
湖南本土牡丹遗传多样性与亲缘关系的ISSR分析
2018-06-07 02:56钟丽凡吕长平许文婷陈海霞
江苏农业科学 2018年10期
钟丽凡, 吕长平, 许文婷, 陈海霞
(湖南农业大学园艺园林学院,湖南长沙 410128)
牡丹(PaeoniasuffruticosaAndr.)为芍药科(Paeniaceae)芍药属(PaeoniaL.)牡丹组(Sect.Moutan DC.)植物,是我国特有的植物种质资源[1-3]。我国牡丹品种多样,观赏牡丹分布地区以西北地区为主。湖南也是牡丹自然分布地区之一,其品种耐高温高湿、抗病虫害,但花色单一、花型简单缺乏观赏价值。因此,如何将观赏牡丹与湖南牡丹的优良特性结合成为当前推广湖南牡丹的首要任务。
目前牡丹育种工作主要以人工定向杂交选择为主[4-5]。但现有湖南牡丹品种大多数亲缘不清,对湖南具体牡丹品种数的统计比较笼统,存在育种工作效率低下、品种出新率低的问题。因此,研究清楚湖南本土牡丹种质资源亲缘关系与遗传多样性对后续的选育工作至关重要。
简单重复序列区间(inter-simple sequence repeat,简称ISSR)分子标记,由简单重复序列(SSR)标记技术发展而来,ISSR分子标记结合了随机扩增多态性DNA(randomly amplified polymorphic DNA,简称RAPD)的优点,同时克服了RAPD、SSR、限制性片段长度多态性(restriction fragment length polymorphism,简称RFLP)等技术的缺点[6-7],具有高效、快速、稳定、易操作、成本低等优点,目前广泛应用于亲缘关系鉴定、种质资源遗传多样性分析等领域。本试验采用ISSR分子标记技术对湖南本土牡丹亲缘关系和遗传多样性进行研究,为选育优良牡丹品种提供理论基础。
1 材料与方法
1.1 试验材料
通过走访湖南当地林业和农业部门、公司、企业、个体从业爱好者,对栽培时间长、来源不清的湖南牡丹种质资源进行调查。2014—2016年于开花期(3月下旬至4月上旬)、种子成熟期(7月下旬)、引种期(10月中下旬)在湖南牡丹的主要产地(邵阳、湘西、长沙)对资源识别、挂牌标记、形态特征、观赏特性及生长情况进行调查,共收集了18份性状稳定、植株平均年龄10年以上的湖南本土牡丹品种(表1)。采集牡丹新鲜嫩叶保存于-80 ℃冰箱。
1.2 DNA提取与检测
采用笔者所在实验室改良的十六烷基三甲基溴化铵(hexadecyl trimethyl ammonium bromide,简称CTAB)法进行提取,提取的样品用紫外分光光度计检测质量浓度,用1%琼脂糖凝胶电泳粗略检测样品DNA的浓度与纯度,最后稀释标定为50 ng/μL,保存到-20 ℃冰箱备用。
1.3 ISSR-PCR优化反应体系和引物筛选
1.3.1 ISSR-PCR基本反应体系 通过多次重复验证试验,筛选出2×TransTaqHigh Fidelity PCR SuperMixⅠ(内含适量的Mg2+、dNTPs、TaqDNA聚合酶)作为总的反应环境。25 μL PCR反应体系:2×TransTaqHigh Fidelity PCR Super MixⅠ 12.5 μL、引物(10 μmol/L)1 μmol/L、模板(50 ng/μL)50 ng,最后加入ddH2O补足25 μL。

表1 18份牡丹品种及来源
1.3.2 ISSR-PCR优化反应体系 按照L9(33)正交试验设计的方法[8-10],进行体系总体积、模板浓度、琼脂糖浓度的正交组合,每个处理重复3次(表2),最终建立的最佳反应体系:20 μL反应总体积,模板浓度30 ng/μL,琼脂糖浓度1.5%。然后在BIO-RAD PowerPac HV下以110 V的恒定电压电泳30 min,最后用凝胶成像系统(BIO-RAD Gel DocTMXR+)拍照分析。
扩增反应在Applied Biosystems 9902 PCR仪上进行,PCR反应程序:95 ℃ 4 min;94 ℃ 45 s,50~60 ℃ 45 s,72 ℃ 1.50 min,共计35个循环;72 ℃延伸5 min,4 ℃保存[11]。
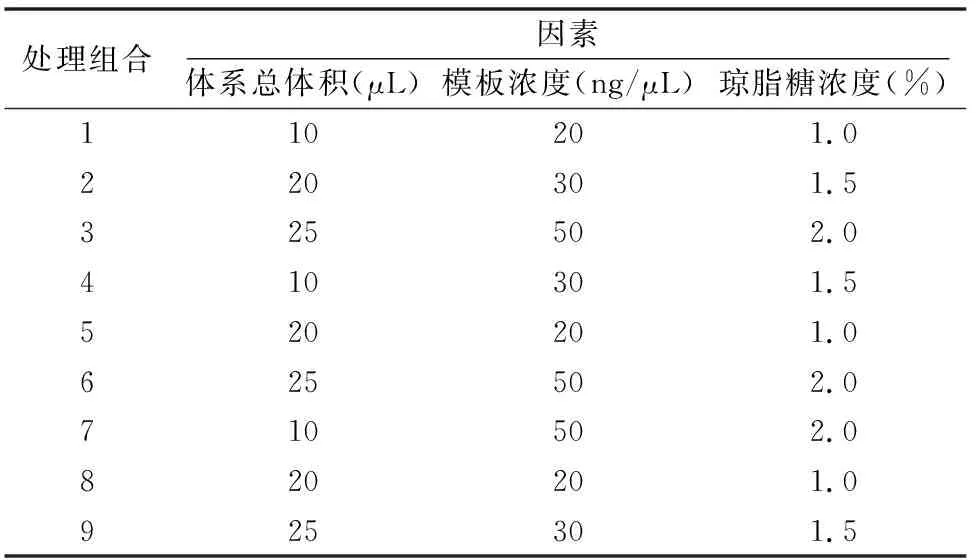
表2 ISSR-PCR反应体系的正交试验
1.3.3 引物筛选 选用哥伦比亚大学(UBC)开发的ISSR 100个通用引物[12-13],利用优化的反应体系进行引物筛选,引物由英潍捷基(上海)贸易有限公司合成;引物退火温度参照相关研究[4,11-12]。最后从100条引物中筛选出11条条带清晰、重复性好的引物。
1.4 图谱分析
图谱以0、1数据统计,在同一引物、同一位点根据条带的有(1)、无(0)形成原始数据矩阵(0、1)矩阵[14-17]。利用PopGen32软件计算多态位点比例(PPB)、Shannon’s信息指数、Nei’s基因多样性指数等。用NTSYS2.10软件计算遗传相似系数和构建品种亲缘关系。
2 结果与分析
2.1 牡丹DNA提取结果
牡丹基因组DNA经1%琼脂糖凝胶电泳,每个点样孔包括5 μL样品DNA和1 μL 上样缓冲液,各个条带大小基本一致,基本无拖尾现象(图1)。
2.2 ISSR-PCR体系筛选结果
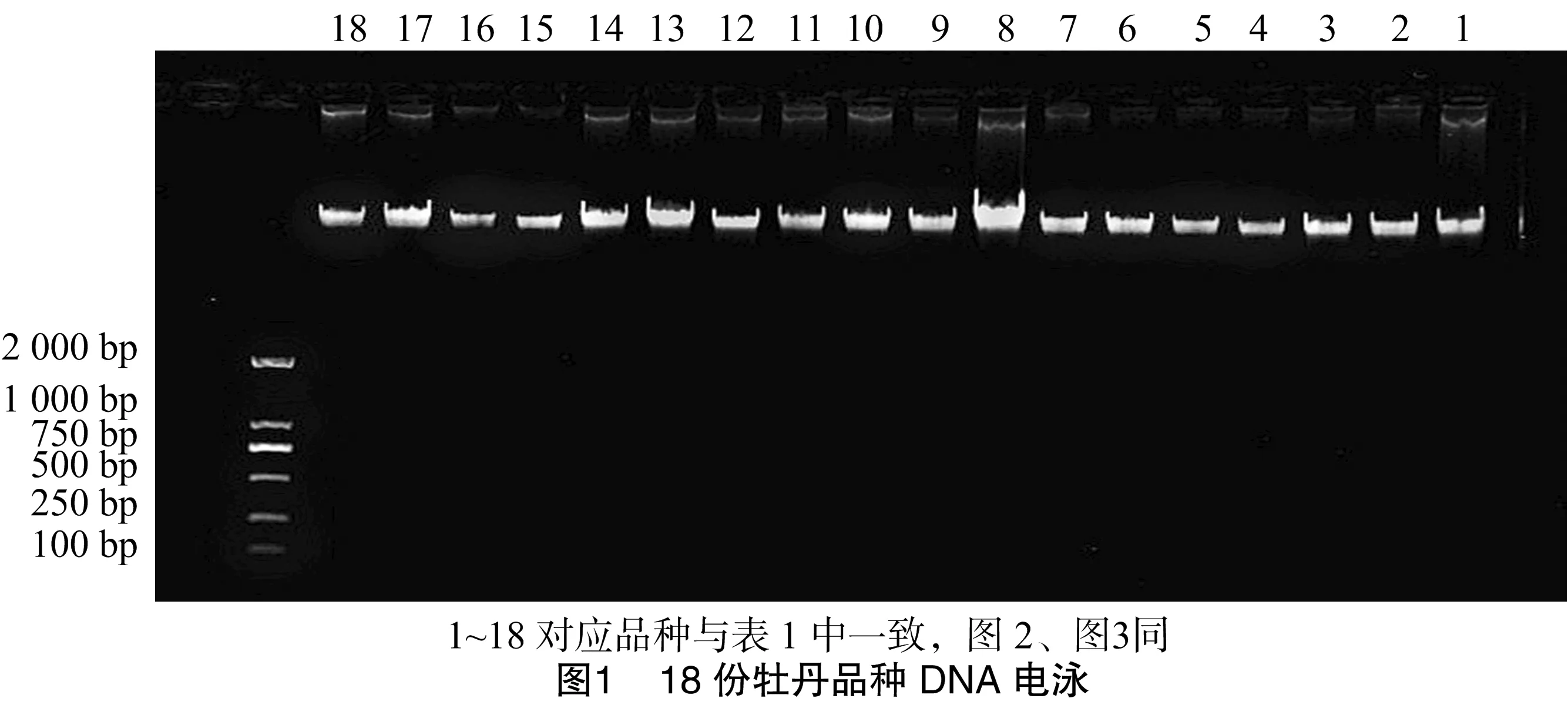
反应体系(20 μL)如下:2×TransTaqHigh Fidelity PCR SuperMixⅠ 10 μL、DNA模板(30 ng/μL)1 μL、引物1 μL、ddH2O 8 μL。用1.5%琼脂糖凝胶对扩增结果进行电泳。由图2可以看出,条带数量多且清晰可见,易于区分。
2.3 引物扩增结果
11条引物共扩增出162条条带,多态性条带133条,平均多态性比例82.6%。其中,引物UBC811扩增的条带数最多,为22条,多态性比例86.4%(图3、表3)。
2.4 18份牡丹品种遗传多样性分析
本试验通过软件PopGen32分析出供试牡丹品种的有效等位基因数(Ne)介于1.05~1.90之间,平均值为1.34[12-17];Shannon’s信息指数(I)介于0.12~0.60之间,平均值为 0.34;Nei’s(1973)基因多样性指数(H)介于0.05~0.40之间,平均值为0.22。
2.5 18份牡丹品种非加权组平均法(unweighted pair-group method with arithmetic means,简称UPGMA)聚类分析
用NTSYSpc2.1软件构建全部材料UPGMA树状图(图4),以相似系数0.733为阈值可将18份牡丹品种分为三大类。凤丹白、凤丹星、凤丹紫、凤丹、凤丹粉被划为第一大类,共5个品种,其中凤丹白与凤丹星的相似系数最近,说明它们的亲缘关系最近。第二大类包括凤丹绿、凤丹玉、凤丹绫、凤丹舞、宝庆红、宝庆粉共6个品种。第三大类包括杨山牡丹、永顺粉、宁乡红、郦家香、紫绣球、湘绣球、湘紫斑。当相似系数值为0.74时,把18份牡丹品种分为4个类群,其中C8和C9单独分为一类,由此说明凤丹绿与凤丹玉2个品种的遗传背景与其他品种差异比较大。
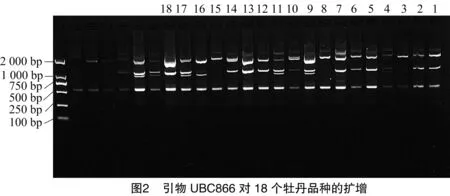
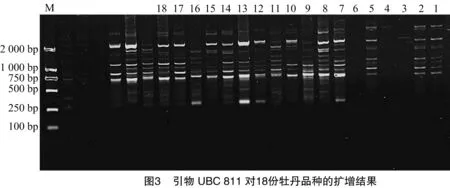
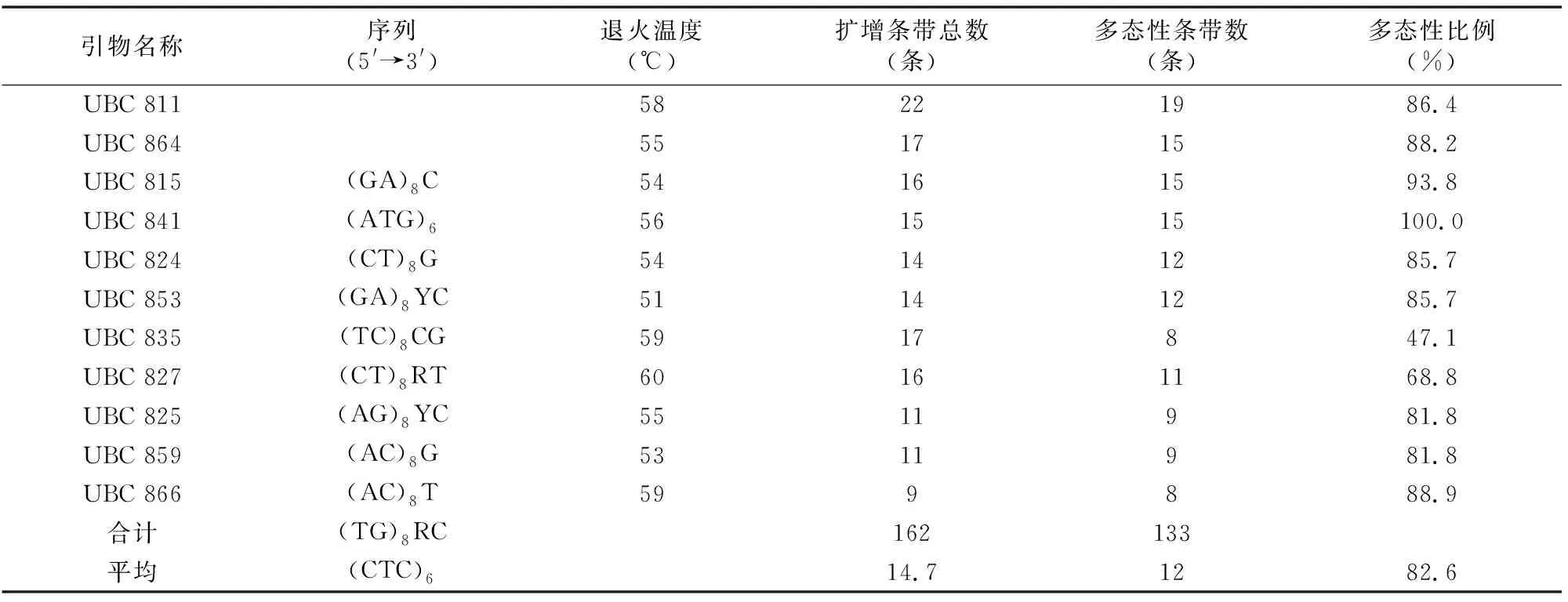
表3 ISSR引物及其扩增产物的多态性水平
3 讨论
本研究通过反复试验挑选出混合了Mg2+、dNTPs、TaqDNA聚合酶在内的反应体系, 然后从总反应体积、 DNA模板浓度、琼脂糖浓度3个方面进行结果筛选对比,最后挑选出了适合大部分引物扩增的最适反应体系,而且结果稳定,有效地节省了试验时间,提高了试验的精确度。

本试验用筛选出的11条引物对18份湖南本土牡丹资源进行扩增,共扩增出164条条带,多态性比例为81.1%,由此说明湖南本土牡丹资源丰富,品种间存在较丰富的遗传变异,具有比较复杂的遗传背景。
本研究中以NTSYS 2.1作为分析软件,利用UPGMA法在阈值为0.74时将供试牡丹品种分为4个类群[18-20],分析结果可以看出,基本品种都显示出花色和花瓣复杂程度的优势优先聚合在一起。第一类牡丹品种花色基本都是白色,只有凤丹粉为粉色,凤丹紫为紫色,初步分析这2个品种可能是凤丹白与非白色系牡丹品种的杂交后代表现出的亲本性状。第二类2个品种花型偏小,植株比较低矮,所以从聚状图上可看出这2个品种单独聚为一类,说明形态学鉴定与分子标记技术具有一定的关联性,表明这2个品种的遗传性状相比其他品种差异比较大。第三类中凤丹绫与凤丹舞都为白色花,凤丹绫的花瓣顶部羽状浅裂,凤丹舞的花瓣顶部锯齿状浅裂,花型都为单瓣型,相似系数比较大,说明亲缘关系比较近。宝庆红与宝庆粉2个品种在形态学性状上基本相似,且在分子标记学上表明它们的亲缘关系很近。第四类几乎所有的品种花色都比较艳丽,宁乡红与郦家香的相似系数最接近,说明它们的遗传背景相似;紫绣球与湘绣球花型都为球花台阁型,花色比较接近,根据聚状图的分析,它们的亲缘关系比较接近,而湘紫斑的相似系数最小,说明它们的亲缘关系最远。从来源地分析,表明同一地域品种相似度比较高,而且基本是2个形状非常相似的品种聚合在一起,推测是由于近距离的异花授粉而出现的性状分离的杂交后代。
目前,牡丹新品种选育基本依赖于传统的人工杂交方法,耗时长,基本每3年才能进行选育;选择亲本的过程比较盲目,而湖南有丰富的牡丹种质资源,牡丹品种具有耐高温、高湿、抗病虫害的优良性状,充分了解湖南本土牡丹品种的遗传多样性与亲缘关系将有助于提高选育的效率与质量,从而为观赏牡丹与湖南本土油用牡丹的杂交提供便利,为牡丹在南方市场的推广打下基础。
参考文献:
[1]洪德元,潘开玉. 牡丹芍药分会:推动牡丹深加工[J]. 中国花卉园艺,2015(5):27.
[2]洪 涛,张家勋,李嘉珏,等. 中国野生牡丹研究(一)芍药属牡丹组新分类群[J]. 植物研究,1992,12(3):223-234.
[3]杨淑达,施苏华,龚 洵,等. 滇牡丹遗传多样性的ISSR分析[J]. 生物多样性,2005,13(2):105-111.
[4]石颜通,周 波,张秀新,等. 牡丹89个不同种源品种遗传多样性和亲缘关系分析[J]. 园艺学报,2012,39(12):2499-2506.
[5]周 波,江海东,张秀新,等. 部分引进牡丹品种的形态多样性[J]. 生物多样性,2011,19(5):543-550.
[6]陈涛林,王汉超,罗军武,等. 汝城白毛茶种质资源遗传多样性和亲缘关系的ISSR分析[J]. 分子植物育种,2017,15(9):3780-3787.
[7]刘 彤,葛智文,陈涛林,等. 柳州九万山野生茶树种质资源遗传多样性的ISSR分析[J]. 南方农业学报,2015,46(11):1943-1948.
[8]麻 扬,白学良,赵东平. 山羽藓ISSR-PCR反应体系优化及引物筛选[J/OL]. 分子植物育种,2017-09-27[2017-11-10]. http://kns.cnki.net/kcms/detail/46.1068.S.20170927.1711.016.html.
[9]任秀霞,张 盈,薛璟祺,等. 滇牡丹天然居群的遗传多样性分析[J]. 植物遗传资源学报,2015,16(4):772-780.
[10]赵孟良,孙雪梅,王丽慧,等. 43份菊芋种质资源遗传多样性的ISSR分析[J]. 西北农林科技大学学报(自然科学版),2015,43(9):150-156,177.
[11]张旻桓,金晓玲,成仿云,等. 湖南产牡丹遗传多样性的ISSR分析[J]. 中草药,2016,47(7):1193-1198.
[12]杨美玲,唐 红. 紫斑牡丹遗传多样性的ISSR分析[J]. 西北植物学报,2012,32 (4):693-697.
[13]高昊丹. 芍药属植物的综合评价及其亲缘关系的研究[D]. 沈阳:沈阳农业大学,2012.
[14]童 芬,谢登峰,曾心美,等. 四川牡丹和圆裂四川牡丹遗传多样性的ISSR分析[J]. 西北植物学报,2016,36(10):1968-1976.
[15]杨美玲. 紫斑牡丹遗传多样性的ISSR分析[D]. 兰州:甘肃农业大学,2012.
[16]蔡 健,李 浩,赵 翔,等. 22个牡丹品种形态性状的多样性分析[J]. 安徽农业科学,2015,43(35):57-59.
[17]孙嘉磊,杨志刚,罗 兵,等. 21个日本牡丹品种DNA指纹图谱构建与EST-SSR标记遗传多样性分析[J]. 江苏农业科学,2015,43(9):50-53.
[18]宋倩倩,付茂旺,何艺凡,等. 福建省浙江红花油茶遗传多样性的ISSR分析[J]. 四川农业大学学报,2017,35(3):370-374.
[19]李 昂,葛 颂. 植物保护遗传学研究进展[J]. 生物多样性,2002,10(1):61-71.
[20]武玉珍,冯睿芝,张 峰. 基于ISSR分子标记的褐马鸡亲缘关系分析[J]. 生态学报,2015,35(4):1059-1067.
猜你喜欢
作物学报(2022年2期)2022-11-06
今日农业(2022年13期)2022-09-15
中国现代中药(2021年9期)2021-11-16
学生天地(2020年2期)2020-08-25
天然产物研究与开发(2018年10期)2018-11-06
中国麻业科学(2018年6期)2018-04-09
现代园艺(2017年13期)2018-01-19
广东农业科学(2017年5期)2017-08-29
北方音乐(2016年12期)2016-08-23
西南农业学报(2016年5期)2016-05-17
