
限饲对妊娠中期母羊血液生化指标及内脏脂肪组织脂肪代谢的影响
2018-06-14 01:00周小玲颜琼娴谭支良
动物营养学报 2018年6期
杨 宏 周小玲,3 颜琼娴 谭支良
(1.中国科学院亚热带农业生态研究所,亚热带农业生态过程重点实验室,畜禽养殖污染控制与资源化技术国家 工程实验室,湖南省畜禽健康养殖工程技术研究中心,农业部中南动物营养与饲料科学观测实验站;长沙 410125; 2.中国科学院大学,北京 100049;3.塔里木大学动物科学学院,阿拉尔 843300;4.湖南省植物功能成分 利用协同创新中心,长沙 410128;5.湖南省畜禽安全生产协同创新中心,长沙 410128)
由于天然饲草资源数量和品质、生产管理技术及环境应激等因素限制,妊娠母羊易出现营养供给不足。妊娠期母羊营养受限不仅影响母体妊娠附属物的发育、后期繁殖性能、胎儿组织器官发育及子代生长性能,而且对母体物质、能量代谢如脂肪代谢也有重要影响[1-3]。前人对母体妊娠期营养受限的研究主要集中在胚胎植入前后及妊娠晚期,在生产中对母体的营养干预也主要在妊娠晚期及泌乳阶段,对妊娠中期母体营养需要及能量代谢的研究较少。妊娠中期是指妊娠1/3~2/3阶段,这一时期是母体妊娠附属物发育和胎儿器官形态建成及功能完善的重要时期[4],期间营养受限时母体的代谢变化及可能的代偿机制对妊娠中期营养需要研究及制订相应饲养管理策略具有重要意义。
妊娠期母体营养受限,母体会动员自身营养储备,尤其是脂肪组织供能来维持胚胎发育和自身能量需求,以缓冲外界不良环境对胚胎发育的影响。脂肪组织是机体重要的能量贮存器官,也是重要的脂肪代谢调控器官。根据分布部位,脂肪组织可以分为皮下脂肪组织和内脏脂肪组织(visceral adipose tissue,VAT)。VAT主要在瘤胃网膜、肠系膜和肾周等分布,其相对于皮下脂肪组织,细胞脂肪动员能力更强、速度更快,其代谢产物能直接通过门静脉进入血液循环,进而影响机体脂质代谢和能量代谢。内脏脂肪代谢与机体胰岛素(INS)抵抗、脂肪代谢紊乱、糖尿病、脂肪肝、代谢综合征等[5-8]的产生密切相关。VAT动员和代谢变化,是反映机体营养物质充足或缺乏的重要指征,也是机体对营养不足环境的重要应答路径。
本试验通过研究限饲对妊娠中期母羊脂肪代谢相关血液生化指标、VAT脂肪酸组成及脂肪代谢相关基因表达的影响,试图初步解析妊娠中期低营养水平状态下妊娠母羊VAT脂肪动员及其对机体脂肪代谢的影响机制。
1 材料与方法
1.1 试验动物及饲养管理
试验采用单因素随机区组试验设计。健康湘东黑山羊16只,妊娠(45±3) d,体重(29.86±3.07) kg,随机分配至对照组(C组,100%妊娠营养需求)和限饲组(R组,60%妊娠营养需求),每组8只。试验期为妊娠45~100 d,试羊共接受55 d试验处理,单栏饲养,饲粮精粗比50∶50,每日等量饲喂2次(08:00和16:00),先精后粗,2组基础饲粮相同,R组饲喂量为C组的60%,自由饮水。限制期间母羊干物质采食量为:C组(1.14±0.04) kg/d,R组(0.60±0.02) kg/d,R组实际限饲量为C组的52%。限饲结束时母羊体增重为:C组(5.91±0.70) kg,R组(2.65±0.22) kg。基础饲粮组成及营养水平见表1。
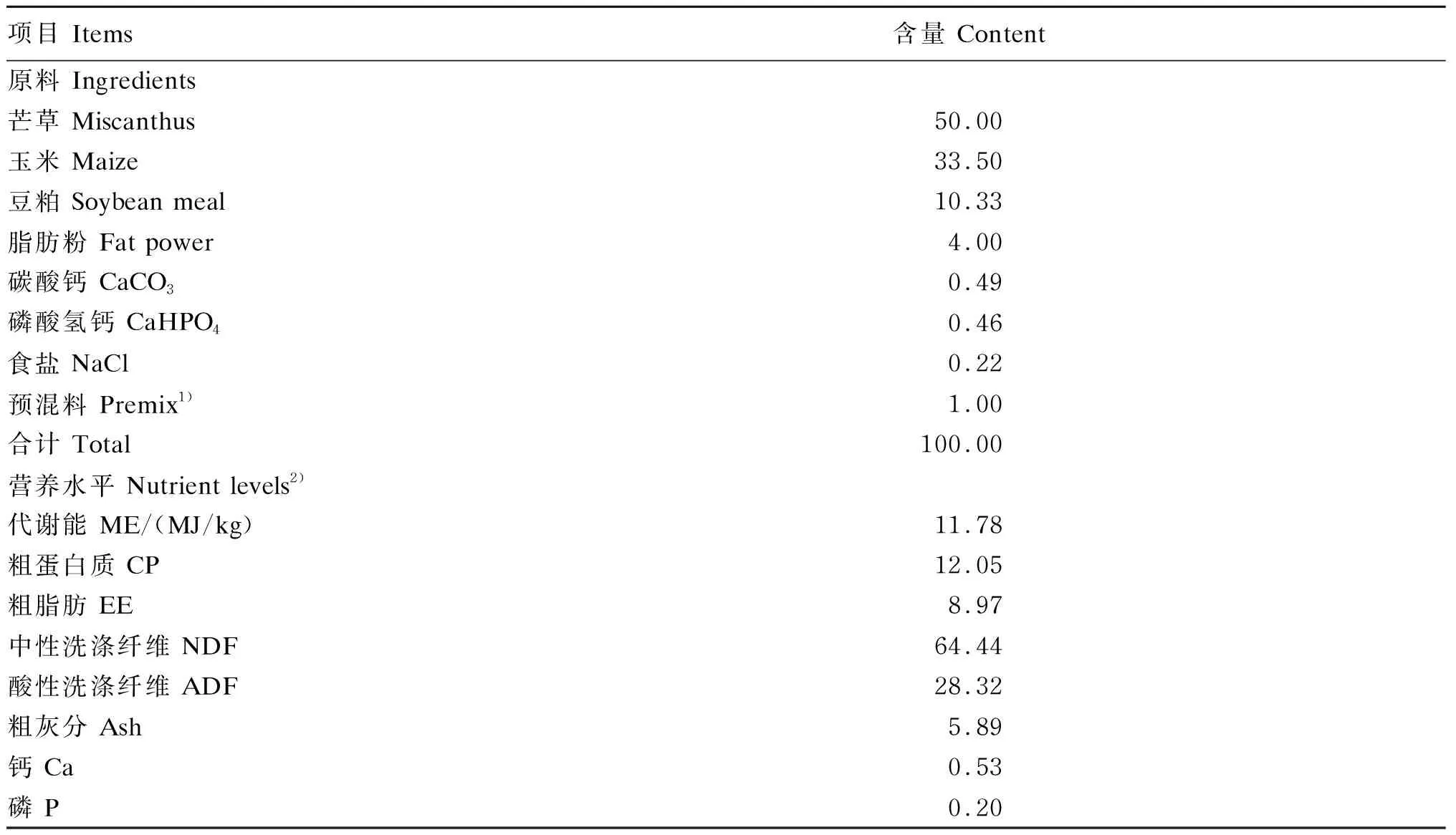
表1 基础饲粮组成及营养水平(干物质基础)
1)每千克预混料含有One kg of premix contained the following:MgSO4·H2O 119 g,FeSO4·7H2O 2.5 g,CuSO4·5H2O 0.8 g,MnSO4·H2O 3 g,ZnSO4·H2O 5 g,Na2SeO310 mg, KI 40 mg,CoCl2·6H2O 30 mg,VA 95 000 IU,VD 17 500 IU,VE 18 000 IU。
2)代谢能为计算值,计算方法参考张宏福等[9],其余指标为实测值,测定方法参考张丽英等[10]。ME was a calculated value calculated using the method of Zhang et al[9], and the others were measured values measured using the methods of Zhang et al[10].
1.2 样品采集及分析
1.2.1 血样采集与分析
试验结束后,试羊空腹24 h,于次日(妊娠101 d)09:00经颈静脉采血,肝素钠抗凝,室温静置4 h,在4 ℃条件下以1 200×g离心10 min分离血浆,-20 ℃保存待测。血液葡萄糖(GLU)、甘油三酯(TG)、总胆固醇(CHOL)、高密度脂蛋白胆固醇(HDL-C)和低密度脂蛋白胆固醇(LDL-C)含量采用日立7600全自动生化分析仪分析(试剂购自北京利德曼生化股份有限公司)。血液INS、胰高血糖素、瘦素(LEP)和脂联素(ADP)含量采用酶联免疫法测定(试剂盒购自武汉华美生物工程有限公司)。血液游离脂肪酸(FFA)含量采用试剂盒测定(试剂盒购自南京建成生物工程研究所)。
1.2.2 脂肪组织样本采集及分析
经颈动脉放血、剥皮开膛后,分离瘤胃网膜脂肪组织(简称网膜脂)、肠系膜脂肪组织(简称系膜脂)和肾周脂肪组织(简称肾周脂),经液氮速冻后于-80 ℃冷冻保存。
1.2.2.1 脂肪酸含量测定
脂肪组织经冷冻干燥和研碎后,称取样品约0.5 g,置于50 mL离心管中,加入4 mL苯-石油醚混合溶剂(按体积比1∶1混合),密闭浸提24 h后,加入4 mL氢氧化钾-甲醇溶液(0.4 mol/L)快速甲酯化,振荡混匀3 min后,静置30 min。加入超纯水分层,9 200×g离心10 min取上层溶液,加入适量无水硫酸钠除去残留水分。取上述200 μL上清液,加800 μL正己烷稀释,经0.22 μm滤膜过滤后备测。采用Agilent 7890A气相色谱仪(Agilent公司,美国)检测上清液中脂肪酸含量,详细步骤参见Ichihara等[11]的方法。采用峰面积归一化法来定量,各脂肪酸的含量以单个脂肪酸在总甲酯化脂肪酸中所占的质量百分比表示。
1.2.2.2 实时定量PCR(RT-PCR)
总RNA提取:用RNAios Plus试剂(TaKaRa公司)提取总RNA,方法见说明书,用超微量紫外分光光度计于260 nm和1%变性琼脂糖凝胶电泳测定总RNA的浓度与纯度,以260和280 nm吸光度值比值(OD260 nm/OD280 nm)在1.8~2.1的为总RNA纯度较好,以电泳后28S rRNA和18S rRNA的灰度值比2∶1为依据,评判提取总RNA的质量。
反转录:取2 μg总RNA采用Prime ScriptRTreagent kit反转录试剂盒(TaKaRa公司)进行反转录反应,反应采用40 μL体系,方法见说明书,反转录产物cDNA于-80 ℃保存备用。
引物设计:目的基因和内参基因β-肌动蛋白(β-actin)的引物使用Primer premier 5.0软件根据GenBank中发表的山羊基因序列设计,用Primer-BLAST工具在线进行引物特异性分析。引物由上海生工生物工程技术服务有限公司合成,引物序列及参数见表2。

表2 RT-PCR特异性引物序列及参数

续表2项目Items引物序列 Primer sequences (5'-3')产物大小Produce size/bp登录号Accession No.肝激酶B1LKB1上游:GGACACCTTCTCTGGCTTCA下游:CCCTTCCCGATGTTCTCAA126XM_018050463.1解偶联蛋白1UCP1上游:TGGGACACACAGCATCAGAC下游:TAAAGGCACTGGAGGTCAGG156XM_018061376.1解偶联蛋白2UCP2上游:GAGGTTGCAGGAATCGTCAT下游:GGGAAAGGTGATGAGGTCAG120XM_018058858.1过氧化物酶体增殖物激活受体γ共激活因子1α PGC-1α上游:CCGAGAATTCATGGAGCAAT下游:GATTGTGTGTGGGCCTTCTT184XM_018049155.1腺苷酸活化蛋白激酶的催化亚基α2 AMPKα2上游:TTGATGATGAGGTGGTGGAG下游:CCGTGAGAGAGCCAGAGAGT138XM_018044652.1腺苷酸活化蛋白激酶的非催化亚基β1 AMPKβ1上游:CCACCACATCTCCTCCAAGT下游:GAGCACCATCACTCCATCCT135XM_013970630.2
使用荧光定量PCR仪(ABI 7900 HT,ABI公司,美国)进行RT-PCR。采用10 μL反应体系:SYBR Premix Ex Taq Ⅱ(2×)5.0 μL,上游引物(10 μmol/L)0.4 μL,下游引物(10 μmol/L)0.4 μL,cDNA模板1.0 μL,dH2O(RNase Free)3.2 μL。反应条件:95 ℃ 30 s预变性;95 ℃ 5 s变性,60 ℃ 20 s延伸,40个循环。熔解曲线分析:95 ℃ 5 s,65 ℃60 s。降温:50 ℃ 30 s。每个样本3个重复。本试验中以β-actin作为内参基因,以对照组基因循环阈值(Ct)的平均值作为对照组Ct,目的基因相对表达量计算采用2-△△Ct法。
1.3 数据统计与分析
所有试验数据经Excel 2013软件整理后采用SPSS 19.0软件进行t检验统计分析,以P<0.05为差异显著,以0.05≤P≤0.10为差异有显著趋势,P>0.10为差异不显著,结果以平均值(mean)及标准误(SE)表示。
2 结 果
2.1 血液生化指标
由表3可知,与C组相比,R组母羊血液中胰高血糖素和FFA含量显著升高(P<0.05),血液LEP、ADP和HDL-C含量显著下降(P<0.05),血液INS、GLU、TG、CHOL、LDL-C含量及HDL-C/CHOL无显著变化(P>0.10)。
2.2 内脏脂肪组织脂肪酸组成
由表4可知,与C组相比,R组网膜花生四烯酸(C20∶4n6)、棕榈油酸(C16∶1)、亚油酸(C18∶2n6c)、二十碳三烯酸(C20∶3n6)和多不饱和脂肪酸(PUFA)含量显著降低(P<0.05),网膜豆蔻酸(C14∶0)和棕榈酸(C16∶0)含量分别有降低和升高的趋势(0.05≤P≤0.10),检测的网膜其余脂肪酸含量2组间无显著差异(P>0.10)。
由表5可知,与C组相比,R组系膜豆蔻酸含量显著降低(P<0.05),系膜亚油酸、α-亚麻酸(C18∶3n6)、二十碳烯酸(C20∶1)、二十碳三烯酸和PUFA含量有降低的趋势(0.05≤P≤0.10),系膜反式油酸(C18∶1n9t)含量有升高趋势(0.05≤P≤0.10),检测的系膜其余脂肪酸含量2组间无显著差异(P>0.10)。
由表6可知,与C组相比,R组肾周脂亚油酸、花生四烯酸、二十碳三烯酸和PUFA含量显著降低(P<0.05),肾周脂豆蔻酸含量有降低趋势(0.05≤P≤0.10),检测的其余脂肪酸含量2组间无显著差异(P>0.10)。
2.3 内脏脂肪组织脂肪代谢相关基因表达
由表7可知,与C组相比,R组网膜脂腺苷酸活化蛋白激酶的催化亚基α2(AMPKα2)基因表达有上调的趋势(0.05≤P≤0.10),其余网膜脂基因表达2组间无显著差异(P>0.10)。

表3 限饲对妊娠中期母羊血液生化指标的影响
0.05≤P≤0.10为有差异趋势,P<0.05为差异显著。下表同。
0.05≤P≤0.10 indicated tendency of difference, andP<0.05 indicated significant difference. The same as below.

表4 限饲对妊娠中期母羊网膜脂脂肪酸组成的影响
饱和脂肪酸=豆蔻酸+棕榈酸+十七烷酸+硬脂酸+花生酸;单不饱和脂肪酸=棕榈油酸+反式油酸+油酸+二十碳烯酸;多不饱和脂肪酸=亚油酸+γ-亚麻酸+α-亚麻酸+二十碳三烯酸+花生四烯酸+二十二碳六烯酸。下表同。
SFA=C14∶0+C16∶0+C17∶0+C18∶0+C20∶0; MUFA=C16∶1+C18∶1n9t+C18∶1n9c+C20∶1; PUFA=C18∶2n6c+C18∶3n6+C20∶3n6+C20∶4n6+C22∶6n3. The same as below.

表5 限饲对妊娠中期母羊系膜脂脂肪酸组成的影响
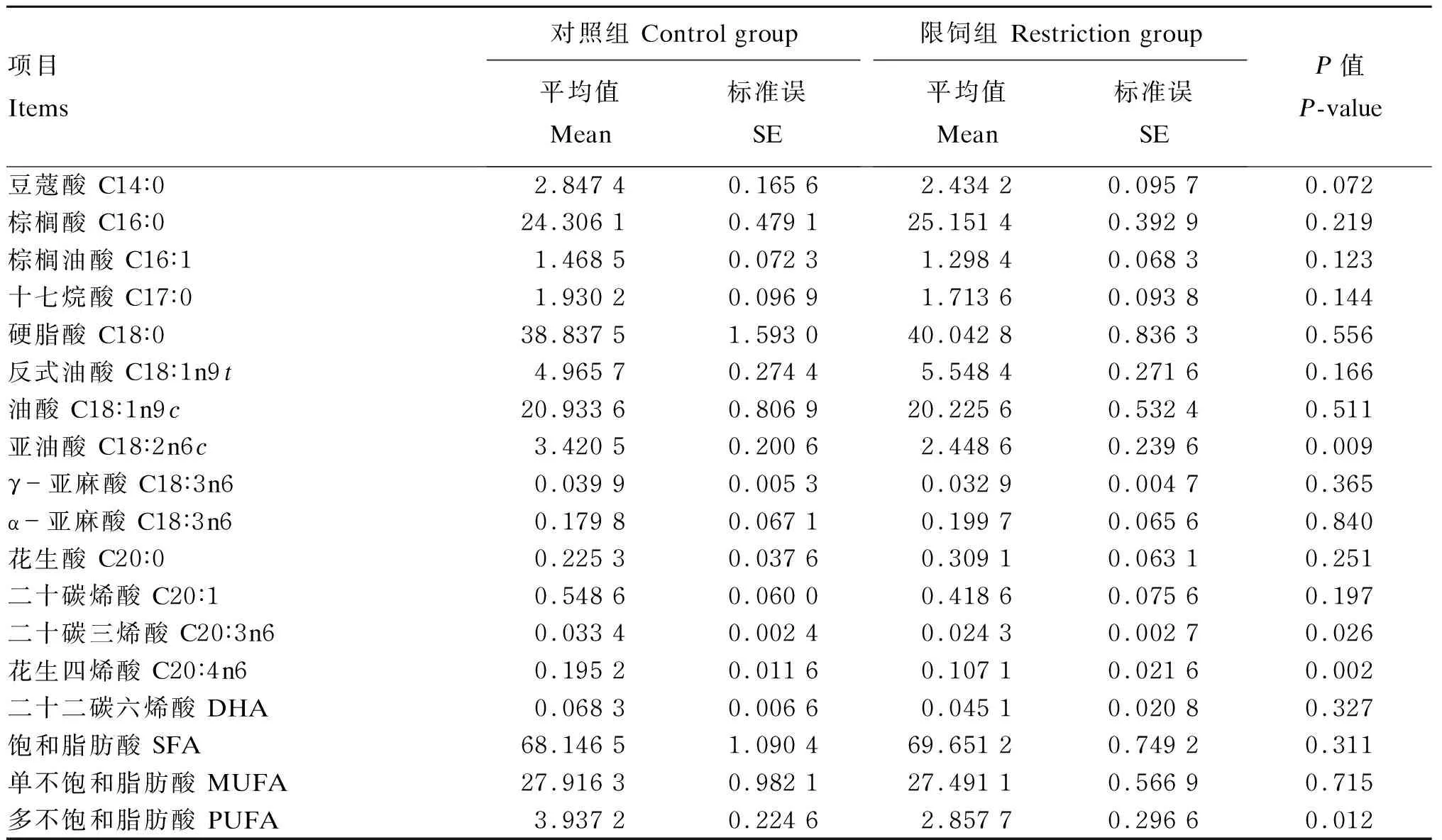
表6 限饲对妊娠中期母羊肾周脂脂肪酸组成的影响
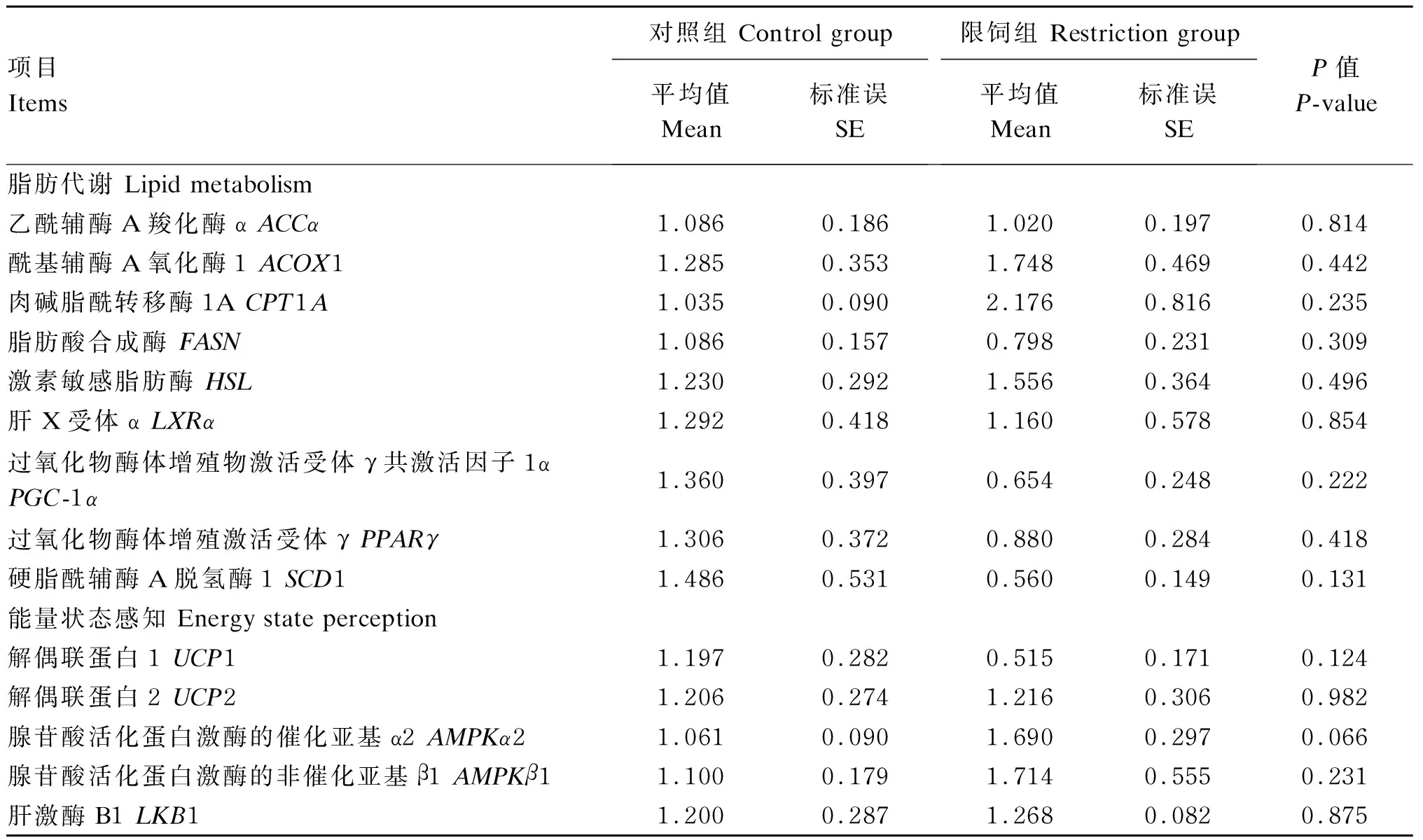
表7 限饲对妊娠中期母羊网膜脂脂肪代谢相关基因表达的影响
由表8可知,与C组相比,R组系膜脂脂肪酸合成酶(FASN)、硬脂酰辅酶A脱氢酶1(SCD1)和解偶联蛋白2(UCP2)基因表达有下调的趋势(0.05≤P≤0.10),系膜脂过氧化物酶体增殖激活受体γ(PPARγ)基因表达有上调的趋势(0.05≤P≤0.10),其余系膜脂基因表达2组间无显著差异(P>0.10)。
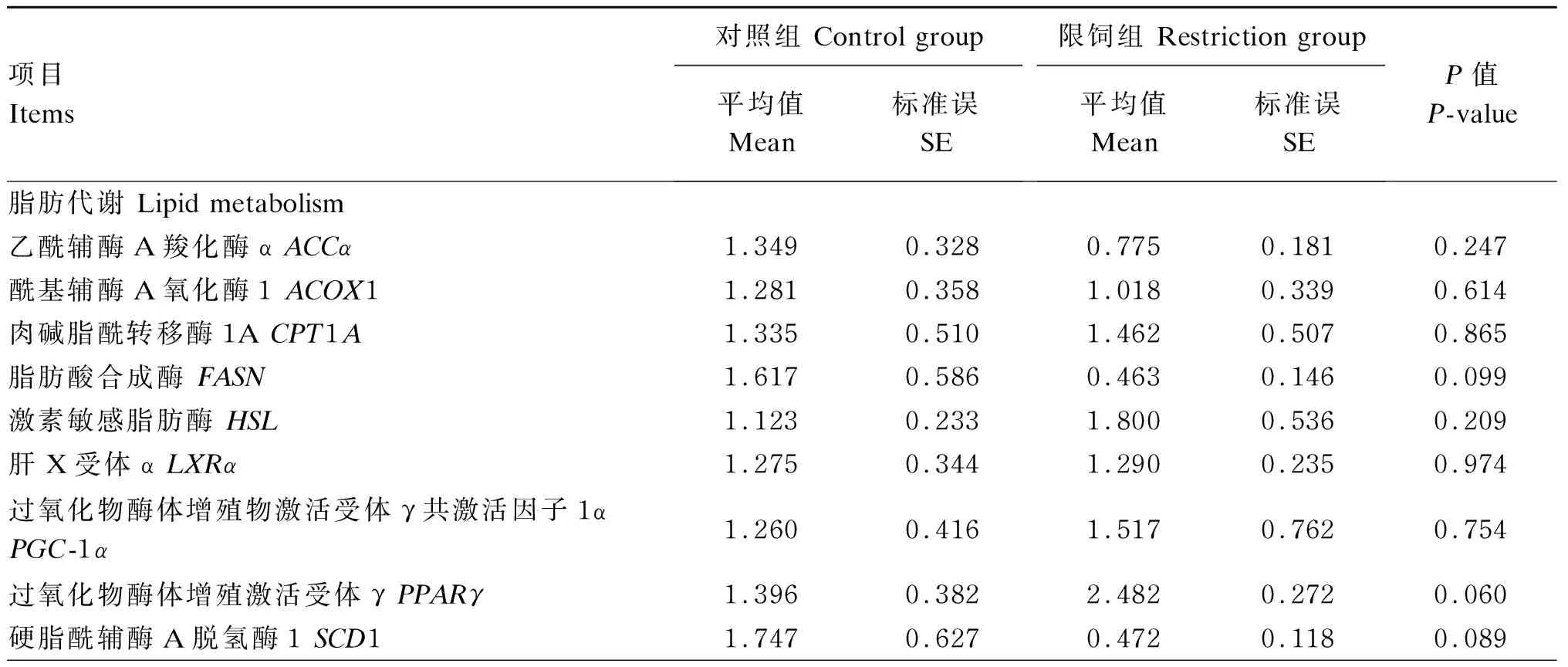
表8 限饲对妊娠中期母羊系膜脂脂肪代谢相关基因表达的影响

续表8项目Items对照组 Control group平均值Mean标准误SE限饲组 Restriction group平均值Mean标准误SEP值P-value能量状态感知 Energy state perception解偶联蛋白1 UCP11.1430.2561.3720.2850.561解偶联蛋白2 UCP21.2020.3140.4000.1340.053腺苷酸活化蛋白激酶的催化亚基α2 AMPKα21.2340.2221.2830.2870.893腺苷酸活化蛋白激酶的非催化亚基β1 AMPKβ11.1430.2251.1780.3210.926肝激酶B1 LKB11.0820.1863.0151.4580.277
由表9可知,与C组相比,R组肾周脂FASN和UCP2基因表达显著下调(P<0.05);肾周脂腺苷酸活化蛋白激酶的非催化亚基β1(AMPKβ1)基因表达显著上调(P<0.05),肾周脂肉碱脂酰转移酶1A(CPT1A)基因表达有上调的趋势(0.05≤P≤0.10),其余肾周脂基因表达2组间无显著差异(P>0.10)。
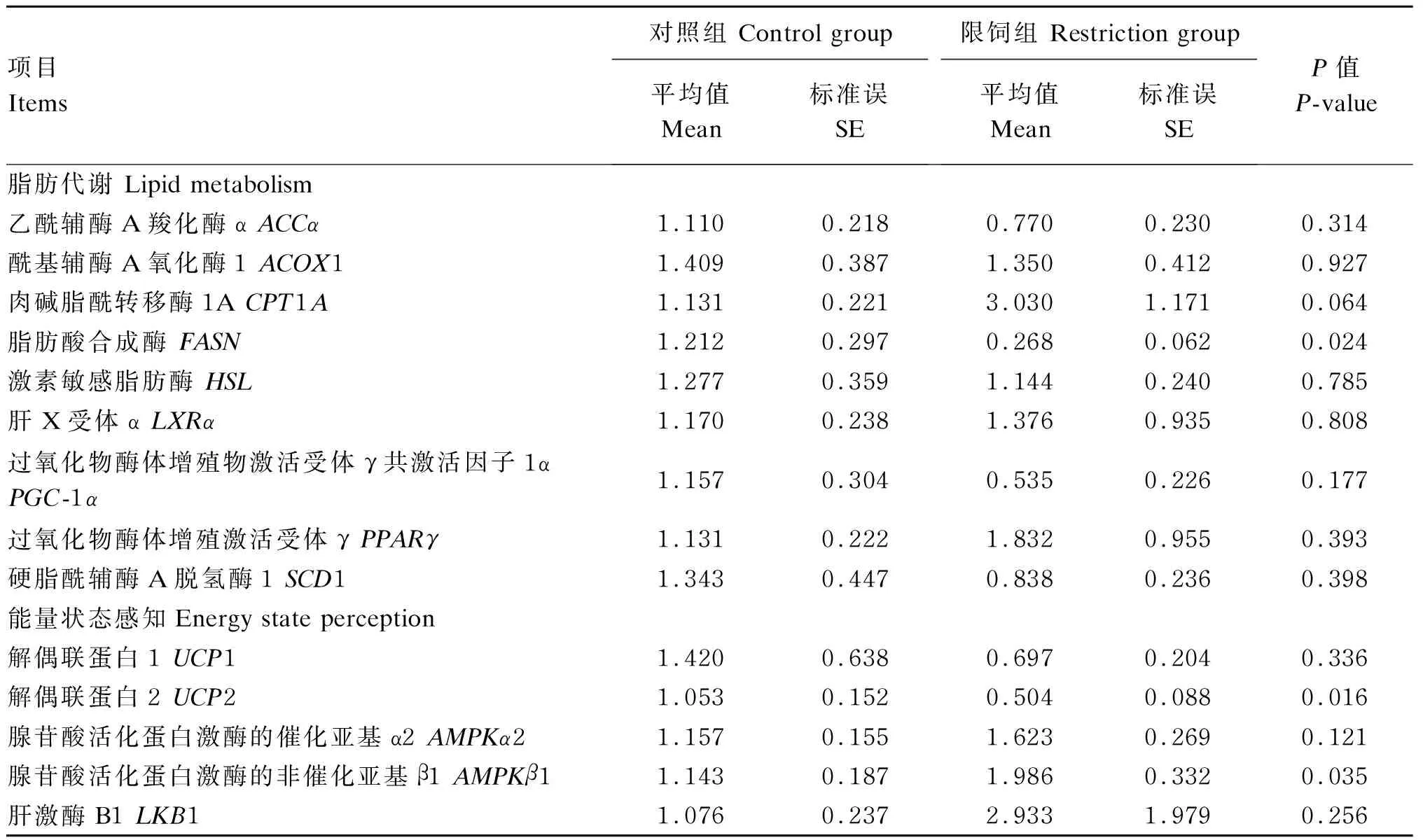
表9 限饲对妊娠中期母羊肾周脂脂肪代谢相关基因表达的影响
3 讨 论
饲粮营养供给不足时,妊娠母体会通过动员自身的能量储备——脂肪组织来维持自身和妊娠能量需求。VAT作为机体能量贮存和脂肪代谢系统的重要组成部分,可通过自身脂肪合成代谢和分解代谢调控来影响血液中脂肪代谢物质的浓度[12],进而影响机体脂肪代谢和能量代谢,以维持机体脂肪代谢和能量代谢的相对平衡。
3.1 限饲对母羊血液生化指标的影响
胰高血糖素是机体能量代谢重要调节激素,其可通过促进糖异生和糖原分解,降低脂肪酸合成和促进脂肪酸分解释放进入血液循环,以便机体组织氧化供能等来维持机体血液GLU和能量稳衡[13]。胰高血糖素的升高会使机体从VAT动员分解更多的FFA进入血液来弥补机体能量负平衡[12]。LEP和ADP是脂肪组织分泌的调控脂肪代谢、脂质稳态和能量代谢的重要调控因子[14-17],低营养条件和血液FFA的增加都会抑制LEP的分泌[18],本试验中R组母羊血液胰高血糖素和FFA含量升高,血液LEP和ADP含量降低,血液GLU含量相对稳定。这说明限饲条件下母体通过升高血液胰高血糖素含量、降低LEP和ADP含量来调整机体脂肪代谢,增强脂肪动员和增加对血液中脂肪分解产物FFA的利用来补偿机体能量不足,以维持血液GLU的相对稳定,从而适应营养限制环境保证妊娠的进行,这与Haghiac等[19]研究结果一致。
3.2 限饲对母羊VAT脂肪酸组成的影响
VAT脂肪酸组成主要受机体内脂肪合成代谢和分解代谢影响,妊娠动物脂肪酸组成对其繁殖性能、乳脂含量和能量代谢有重要意义。本试验中,限饲引起VAT脂肪酸组成发生明显变化,饱和脂肪酸豆蔻酸和不饱和脂肪酸(C18和C20为主)在不同VAT部位含量均降低。反刍动物中,中短链脂肪酸主要来自于乙酸和丁酸的从头合成,PUFA主要来自于瘤胃微生物生物转化和饲粮沉积[20-22]。这提示R组VAT脂肪酸变化的可能原因如下:一方面来源下降,限饲造成脂肪酸合成前体物乙酸和丁酸的来源减少,导致合成脂肪酸的微生物减少[23],同时饲粮来源PUFA也减少;另一方面去路增加,饲粮供能不足导致妊娠母羊脂肪动员,长链脂肪分解和脂肪酸氧化供能增加。2方面共同作用,导致了VAT脂肪酸组成的变化。
3.3 限饲对母羊VAT脂肪代谢相关基因表达的影响
机体营养状态可通过影响与脂肪代谢相关基因的表达来调节脂肪组织脂肪合成、脂肪动员和能量状态感知的功能[24],进而调控机体脂肪代谢和能量代谢。本试验主要研究限饲对VAT脂肪合成、脂肪动员及能量状态感知关键基因表达的影响。
FASN、SCD1、ACCa、肝X受体α(LXRα)和PPARγ是参与脂肪从头合成、脂质储存、脂肪酸稳态和脂肪细胞分化调控的关键基因[24-25],其中FASN主要调控脂肪酸从头合成,影响脂肪沉积[26-27];SCD1参与催化不饱和脂肪酸合成及饱和脂肪酸去饱和化[28]。本试验中,与C组相比,R组VAT中,FASN在肾周脂和系膜脂、SCD1在系膜脂均不同程度下调,表明限饲引起的营养不足导致肾周脂和系膜脂中长链脂肪酸从头合成能力减弱。VAT亚油酸、α-亚麻酸、二十碳烯酸等脂肪酸含量降低印证了基因表达的结果。
激素敏感脂肪酶(HSL)、CPT1A、酰基辅酶A氧化酶1(ACOX1)和过氧化物酶体增殖物激活受体γ共激活因子1α(PGC-1α)是调控脂肪动员、脂肪酸氧化、细胞脂质摄取及分解和线粒体合成及产能的关键调控基因[29-31],尤其CPT1A是长链脂肪酸β氧化早期步骤的关键调控因子,参与调节组织中脂肪酸氧化速率[32-33]。本试验中,R组VAT,肾周脂CPT1A有上调趋势,表明肾周脂中长链脂肪酸氧化速率加快,这与肾周脂长链脂肪酸含量降低的结果一致,提示母体可能通过此途径来补偿营养限制造成的机体能量短缺。
AMPK通路能感应机体能量需求状况,调节机体产能和耗能代谢,被称为细胞“代谢感受器”和“能量总开关”,是GLU代谢、脂质代谢和脂肪酸组成的中心调控因子[24],其还能与肝激酶B1(LKB1)互作维持脂质和能量平衡[34]。通路中AMPKα2和AMPKβ1分别编码α2和β1催化亚基。本试验中,R组VAT中,肾周脂AMPKβ1基因表达上调,网膜脂AMPKα2基因表达有上调趋势,表明限饲促进肾周脂和网膜脂能量感知开关基因的表达。解偶联蛋白(UCP)家族与机体能量代谢平衡、脂肪沉积、产热等密切相关,其中解偶联蛋白1(UCP1)主要调控棕色脂肪组织非颤抖性产热;UCP2主要通过上调解偶联作用来调控基础能量代谢,其还能影响FASN基因表达来调节脂质合成[35-36]。本试验中,R组VAT中,UCP2基因在肾周脂表达下调,在系膜脂也有下调的趋势,提示限饲使母体基础能量代谢水平趋于下降,有利于机体适应限饲引起的能量不足,这可以理解为母体适应外界不良环境的一种自我保护。De Queiroz等[37]研究表明,UCP2基因表达与脂肪组织脂肪酸含量呈正相关,脂肪FASN和UCP2基因表达变化也有同步性[35]。本试验UCP2、FASN基因表达变化及脂肪酸含量变化与上述研究结果一致。综上所述,限饲上调了母体VAT能量感知开关,并使母体的基础能量代谢水平有降低的趋势。
此外,本试验发现网膜脂和肾周脂脂肪酸组成受限饲影响程度大于系膜脂;限饲对肾周脂脂肪代谢基因表达的影响程度明显大于网膜脂和系膜脂。这提示不同VAT部位对限饲的响应强度存在差异性,从强到弱依次为肾周脂、网膜脂和系膜脂,推测这可能与饥饿-下丘脑-肾上腺轴对营养限制的快速感知和响应路径有关,具体原因有待深入研究。
4 结 论
妊娠中期限饲导致母羊血液中脂肪代谢调控因子(LEP和ADP)含量下降,脂肪代谢产物(FFA)含量上升;VAT脂肪酸组成改变(PUFA下降);并经下调脂肪合成基因(FASN和SCD1)、上调脂肪动员基因(CPT1A)及下调基础能量代谢基因(UCP2)表达的途径,使母体VAT脂肪合成作用减弱、脂质分解和脂肪酸氧化增强来调节机体能量稳态。
[2] ROOS S,LAGERLÖF O,WENNERGREN M,et al.Regulation of amino acid transporters by glucose and growth factors in cultured primary human trophoblast cells is mediated by mTOR signaling[J].American Journal of Physiology Cell Physiology,2009,297(3):C723-C731.
[3] ROSARIO F J,JANSSON N,KANAI Y,et al.Maternal protein restriction in the rat inhibits placental insulin,mTOR,and STAT3 signaling and down-regulates placental amino acid transporters[J].Endocrinology,2011,152(3):1119-1129.
[4] BARCROFT J.Researches on pre-natal life[M].Oxford:Blackwell Scientific Publications,1946.
[5] VIRTUE S,VIDAL-PUIG A.Adipose tissue expandability,lipotoxicity and the metabolic syndrome—an allostatic perspective[J].Genes & Nutrition,2007,2(1):41-45.
[6] NIELSEN M O,KONGSTED A H,THYGESEN M P,et al.Late gestation undernutrition can predispose for visceral adiposity by altering fat distribution patterns and increasing the preference for a high-fat diet in early postnatal life[J].British Journal of Nutrition,2013,109(11):2098-2110.
[7] DE BLASIO M J,GATFORD K L,ROBINSON J S,et al.Placental restriction of fetal growth reduces size at birth and alters postnatal growth,feeding activity,and adiposity in the young lamb[J].American Journal of Physiology:Regulatory,Integrative and Comparative Physiology,2007,292(2):R875-R886.
[8] KHANAL P,HUSTED S V,AXEL A M,et al.Late gestation over-and undernutrition predispose for visceral adiposity in response to a post-natal obesogenic diet,but with differential impacts on glucose-insulin adaptations during fasting in lambs[J].Acta Physiologica,2013,210(1):110-126.
[9] 张宏福.动物营养参数与饲养标准[M].2版.北京:中国农业出版社,2010.
[10] 张丽英.饲料分析及饲料质量检测技术[M].2版.北京:中国农业大学出版社,2003.
[11] ICHIHARA K,SHIBAHARA A,YAMAMOTO K,et al.An improved method for rapid analysis of the fatty acids of glycerolipids[J].Lipids,1996,31(5):535-539.
[12] WAJCHENBERG B L.Subcutaneous and visceral adipose tissue:their relation to the metabolic syndrome[J].Endocrine Reviews,2000,21(6):697-738.
[13] HABEGGER K M,HEPPNER K M,GEARY N,et al.The metabolic actions of glucagon revisited[J].Nature Reviews Endocrinology,2010,6(12):689-697.
[14] EHRHARDT R A,SLEPETIS R M,BELL A W,et al.Maternal leptin is elevated during pregnancy in sheep[J].Domest Anim Endocrinol,2001,21(2):85-96.
[16] VASSEUR F,LEPRTRE F,LACQUEMANT C,et al.The genetics of adiponectin[J].Current Diabetes Reports,2003,3(2):151-158.
[17] DIEZ J J,IGLESIAS P.The role of the novel adipocyte-derived hormone adiponectin in human disease[J].European Journal of Endocrinology,2003,148(3):293-300.
[18] HARRIS R B S,APOLZAN J W.Changes in glucose tolerance and leptin responsiveness of rats offered a choice of lard,sucrose,and chow[J].American Journal of Physiology:Regulatory, Integrative and Comparative Physiology,2012,302(11):R1327-R1339.
[19] HAGHIAC M,BASU S,PRESLEY L,et al.Patterns of adiponectin expression in term pregnancy:impact of obesity[J].The Journal of Clinical Endocrinology & Metabolism,2014,99(9):3427-3434.
[20] 茅慧玲,刘建新.反刍动物肌肉脂肪酸营养调控研究进展[J].饲料工业,2010,31(23):30-34.
[21] BANSKALIEVA V V,SAHLU T,GOETSCH A L.Fatty acid composition of goat muscles and fat depots:a review[J].Small Ruminant Research,2000,37(3):255-268.
[22] 双金,敖力格日玛,敖长金.苏尼特羊体脂脂肪酸组成的研究[J].畜牧兽医学报,2015,46(8):1363-1374.
[23] DERVISHI E,SERRANO C,JOY M,et al.Effect of the feeding system on the fatty acid composition,expression of the Δ9-desaturase,peroxisome proliferator-activated receptor alpha,gamma,and sterol regulatory element binding protein 1 genes in the semitendinous muscle of light lambs of theRasaaragonesabreed[J].BMC Veterinary Research,2010,6:40.
[24] DERVISHI E,SERRANO C,JOY M,et al.The effect of feeding system in the expression of genes related with fat metabolism in semitendinous muscle in sheep[J].Meat Science,2011,89(1):91-97.
[25] 蒋金航,马云,王新庄.PPARγ基因调控脂肪细胞分化的研究进展[J].中国畜牧杂志,2014,50(9):91-95.
[26] YEON S H,LEE S H,CHOI B H,et al.Genetic variation ofFASNis associated with fatty acid composition of Hanwoo[J].Meat Science,2013,94(1):133-138.
[27] MENENDEZ J A,LUPU R.Fatty acid synthase-catalyzeddenovofatty acid biosynthesis:from anabolic-energy-storage pathway in normal tissues to jack-of-all-trades in cancer cells[J].Archivum Immunologiae et Therapiae Experimentalis,2004,52(6):414-426.
[28] PATON C M,NTAMBI J M.Biochemical and physiological function of stearoyl-CoA desaturase[J].American Journal of Physiology Endocrinology and Metabolism,2009,297(1):E28-E37.
[29] HOLM C.Molecular mechanisms regulating hormone-sensitive lipase and lipolysis[J].Biochemical Society Transactions,2003,31(6):1120-1124.
[30] PUIGSERVER P,WU Z D,PARK C W,et al.A cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis[J].Cell,1998,92(6):829-839.
[31] 孙亮,朱小泉,王沥,等.核辅激活因子PGC-1表达的分子调控机制[J].中国生物化学与分子生物学报,2005,21(4):431-439.
[32] 张艳芳.CPT1基因对猪脂肪沉积的影响及其调控机制的研究[D].博士学位论文.杭州:浙江大学,2010.
[33] BARTELDS B,TAKENS J,SMID G B,et al.Myocardial carnitine palmitoyltransferase Ⅰ expression and long-chain fatty acid oxidation in fetal and newborn lambs[J].American Journal of Physiology:Heart and Circulatory Physiology,2004,286(6):H2243-H2248.
[34] MCINNES K J,BROWN K A,HUNGER N I,et al.Regulation ofLKB1 expression by sex hormones in adipocytes[J].International Journal of Obesity,2012,36(7):982-985.
[35] WU B,DU Y,LIU C,et al.Effect of repeated fasting/refeeding on body weight control and energy balance regulation in rats[J].Journal of Hygiene Research,2010,39(5):601-605.
[36] MOON J S,LEE S,PARK M A,et al.UCP2-induced fatty acid synthase promotes NLRP3 inflammasome activation during sepsis[J].Journal of Clinical Investigation,2015,125(2):665-680.
[37] DE QUEIROZ K B,GUIMARES J B,COIMBRA C C,et al.Endurance training increases leptin expression in the retroperitoneal adipose tissue of rats fed with a high-sugar diet[J].Lipids,2014,49(1):85-96.
猜你喜欢
昆明医科大学学报(2022年1期)2022-02-28
现代畜牧科技(2021年10期)2021-11-19
现代畜牧科技(2021年8期)2021-10-13
中老年保健(2021年9期)2021-08-24
昆明医科大学学报(2021年8期)2021-08-13
现代畜牧科技(2021年7期)2021-07-28
现代畜牧科技(2021年6期)2021-07-16
现代畜牧科技(2021年2期)2021-03-19
临床超声医学杂志(2020年10期)2020-12-13
河南医学研究(2014年2期)2014-02-27
