作站上运行实验程序,以求实验中的最佳体验。头盔问题是目前虚拟现实实验技术中的瓶颈, 在短时间内还难以解决。最后,虚拟现实技术在提高实验的生态效度的同时也为实验的精确控制提出了挑战。自然场景条件下我们的搜索是完全不受限制的, 但是在虚拟现实技术中,这种限制总是无法避免, 比如头盔的视野范围, 触压觉、动觉等感官信息的提供, 仿真模型彼此之间在大小与形状上的差异导致遮挡或物件的知觉水平可能有差异等等。尽管如此, 虚拟现实在改善心理学已有实验的生态效度上确实是前所未有的, 在心理学领域的未来应用前景值得期待。目前, 虚拟现实技术在心理学上的应用还比较少, 就我们所做的数据库检索发现, 目前, 虚拟现实技术在心理学上的应用还比较少, 有文献支持临床干预的有效性(Urech, Krieger, Chesham, Mast, & Berger, 2015),但在基础心理研究上除了传统的空间认知研究外几乎没有多少应用, 关于情绪刺激对注意偏向影响的研究根本没有。本研究不仅为已有研究提供了有力的支持, 还为新技术、新方法应用于心理学研究提供了范例。在未来的研究中, 我们期待将虚拟现实技术特有的沉浸性、互动性和构想性特点与心理学实验设计紧密结合, 进一步提高心理学实验的生态效度。
5 结论
本研究的结论是, 在自然场景的虚拟仿真场景中, 人们搜索威胁性刺激要快于非威胁性刺激。生态效度与理论假说相结合为注意偏向加工说提供了进一步的事实依据, 同时本研究也是运用虚拟现实技术提高实验生态效度的一次尝试。
致谢:
李然、姚维佳对本文进行了认真的校对, 在此表示感谢。Buss, D. M. (1999).
Evolutionary psychology: The new science of the mind
. Boston: Allyn and Bacon.Cisler, J. M., Bacon, A. K., & Williams, N. L. (2009).Phenomenological characteristics of attentional biases towards threat: A critical review.
Cognitive Therapy &Research, 33
(2), 221–234.Cisler, J. M., & Koster, E. H. W. (2010). Mechanisms of attentional biases towards threat in anxiety disorders:An integrative review.
Clinical Psychology Review,30
(2), 203–216.Dudeney, J., Sharpe, L., & Hunt, C. (2015). Attentional bias towards threatening stimuli in children with anxiety:A meta-analysis.
Clinical Psychology Review, 40
,66–75.Felnhofer, A., Kothgassner, O. D., Schmidt, M., Heinzle, A.K., Beutl, L., Hlavacs, H., & Kryspin-Exner, I. (2015). Is virtual reality emotionally arousing? Investigating five emotion inducing virtual park scenarios.
International Journal of Human-Computer Studies, 82
, 48–56.Gibson, J. J. (1979).
The ecological approach to visual perception.
Boston, MA: Houghton Mifflin.Hansen, C. H., & Hansen, R. D. (1988). Finding the face in the crowd: An anger superiority effect.
Journal of Personality & Social Psychology, 54
(6), 917–924.Itti, L., & Koch, C. (2000). A saliency-based search mechanism for overt and covert shifts of visual attention.
Vision Research, 40
(10–12), 1489–1506.Janczyk, M., Augst, S., & Kunde, W. (2014). The locus of the emotional Stroop effect: A study with the PRP paradigm.
Acta Psychologica, 151
, 8–15.Juth, P., Lundqvist, D., Karlsson, A., & Öhman, A. (2005).Looking for foes and friends: Perceptual and emotional factors when finding a face in the crowd.
Emotion, 5
(4),379–395.Ledoux, J. E. (2000). Emotion circuits in the brain.
Annual Review of Neuroscience, 23
, 155–184.Lee, D. N., & Kalmus, H. (1980). The optic flow field: The foundation of vision [and discussion].
Philosophical Transactions of the Royal Society B:
Biological Sciences,290
(1038), 169–179.Lindsay, P. H., & Norman, D. A. (1977).
Human information processing: An introduction to psychology
.New York: Academic Press.LoBue, V. (2010). And along came a spider: An attentional bias for the detection of spiders in young children and adults.
Journal of Experimental Child Psychology, 107
(1),59–66.LoBue, V., & Rakison, D. H. (2013). What we fear most: A developmental advantage for threat-relevant stimuli.
Developmental Review, 33
(4), 285–303.Matsumoto, E. (2010). Bias in attending to emotional facial expressions: Anxiety and visual search efficiency.
Applied Cognitive Psychology, 24
(3), 414–424.Mogg, K., & Bradley, B. P. (2006). Time course of attentional bias for fear-relevant pictures in spider-fearful individuals.
Behaviour Research & Therapy, 44
(9), 1241–1250.Mogg, K., Bradley, B. P., & Hallowell, N. (1994).Attentional bias to threat: Roles of trait anxiety, stressful events, and awareness.
The Quarterly Journal of Experimental Psychology Section A: Human Experimental Psychology, 47
(4), 841–864.Öhman, A. (2009). Of snakes and faces: An evolutionary perspective on the psychology of fear.
Scandinavian Journal of Psychology, 50
(6), 543–552.Öhman, A.,Flykt, A., & Esteves, F. (2001). Emotion drives attention: Detecting the snake in the grass.Journal of Experimental Psychology: General, 130
(3), 466–478.Öhman, A., Lundqvist, D., & Esteves, F. (2001). The face in the crowd revisited: A threat advantage with schematic stimuli.
Journal of Personality & Social Psychology,80
(3), 381–396.Öhman, A., & Mineka, S. (2001). Fears, phobias, and preparedness: Toward an evolved module of fear and fear learning.
Psychological Review, 108
, 483–522.Öhman, A., Soares, S. C., Juth, P., Lindström, B., &Esteves, F. (2012). Evolutionary derived modulations of attention to two common fear stimuli: Serpents and hostile humans.
Journal of Cognitive Psychology, 24
(1),17–32.Peira, N., Golkar, A., Öhman, A., Anders, S., & Wiens, S.(2012). Emotional responses in spider fear are closely related to picture awareness.
Cognition and Emotion,26
(2), 252–260.Peng, D. L. (2012).
General psychology
(4th ed.). Beijing,China: Beijing Normal University Press.[彭聃龄. (2012).
普通心理学
(第4版). 北京: 北京师范大学出版社.]Savage, R. A., Lipp, O. V., Craig, B. M., Becker, S. I., &Horstmann, G. (2013). In search of the emotional face:Anger versus happiness superiority in visual search.
Emotion, 13
(4), 758–768.Schubert, T., Friedmann, F., & Regenbrecht, H. (2001).The experience of presence: Factor analytic insights.
Presence, 10
(3), 266–281.Soares, S. C. (2012). The lurking snake in the grass:Interference of snake stimuli in visually taxing conditions.
Evolutionary Psychology, 10
(2), 187–197.Soares, S. C., Esteves, F., Lundqvist, D., & Öhman, A. (2009).Some animal specific fears are more specific than others:Evidence from attention and emotion measures.
Behaviour Research & Therapy, 47
(12), 1032–1042.Soares, S. C., Lindström, B., Esteves, F., & Öhman, A.(2014). The hidden snake in the grass: Superior detection of snakes in challenging attentional conditions.
PLoS One
,9
(12), e114724.Stormark, K. M., & Hugdahl, K. (1996). Peripheral cuing of covert spatial attention before and after emotional conditioning of the cue.
International Journal of Neuroscience, 86
(3–4), 225–240.Stormark, K. M., Nordby, H., & Hugdahl, K. (1995).Attentional shifts to emotionally charged cues:Behavioural and ERP data.
Cognition & Emotion, 9
(5),507–523.Sun, H. J., Carey, D. P., & Goodale, M. A. (1992). A mammalian model of optic-flow utilization in the control of locomotion.
Experimental Brain Research,91
(1), 171–175.Theeuwes, J. (2010). Top-down and bottom-up control of visual selection.
Acta Psychologica, 135
(2), 77–99.Thrasher, C., & LoBue, V. (2016). Do infants find snakes aversive? Infants' physiological responses to"fear-relevant" stimuli.
Journal of Experimental Child Psychology, 142
, 382–389.Tooby, J., & Cosmides, L. (1990). The past explains the present: Emotional adaptations and the structure of ancestral environments.
Ethology & Sociobiology,11
(4–5), 375–424.Torralba, A., Oliva, A., Castelhano, M. S., & Henderson, J.M. (2006). Contextual guidance of eye movements and attention in real-world scenes: The role of global features in object search.
Psychological Review, 113
(4),766–786.Tsotsos, J. K., Culhane, S. M., Kei, W. W. Y., Lai, Y. Z.,Davis, N., & Nuflo, F. (1995). Modeling visual attention via selective tuning.
Artificial Intelligence, 78
(1-2),507–545.Urech, A., Krieger, T., Chesham, A., Mast, F. W., &Berger, T. (2015). Virtual reality-based attention bias modification training for social anxiety: A feasibility and proof of concept study.
Frontiers in Psychiatry, 6
,154.van Le, Q., Isbell, L. A., Matsumoto, J., Nguyen, M., Hori,E., Maior, R. S., … Nishijo, H. (2013). Pulvinar neurons reveal neurobiological evidence of past selection for rapid detection of snakes.
Proceedings of the National Academy of Sciences of the United States of America,110
(47), 19000–19005.Vuilleumier, P., Armony, J. L., Driver, J., & Dolan, R. J.(2003). Distinct spatial frequency sensitivities for processing faces and emotional expressions.
Nature Neuroscience, 6
(6), 624–631.Wang, F. X., Li, W. J., Yan, Z. Q., Duan, Z. H., & Li, H.(2015). Children's attention detection to snakes:Evidence from eye movements.
Acta Psychologica Sinica, 47
(6), 774–786.[王福兴, 李文静, 颜志强, 段朝辉, 李卉. (2015). 幼儿对威胁性刺激蛇的注意觉察: 来自眼动证据.
心理学报, 47
(6), 774–786.]Yiend, J. (2010). The effects of emotion on attention: A review of attentional processing of emotional information.
Cognition & Emotion, 24
(1), 3–47.Zhang, Y., Luo, Y., Zhao, S. Y., Chen, W., & Li, H. (2014).Attentional bias towards threat: Facilitated attentional orienting or impaired attentional disengagement.
Advances in Psychological Science, 22
(7), 1129–1138.[张禹, 罗禹, 赵守盈, 陈维, 李红. (2014). 对威胁刺激的注意偏向: 注意定向加速还是注意解除困难?.
心理科学进展
,22
(7), 1129–1138.]猜你喜欢
辽河(2022年4期)2022-06-09 01:56:03
心理学报(2022年1期)2022-01-21 02:50:24
当代陕西(2020年23期)2021-01-07 09:25:24
智慧少年·故事叮当(2020年2期)2020-03-08 14:19:16
人与自然(2019年4期)2019-07-26 12:52:37
当代陕西(2019年12期)2019-07-12 09:12:02
小朋友·快乐手工(2018年3期)2018-04-22 11:48:52
小学阅读指南·低年级版(2017年6期)2017-06-12 08:22:47
小朋友·快乐手工(2015年1期)2015-03-13 00:05:24
当代经济科学(2015年2期)2015-02-03 03:00:52
