
功能性低聚糖及其缓解机体过敏反应的研究进展
2018-06-14 00:59常美楠朱亚如秦贵信
动物营养学报 2018年6期
常美楠 朱亚如 赵 元 秦贵信
(吉林农业大学动物科技学院,动物生产及产品质量安全教育部重点实验室,吉林省动物营养与 饲料科学重点实验室,长春 130118)
近年来,食物和饲粮过敏现象普遍流行,严重威胁人和动物的健康。引起过敏的过敏原有奶、蛋、坚果类、海鲜[1]、小麦蛋白[2]、大豆蛋白[3]、卵清蛋白(OVA)[4]等。研究表明,人和动物在口服或接触了这些过敏原后,既有特异性抗原抗体间的速发型变态反应,又有T淋巴细胞介导的迟发型过敏反应,其中速发型变态反应发病率较高,主要由免疫球蛋白E(IgE)介导,导致肥大细胞和嗜碱性粒细胞释放组胺等活性物质,使人和动物在口服或接触了这些过敏原后,产生皮肤出现红斑、胃肠道不适、消化力下降、呕吐、腹泻等症状,甚至危及生命,严重影响了人类的健康和畜牧业的生产效益[5]。
因而,如何消除或缓解过敏反应的发生,是人类食品及畜禽饲料安全亟待解决的问题。以往食品和饲料原料采用加工处理技术来钝化其中的致敏原,如热处理、高压膨化处理[6]、发酵处理[7]等。此外,采用新型天然抗过敏活性物质来预防和治疗过敏反应也是有效缓解过敏的一种方式,且逐渐成为一个新的研究热点。目前,已开展的相关研究有硫辛酸[8]、中草药等植物及其提取物[9]、益生菌[10]、益生元[11]等。
功能性低聚糖是化学益生元,能改变宿主肠道内的菌群组成,增殖双歧杆菌,优化肠道菌群,是一种活性膳食补充剂,现已作为一类食品和饲料添加剂应用于功能性保健食品和动物饲料。适量的功能性低聚糖能提高机体的免疫和屏障功能,但仅有一些关于功能性低聚糖缓解食物和饲粮过敏反应的研究,所以探究功能性低聚糖的抗过敏功效,进一步探究其抗过敏机理,对于开发新型食品和饲料添加剂具有极其重要的意义。
1 功能性低聚糖的分类与组成
功能性低聚糖,又称功能性寡糖,对人和动物的免疫系统和肠道健康都有益处,如抗癌、调节免疫活性等。功能性低聚糖广泛分布于植物、藻类、细菌和高等真菌中,现已应用于许多领域,如营养、医药、饲料、化妆品、免疫刺激剂和益生元成分等[12]。应用较多的功能性低聚糖包括低聚果糖(FOS)、低聚半乳糖(GOS)、甘露寡糖(MOS)、低聚麦芽糖、低聚异麦芽糖、大豆低聚糖、水苏糖、棉子糖、低聚壳聚糖、龙须菜寡糖等,另外还有阿拉伯低聚糖、低聚乳果糖、低聚葡萄糖、低聚龙胆糖等。功能性低聚糖的化学组成见表1。
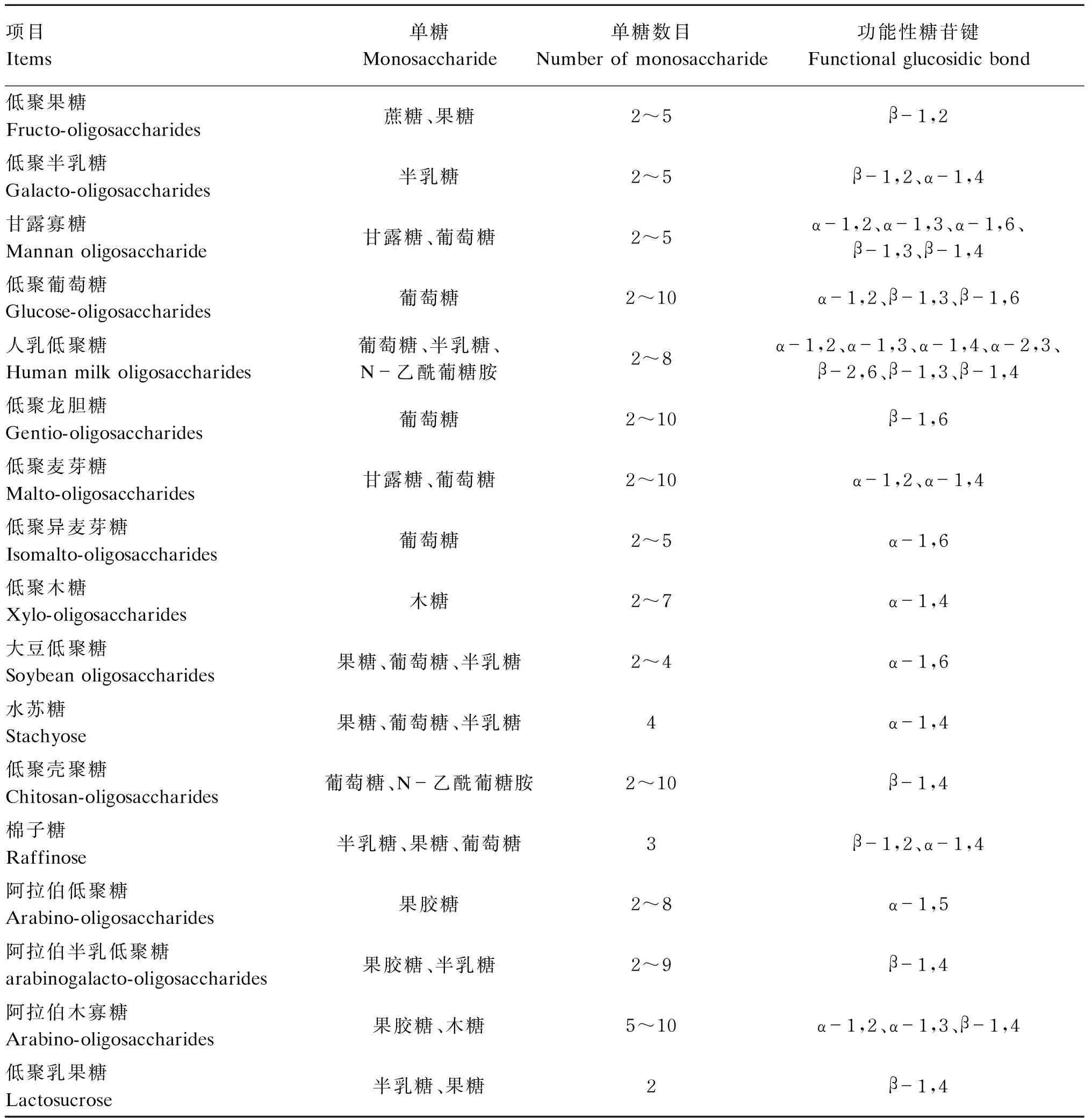
表1 功能性低聚糖的化学组成
2 功能性低聚糖的生理功能
2.1 促进益生菌增殖
功能性低聚糖可改善肠道内菌群的结构和组成,并选择性地促进双歧杆菌等益生菌的增殖。双歧杆菌对机体具有很好的保健作用,是肠道唯一一类不产生毒素、无致病性的功能性有益微生物。人和动物体内没有水解功能性低聚糖的酶类,它们可直接进入大肠内优先被双歧杆菌利用,显著促进机体内双歧杆菌的增殖[12]。功能性低聚糖用于动物饲粮中可抑制病原菌的定植和生长,增加肠道菌群的多样性[16],提高畜禽的生长速度、生产性能和免疫机能[17]。
2.2 提高免疫力
功能性低聚糖可作为免疫刺激因子,促进肠道免疫细胞的生长,提高血清免疫球蛋白A(IgA)、免疫球蛋白M(IgM)、免疫球蛋白G(IgG)和白细胞介素-2(IL-2)的含量,从而增加动物体液及细胞免疫功能。低聚糖进入肠道后会竞争性地和病原菌表面的外源凝集素结合,阻止病原菌在肠上皮上黏附,同时低聚糖又有促进肠道蠕动的作用,使病原菌随粪便排出,减少对肠道的损害[18]。此外,低聚糖可促进肠道内双歧杆菌等有益菌大量增殖,双歧杆菌具有免疫激活作用,可增强巨噬细胞、淋巴细胞的吞噬活性,诱导细胞因子的产生;这些有益菌在肠道内会产生乙酸、丁酸等短链脂肪酸(SCFAs),降低肠道pH,酸化肠道内环境,缩短病原物质与肠黏膜的接触时间,从而调节机体的免疫应答[19]。
2.3 促进矿物质和营养物质的吸收,润肠通便
功能性低聚糖到达机体后被微生物发酵分解,释放出矿物质离子,并且可促进钙离子、铁离子的吸收和提高钙离子、镁离子的溶解度,有利于盲肠、结肠等对矿物质元素的吸收[20-21]。微生物的发酵过程还会产生机体所需的维生素,促进和改善脂质、蛋白质等营养物质的吸收[22]。另外功能性低聚糖被双歧杆菌利用后,可产生大量的SCFAs,有助于增进肠道蠕动,起到润肠通便的作用[23]。作为饲料添加剂,功能性低聚糖具有抗生素的功能,可提高动物生长、育肥和增加泌乳量,还可降低粪便中氨气的含量。
3 功能性低聚糖缓解过敏反应的研究
在动物模型方面,关于功能性低聚糖缓解过敏反应的研究较少,大多数都是预防的案例,治疗的很少。其中研究较多的低聚糖有FOS、GOS、GOS/菊粉混合益生元以及MOS等。低聚糖主要通过以下作用功能来实现缓解过敏的目的:调节机体免疫力、增强肠黏膜屏障功能、改善肠道菌群平衡。
3.1 调节机体免疫力
早期幼龄动物免疫系统和肠道菌群系统尚未发育完善,存在复杂性和多样性,易受到外界过敏原的刺激而产生过敏型疾病。正常机体内辅助性T淋巴细胞(Th细胞)亚群Th1和Th2细胞功能处于动态平衡状态,维持机体的细胞免疫和体液免疫功能[24]。Th1细胞主要分泌IL-2、干扰素-γ(IFN-γ)、干扰素-α(IFN-α)、肿瘤坏死因子-β(TNF-β)等,参与有关炎症的免疫应答。Th2细胞主要分泌白细胞介素-4(IL-4)、白细胞介素-5(IL-5)、白细胞介素-6(IL-6)和白细胞介素-10(IL-10)等细胞因子,刺激B淋巴细胞增殖并生成大量IgE和IgG抗体[25]。在由食物抗原引起的人和动物过敏反应中速发型变态反应较多,表现为过敏体内的Th1/Th2免疫失衡,因此Th1/Th2平衡在过敏反应中有着至关重要的作用。
研究发现,功能性低聚糖可通过诱导免疫相关因子,继而影响免疫平衡达到预防和治疗过敏的目的。GOS/菊粉混合益生元能使围产期和断奶后期小鼠IFN-γ、TGF-β和IgA含量升高,小鼠幼鼠小肠内IL-10、IgA和IgG2a表达量显著升高[11,26];GOS/菊粉混合益生元也能增加粪便中双歧杆菌的数量,使血清IgG1、IgG2、IgG3含量大幅上升,显著降低婴儿过敏性疾病的发病率,并且这个影响具有长效性[27-29]。龙须菜寡糖可使致敏小鼠血清特异性IgE和IgG1含量显著降低,血清IgG2a含量显著升高,抑制Th2细胞因子[IL-4、IL-5、白细胞介素-(IL-13)]含量,促进Th1细胞因子(IFN-γ)的表达[30]。MOS可能与肠黏膜上皮细胞的受体结合从而刺激淋巴细胞,促进血清IgA、IgG和IL-10释放,降低血清TNF-α含量,进而调节畜禽机体免疫力[31-33]。这些免疫应答证明功能性低聚糖能有效调节机体免疫球蛋白和一些细胞因子,使Th1/Th2细胞重新处于相对平衡状态,对于过敏反应有预防和治疗功效,尤其是在妊娠期、分娩期和哺乳期这些重要时期,对免疫机制有显著的调节作用。
功能性低聚糖还能通过改善肥大细胞脱颗粒现象来提高机体免疫力。Fujitani等[34]研究发现,FOS可降低因OVA过敏小鼠的十二指肠肥大细胞的数量和水肿率。Vonk等[35]用FOS饲喂牛奶蛋白乳清致敏的小鼠,发现急性过敏反应和肥大细胞的脱颗粒得到有效缓解,防止特异性IgE含量的升高,降低T细胞应答反应,最终提高小鼠的口服免疫耐受性。另外功能性低聚糖可通过增加血清中半乳凝集素9的含量而影响免疫系统[36]。
3.2 增强肠黏膜屏障功能
肠黏膜屏障主要由机械屏障、化学屏障、免疫屏障和生物屏障4部分构成,其中任何一部分损伤都会危害机体健康[37]。肠黏膜屏障功能使肠道内的细菌和内毒素等物质被局限于肠腔内而不会转移到外周脏器。肠上皮细胞通透性的增加和破坏会将黏膜暴露于肠腔食物或细菌的大量抗原中,刺激免疫兴奋,导致肠道炎症的发生,从而引起多种胃肠道疾病以及过敏反应的发生。研究表明,由于食物和饲粮过敏引起的多种胃肠道疾病和肠道炎症的发生都与肠黏膜屏障的损伤有关[38]。低聚糖作为一种微生态制剂,可促进肠内有益菌的增殖,改善肠壁的通透性,有利于维护肠黏膜屏障功能。
功能性低聚糖通过增强肠黏膜屏障功能预防和治疗过敏反应的形式主要表现在以下3个方面:肠道黏膜形态、黏膜通透性以及紧密连接蛋白。寡糖益生元、益生菌可通过改善肠道微生物菌群平衡,影响肉仔鸡和仔猪肠道黏膜形态和结构,对空肠和结肠的黏膜完整性有一定的保护作用[39-40]。血浆二胺氧化酶(DAO)是所有哺乳动物肠黏膜上皮细胞胞浆中活性较高的酶,在其他组织中活性较低。若肠黏膜系统受损,DAO就会进入外周血液,导致肠黏膜通透性增加。GOS可通过影响DAO活性影响肠黏膜通透性。此外,GOS可能通过调节转录环节mRNA水平,增加跨膜蛋白(occludins)的含量,进而改善紧密连接,维护肠黏膜屏障功能[41]。功能性低聚糖可通过紧密连接蛋白诱导肠上皮细胞产生保护性物质,如热休克蛋白(HSP)、上皮黏蛋白(MUC),分泌SCFAs来调节肠黏膜屏障功能。Gourbeyre等[26]研究发现,饲喂GOS/菊粉混合益生元的小鼠盲肠内容物中丙酸盐含量上升,空肠中HSP-25、闭锁小带蛋白1(ZO-1)和黏蛋白2(MUC-2)含量增加。
3.3 改善肠道菌群平衡
动物肠道内的微生物菌群是一个动态的微生态系统,防止外源的食物抗原和病原菌入侵机体,参与肠上皮细胞的新陈代谢以及免疫机制的调控,有利于动物对饲料中营养物质的消化吸收。低聚糖益生元作为肠内益生菌的专属食物,进入机体后定向的被有益菌利用,增加肠道内有益菌的数量和种类。肠道内有益菌占据优势后就会抑制有害菌群的定植,从而稳定肠道内环境,保护肠道健康,达到预防和治疗过敏的目的。Watanabe等[42]给Balb/c小鼠饲喂含有FOS的饲粮后,会使小鼠肠道内的双歧杆菌增殖,降低2,4-二硝基氟苯引起的接触性超敏反应现象。外源添加低聚糖(GOS、FOS)和有益菌(乳酸杆菌、双歧杆菌)可显著增加禽类肠道内微生物的代谢活性,降低沙门氏菌等有害菌的定植[43-44]。GOS能有效改善仔猪肠道菌群结构和微生物平衡,调节肠道防御体系[40]。
微生物菌群对免疫应答调节和肠道屏障功能都有重要作用,许多有益菌可与小肠上皮细胞、树突细胞、Th1细胞、Th2细胞相互作用,通过刺激B淋巴细胞产生IgA,IgA再与抗原结合,抑制抗原进入上皮细胞,从而达到调节免疫应答的目的[45]。功能性低聚糖经微生物发酵后,形成的SCFAs参与机体的免疫应答和屏障保护,可间接地预防过敏性疾病的发生。有研究显示,SCFAs可降低促炎症细胞因子含量,包括白细胞介素-10β(IL-10β)、IL-6、TNF-α,提高抗炎细胞因子IL-10含量[46-47];还能诱导肠上皮细胞分泌白细胞介素-18(IL-18)、MUC和抗菌肽,加强紧密连接结构,维持肠黏膜屏障的完整性[48-49]。丁酸盐可作为识别组蛋白修饰乙酰化的调节器,在肠上皮细胞模型中通过紧密连接蛋白提高小肠屏障功能[50]。婴儿食用含低聚糖的奶粉后,粪便中葡萄球菌和链球菌减少,菌群(尤其是双歧杆菌)丰富度和多样性增加,粪便中乙酸、丙酸、丁酸含量升高,湿疹发病率降低[51]。
4 小 结
综上所述,过敏反应的发生导致机体肠道结构破坏,菌群紊乱,肠道屏障系统和免疫机能严重破坏,功能性低聚糖作为一类对机体无毒副作用的益生元,不但能改变肠道菌群的丰富度和多样性,而且对机体的免疫机制和肠道屏障功能的提高有很大的帮助,因此对过敏反应有一定的预防和治疗效果。功能性低聚糖在食品和动物饲料中有广阔的应用前景,目前多种低聚糖都已被作为食品和饲料添加剂,用于提高人和动物的生长性能,但在抗过敏方面,研究更多的是小鼠和人,畜禽领域相对较少,因此未来此方面仍有待进一步探讨研究。已有的食物过敏方面的研究主要从提高免疫力、增强肠道屏障功能以及改善菌群平衡来实现,但由于过敏反应发生和缓解机制比较复杂,目前的研究尚不完善,过敏仍然是困扰人类和动物健康的一大难题。
[1] SAVAGE J,JOHNS C B.Food allergy:epidemiology and natural history[J].Immunology and Allergy Clinics of North America,2015,35(1):45-59.
[2] KIM J H,JEUN E J,HONG C P,et al.Extracellular vesicle-derived protein fromBifidobacteriumlongumalleviates food allergy through suppression of mast cell suppression[J].Journal of Allergy and Clinical Immunology,2016,137(2):507-516.e8.
[3] SUN P,LI D F,DONG B,et al.Vitamin C:an immunomodulator that attenuates anaphylactic reactions to soybean glycinin hypersensitivity in a swine model[J].Food Chemistry,2009,113(4):914-918.
[4] FANG S F,ZHANG Y,ZHANG Y,et al.LactobacillusrhamnosusGG improves symptoms and its mechnism in mice with ovalbumin-induced food allergy[J].Chinese Journal of Cellular and Molecular Immunology,2017,33(5):597-600.
[5] 孙鹏.大豆抗原蛋白Glycinin诱发仔猪过敏反应的机理及其缓解机制的研究[D].博士学位论文.北京:中国农业大学,2009.
[6] 刘丹丹,赵元,张诗尧.大豆抗原蛋白对肠道致敏性的研究进展[J].中国畜牧杂志,2015,51(5):77-81.
[7] 张建营,秦贵信,鲍男.发酵豆粕对反刍动物的影响研究[J].黑龙江畜牧兽医,2015(13):111-113.
[8] CHO Y S,LEE J,LEE T H,et al.α-Lipoic acid inhibits airway inflammation and hyperresponsiveness in a mouse model of asthma[J].Journal of Allergy and Clinical Immunology,2004,114(2):429-435.
[9] WANG J L,PATIL S P,YANG N,et al.Safety,tolerability,and immunologic effects of a food allergy herbal formula in food allergic individuals:a randomized,double-blinded,placebo-controlled,dose escalation,phase 1 study[J].Annals of Allergy, Asthma & Immunology,2010,105(1):75-84.
[10] 季晓梅,李艾黎,姚蕾,等.嗜酸乳杆菌缓解牛乳过敏的研究[J].中国食品学报,2016,16(7):33-38.
[11] GOURBEYRE P,DESBUARDS N,GRÉMY G,et al.Perinatal and postweaning exposure to galactooligosaccharides/inulin prebiotics induced biomarkers linked to tolerance mechanism in a mouse model of strong allergic sensitization[J].Journal of Agricultural and Food Chemistry,2013,61(26):6311-6320.
[12] ZHAO C,WU Y J,LIU X Y,et al.Functional properties,structural studies and chemo-enzymatic synthesis of oligosaccharides[J].Trends in Food Science & Technology,2017,66:135-145.
[13] 单黎然,龚月桦,贾建光,等.4种重要功能性低聚糖的研究进展[J].西北农林科技大学(自然科学版),2006,34(7):96-100.
[14] 魏红军.功能性低聚糖的生理功能及其在食品中的应用[J].农产品加工(学刊),2011(9):127-128.
[15] 李鑫,张梁,羊梦诗,等.低聚壳聚糖几何结构和物理化学属性的理论研究[J].物理学报,2014,63(7):076102.
[16] ZHOU X L,KONG X F,LIAN G Q,et al.Dietary supplementation with soybean oligosaccharides increases short-chain fatty acids but decreases protein-derived catabolites in the intestinal luminal content of weanedHuanjiangmini-piglets[J].Nutrition Research,2014,34(9):780-788.
[17] SELIM K M,REDA R M.Beta-glucans and mannan oligosaccharides enhance growth and immunity in Nile Tilapia[J].North American Journal of Aquaculture,2015,77(1):22-30.
[18] 李涵.低聚糖在母猪繁殖和仔猪生产的应用[J].中国动物保健,2017,19(11):22-25.
[19] ATTIA Y A,AI-HAMID A E,ALLAKANY H F,et al.Supplementation of non-nutritive feed additive during days 21-42 of age following 3 weeks of feeding aflatoxin to broiler chickens[J].Journal of Applied Animal Research,2016,44(1):87-98.
[20] 王乃强,李国庆,刘辉,等.低聚果糖促进钙离子吸收的研究[J].精细与专用化学品,2013,21(5):20-23.
[21] 张丹凤.功能性低聚糖及其在乳制品中的应用[J].新疆畜牧业,2005(6):21-23.
[22] 刘花兰,姜竹茂,刘云国,等.功能性低聚糖的制备、功能及应用研究进展[J].中国食品添加剂,2015(12):158-166.
[23] 刘协,胡启之,李小宁,等.低聚果糖的润肠通便功能研究[J].江苏预防医学,2002,13(1):62-63.
[24] 刘力,张欢,马文军,等.马齿苋水煎剂对溃疡性结肠炎小鼠结肠组织中IL-8、IL-10表达的影响[J].辽宁中医药大学学报,2011,13(5):18-20.
[25] CANANI R B,COATANZO M D.Gut microbiota as potential therapeutic target for the treatment of cow’s milk allergy[J].Nutrients,2013,5(3):651-662.
[26] GOURBEYRE P,DESBUARDS N,GRÉMY G,et al.Exposure to a galactooligosaccharides/inulin prebiotic mix at different developmental time points differentially modulates immune responses in mice[J].Journal of Agricultural and Food Chemistry,2012,60(48):11942-11951.
[27] CUELLO-GARCIA C,FIOCCHI A,PAWANKAR R,et al.Prebiotics for the prevention of allergies:a systematic review and meta-analysis of randomized controlled trials[J].Clinical and Experimental Allergy,2017,47(11):1468-1477.
[28] MORO G,ARSLANOGLU S,STAHL B,et al.A mixture of prebiotic oligosaccharides reduces the incidence of atopic dermatitis during the first six months of age[J].Archives of Disease in Childhood,2006,91(10):814-819.
[29] ARSLANOGLU S,MORO G E,SCHMITT J,et al.Early dietary intervention with a mixture of prebiotic oligosaccharides reduces the incidence of allergic manifestations and infections during the first two years of life[J].Journal of Nutrition,2008,138(6):1091-1095.
[30] 时超岚,曾润颖,曹敏杰,等.龙须菜寡糖分离纯化和抗食物过敏功效的研究[J].集美大学学报(自然科学版),2015,20(1):14-22.
[31] 李玉欣.毕赤酵母甘露寡糖对猪生产性能和免疫性能的影响[D].博士学位论文.北京:中国农业大学,2015.
[32] CHE T M.Effects of mannan oligosaccharide on immune function and disease resistance in pigs[D].Ph.D.Thesis.Champaign:University of Illinois at Urbana,2010.
[33] 丁鹏,胡贵丽,刘勇强,等.酵母甘露寡糖对猪鸡生产性能和免疫功能的影响[J].湖南饲料,2017(5):24-30.
[34] SHINOBU F,KOZO U,TARO K,et al.Increased number of CCR4-positive cells in the duodenum of ovalbumin-induced food allergy model Nc/jic mice and antiallergic activity of fructooligosaccharides[J].Allergology International,2007,56(2):131-138.
[35] VONK M M,DIKS M A P,WAGENAAR L,et al.Improved efficacy of oral immunotherapy using non-digestible oligosaccharides in a murine cow’s milk allergy model:a potential role for foxp3+ regulatory t cells[J].Frontiers in Immunology,2017,8:1230.
[36] DE KIVIT S,SAELAND E,KRANEVELD A D,et al.Galectin-9 induced by dietary synbiotics is involved in suppression of allergic symptoms in mice and humans[J].Allergy,2012,67(3),343-352.
[37] 赵海君,钱易,崔毓桂,等.紧密连接的结构与功能[J].国际生殖健康/计划生育杂志,2013,32(5):384-386.
[38] FORBES E E,GROSCHWITZ K,ABONIA J P,et al.IL-9 and mast cell-mediated intestinal permeability predisposes to oral antigen hypersensitivity[J].Journal of Experimental Medicine,2008,205(4):897-913.
[39] AWAD W A,GHAREEB K,ABDEL-RAHEEM S,et al.Effects of dietary inclusion of probiotic and synbiotic on growth performance,organ weights,and intestinal histomorphology of broiler chickens[J].Poultry Science,2009,88(1):49-56.
[40] ALIZADEH A,AKBARI P,DIFILIPPO E,et al.The piglet as a model for studying dietary components in infant diets:effects of galacto-oligosaccharides on intestinal functions[J].British Journal of Nutrition,2016,115(4):605-618.
[41] 谢婷.益生元对CRS大鼠肠上皮紧密连接及屏障功能的影响[D].硕士学位论文.上海:第二军医大学,2007.
[42] WATANABE J,SASAJIMA N,ARAMAKIA A,et al.Consumption of fructo-oligosaccharide reduces 2,4-dinitrofluorobenzene-induced contact hypersensitivity in mice[J].British Journal of Nutrition,2008,100(2):339-346.
[43] TANNER S A,LACROIX C,DEL’HOMME C,et al.Effect ofBifidobacteriumthermophilumRBL67 and fructo-oligosaccharides on the gut microbiota in Göttingen minipigs[J].British Journal of Nutrition,2015,114(5):746-755.
[44] JUNG S J,HOUDE R,BAURHOO B,et al.Effects of galacto-oligosaccharides and a Bifidobacteria lactis-based probiotic strain on the growth performance and fecal microflora of broiler chickens[J].Poultry Science,2008,87(9):1694-1699.
[45] GOURBEYRE P,DENERY S,BODINIER M.Probiotics,prebiotics,and synbiotics:impact on the gut immune system and allergic reactions[J].Journal of Leukocyte Biology,2011,89(5):685-695.
[46] TEDELIND S,WESTBERG T,KJERRULF M,et al.Anti-inflammatory properties of the short-chain fatty acids acetate and propionate:a study with relevance to inflammatory bowel disease[J].World Journal of Gastroenterology,2007,13(20):2826-2832.
[47] LIU H,ZHANG J,ZHAGN S H,et al.Oral administration ofLactobacillusfermentumI5007 favors intestinal development and alters the intestinal microbiota in formula-fed piglets[J].Journal of Agricultural and Food Chemistry,2014,62(4):860-866.
[48] SUN M M,WU W,LIU Z J,et al.Microbiota metabolite short chain fatty acids,GPCR,and inflammatory bowel diseases[J].Journal of Gastroenterology,2017,52(1):1-8.
[49] 林日添,吴维,刘占举.短链脂肪酸对肠黏膜稳态免疫调节作用的研究进展[J].免疫学杂志,2017,33(10):900-904.
[50] CAVAGLIERI C R,NISHIYAMA A,FERNANDES L C,et al.Differential effects of short-chain fatty acids on proliferation and production of pro- and anti-inflammatory cytokines by cultured lymphocytes[J].Life Sciences,2003,73(13):1683-1690.
[51] WOPEREIS H,SIM K,SHAW A,et al.Intestinal microbiota in infants at high risk for allergy:effects of prebiotics and role in eczema development[J].Journal of Allergy and Clinical Immunology,2017,doi:10.1016/j.jaci.2017.05.054.
猜你喜欢
江苏安全生产(2022年8期)2022-11-01
食品科学(2022年13期)2022-07-29
小资CHIC!ELEGANCE(2021年36期)2021-10-15
艺术评鉴(2020年5期)2020-04-30
人大建设(2018年10期)2018-12-07
大豆科技(2016年6期)2016-03-11
应用化工(2014年11期)2014-08-16
中国药业(2014年19期)2014-05-17
食品工业科技(2014年13期)2014-03-11
食品科学(2013年17期)2013-03-11
