
三叠系—侏罗系环境变化及界线研究方法综述
2018-06-07 06:57谭丽娟葛毓柱郭优优
地球科学与环境学报 2018年3期
谭丽娟,师 萌,葛毓柱,郭优优
(中国石油大学(华东) 地球科学与技术学院,山东 青岛 266580)
0 引 言
在晚三叠世至早侏罗世(即距今大约2 Ma以前),中部大西洋岩浆省的系列密集火山爆发先喷出了一部分SO2和其他火山挥发物,引起海水的初步酸化并形成了短暂的干冷气候;随后,火山活动释放的巨量CO2导致长期且强烈的温室效应,加剧海水酸化,同时带动洋底甲烷水合物的失稳,从而释放出大量甲烷,之后氧化成CO2排放到空气中,进一步促进温度的提高及海水酸化。前人研究表明,该地质转折时期温度上升了3 ℃~4 ℃[1-4]。海平面变化表现为一个迅速的海退后接着上升速率至少为每百年0.5 cm的海进旋回[5]。温度变化及水体环境改变导致陆相植被的更替及大规模的海相生物灭绝事件[4]。除此以外,大量火山喷发的CO2及其他气体也扰乱了全球碳循环,造成了碳同位素值的负偏移现象[2]。
目前,国际上已经综合利用古生物学、有机碳同位素分析及其他化学元素分析方法界定三叠系—侏罗系界线位置,恢复古地理环境及古气候变化。在高纬度的北美洲、欧洲地区海相剖面分布点,该研究已经十分详细,但在低纬度地区以及亚洲地区该研究仍然十分匮乏。迄今为止,中国仅见刘兆生等针对西藏地区、准噶尔地区及四川地区展开了相关研究[6-11]。全球范围内需要更多的研究区域和剖面来完善关于三叠系—侏罗系事件研究,且中国方面的数据有不可或缺的作用,因此,本文通过总结国际上研究该事件的相关进展及常用方法,以期为国内学者研究三叠系—侏罗系地层学提供参考,并为全球三叠系—侏罗系界线的研究提供相关证据。
1 三叠纪—侏罗纪之交研究现状
目前,三叠系—侏罗系界线的研究主要集中在古环境变化上,包括气候变化、海平面变化、生物种群变化等。
1.1 构造活动
三叠纪—侏罗纪全球性最重要的构造事件有:古特提斯洋闭合,泛大陆亚洲部分汇聚;泛大陆开始在未来中部大西洋区域分裂,并由裂谷活动向漂移运动转变;中部大西洋岩浆省(CAMP)发生了广泛的火山活动[12-13](图1)。总的来看,全球形成了特提斯和大西洋两大构造域。大西洋构造域以拉伸性构造活动为主,而特提斯构造域以挤压性构造活动为主。

图件引自文献[14]图1 全球三叠系—侏罗系界线及海相剖面大致古地理位置分布Fig.1 Approximate Paleolocation Distributions of Global Triassic-Jurassic Boundary and Marine Profiles
在特提斯构造域,古特提斯洋关闭和泛大陆亚洲部分的汇聚形成了泛大洋火山链,其发育于早二叠世,在其后各个时期具有活跃表现。在大西洋构造域,伴随着泛大陆的分裂活动,中部大西洋岩浆省出现并在三叠纪—侏罗纪火山活动强烈,被认为最有可能导致三叠纪—侏罗纪之交生物灭绝事件的发生[4]。中部大西洋岩浆省以拉斑玄武岩为主,面积大于7×106km2,分布于4个大陆之上。由于后期风化侵蚀效应,现存面积小于1×106km3,以岩墙、岩盖为主[15]。
1.2 大气和气候变化


图件引自文献[1]、[17]、[20]~[23]图2 三叠纪—侏罗纪之交全球环境、生物种群及有机碳变化Fig.2 Variations of Global Environmental, Biological Species and Organic Carbon During Triassic-Jurassic
晚三叠世另外一个显著的气候特征表现为巨型季风性,这与晚三叠世古地理格局有关。晚三叠世地球上所有大陆聚集在一起,形成几乎以赤道对称分布的泛大陆,周围被泛大洋所包围,同时特提斯洋在泛大陆中部形成了狭长的海道。巨型季风性气候产生以下效应:丰沛极端的季节性降水主要集中在夏季和北半球;泛大陆赤道区气候相对干旱,高纬度地区和特提斯滨海域气候相对湿润;全球性气候分带不明显[24]。
1.3 海平面变化
Hallam曾根据三叠系—侏罗系界线下伏地层顶部存在沉积间断及其与上覆地层岩性的显著不同,提出三叠纪—侏罗纪之交在包括欧洲、南美洲、北极、加拿大等地区,海平面变化表现为一个海退-海进旋回,并计算出海平面上升速率至少为每百年0.5 cm[25],之后很多学者通过不同剖面研究也证实了这个观点。但在中国西藏及其附近地区未发现明显相变,未见海平面下降的证据,Greene等认为海平面的迅速下降可能只是局部现象,主要发生在临近中部大西洋岩浆省火山活动的地区[12]。关于引发海平面变化的因素目前众说纷纭:多数学者提出三叠纪—侏罗纪海平面先短期下降后持续上升,并与中部大西洋岩浆省火山活动有关;而Hesselbo等提出三叠纪—侏罗纪海退-海进旋回是由全球海平面升降引发的,是一个长期的变化,在界线处实际不存在短期并且剧烈的海平面变化[1];Honig等认为古赤道区的阿拉伯海域海平面升降是由全球海平面变化及局部构造活动造成的[26]。海平面变化受多种因素影响,构造、气候、天文周期均可影响海平面变化,关于三叠纪—侏罗纪之交海平面变化(尤其是局部海平面变化)的控制因素研究可能对古环境探讨更具有揭示意义(图2)。
1.4 生物种群灭绝事件及灭绝原因
1.4.1 灭绝事件
受中部大西洋岩浆省火山活动的影响,晚三叠世的生物种群发生大规模灭绝,海、陆生物种群均受到影响。其具体表现为陆地植物从木质松柏科植物逐渐演替成草本蕨类植物[27-28],同时伴随种子蕨类Dicroidium和Peltasperm群的重大损失[24,29];海洋生物受到的影响则更加严重,其中大约53%的属和23%的科遭受灭绝,严重程度超过了白垩纪末灭绝事件,其中菊石、双壳、珊瑚和介形虫遭受灭绝程度最为严重[29]。Tanner等发现海洋生物中牙形石、双壳类及菊石类生物均表现为渐进式灭绝,放射虫在科的水平上没有展示重大灭绝,但是在种的水平上出现重大转折[4]。海洋光合浮游植物总体由红藻类转变为绿藻类,其中钙质微型浮游生物和颗石藻灭绝,青绿藻和疑源类爆发[28,30-32]。
值得注意的是,此次生物灭绝事件具有选择性。礁类居住者比水底居住者灭绝率高;近滨灭绝率大于远滨灭绝率;喜碳酸盐基底者灭绝率高于喜硅质基底者;低纬度地区灭绝率高于高纬度地区;内生文石壳类灭绝率高于表生滤食钙质壳类[33-34]。另外,不同地区生物灭绝时期不同。例如,东、西特提斯洋菊石灭绝表现为穿时性特征[10],而古赤道温暖的海水可能保持了文石的过饱和状态,生物在中部大西洋岩浆省火山活动初期仍可以生存[26,35]。
此外,关于海、陆生物灭绝的时间先后问题也引发了探讨。尽管Palfy等根据海、陆三叠系—侏罗系剖面同期火山物质定年结果对比,认为陆地生物灭绝可能早于海洋生物灭绝至少0.7 Ma[36],但陆地三叠纪孢粉消失与海洋生物灭绝同时,表明海、陆生物灭绝事件同步[27,37]。同时,Schaltegger等利用火山灰中锆石测定三叠系—侏罗系界线的时代大约为201 Ma,再次印证了海洋生物灭绝时间与中部大西洋岩浆省火山活动时间相吻合[38](图2)。
1.4.2 灭绝原因
(1)温度降低。Guex等认为灭绝事件与火山SO2的喷发而导致气候变冷有关[18],这一观点受到孢粉研究的支持。但是气候变冷事件是否为全球性事件值得探讨,在中国西藏剖面生物特征就不支持这一观点[10]。
(2)海平面波动与海水缺氧。海平面变化会影响浅海生物栖息范围和浅海沉积基底、盐度等。这种情况在缺乏礁类建造的碳酸盐岩缓坡尤为显著,轻微的海平面变化都会导致沿海地区大面积的暴露和淹没。在三叠系—侏罗系界线,海平面变化在多地表现为一个海退-海进旋回。资料显示,海平面下降期出现了菊石和浅水双壳类灭绝,海平面上升期出现了放射虫变化和牙形虫的最后灭绝[18,39]。但单凭海平面波动很难造成大型生物灭绝事件,而且海平面波动也无法解释陆地生物灭绝事件[5,40]。在地质历史时期,许多明显的海平面波动并不伴随重要的生物变动。基于一些研究剖面中沉积有黑色页岩,海水缺氧事件被提出作为生物灭绝原因之一[18]。晚三叠世全球温室效应及海平面上升可能导致海水水体交换受阻,分层性加强,有利于出现海水缺氧现象。但是,三叠系—侏罗系界线海水缺氧现象缺乏全球性表现,一些剖面氮同位素和微量元素研究表明氧化状况得到改善[41]。另外,生物选择性灭绝也不完全支持海水缺氧作为生物灭绝原因。
(3)海水盐度降低及循环受阻。海洋是个动力系统,表面水体主要受风力驱动,深部水体运动与温盐差有关。晚三叠世升高的CO2浓度和加强的季节性气候促进陆地风化侵蚀作用,大量淡水和陆源物质向滨海区域输入,使沿海地区盐度降低,一部分古微生物(有孔虫及介形虫)灭绝[20]。由于气候变暖,海水分层性加强,水体循环受阻,整个海洋活动受滞。海水营养物质循环同样受阻,深部生物体分解所产生的营养元素无法向表层生物供应,尤其是元素N,即使有陆源输入也无法弥补,这无疑会对海洋初始生产率造成重大影响,从而固氮菌藻类爆发。Kasprak等指出三叠系—侏罗系界线在泛大洋东北处出现光合区停滞现象[42]。
(4)海水酸化及碳酸盐饱和度降低。火山活动或天然气水合物失稳会产生大量CO2并融入到海水之中,从而使海水pH值减小,产生海水酸化现象[43]。晚三叠世和早侏罗世,海水中主要沉积的碳酸盐矿物为文石,海水酸化现象会降低碳酸盐饱和度,对分泌文石的海相生物种群造成损害。实验表明,碳酸盐饱和度降低的情况下,原本分泌文石的藻类开始分泌低Mg方解石,这不利于文石生物的繁殖和竞争力[44]。同时,在高CO2浓度和海水酸化条件下,生物倾向于方解石沉淀,这也可以解释文石生物的选择性灭绝现象[40]。

图件引自文献[14]图3 理想条件下三叠系—侏罗系界线的沉积剖面类型Fig.3 Idealized Sedimentary Profile Types of Triassic-Jurassic Boundary
1.5 岩石地层情况
很多学者认为受大量CO2释放导致的海水酸化影响,三叠纪—侏罗纪之交沉积的碳酸盐岩剖面中存在着一个全球性的沉积间断(碳酸盐岩含量(质量分数,下同)不同幅度的下降),并与碳同位素值的负偏移相对应,这也从侧面印证三叠纪—侏罗纪海水酸化导致碳酸盐不饱和的结果,但是在碎屑岩沉积剖面中不会有受海水酸化影响的痕迹。现有的三叠系—侏罗系界线的岩石记录大致可以分为3种。
(1)存在沉积间断的碳酸盐岩剖面[图3(a)]。英属哥伦比亚威利斯顿湖剖面在三叠系—侏罗系界线下沉积不整合层,该层沉积物由富双壳类的碳酸盐岩过渡为含硅质碎屑的碳酸盐岩混杂层;在三叠纪—侏罗纪之交,沉积一段80 cm厚的碳酸盐岩含量仅为4%的夹层,这可能是该区由海水酸化引起的生物钙化危机的一种表现[39]。美国纽约纳维达剖面中,三叠纪末期的生物灭绝发生在加比组米勒峡谷段底,上覆岩层即为三叠系—侏罗系界线层,同样出现了碳酸盐岩大幅度降低的情况,含量大约为10%,而沉积在界线层附近的碳酸盐岩含量则超过80%[18]。Greene等认为碳酸盐岩含量的剧烈降低不是大量的硅质碎屑输入导致的,而是因为碳酸盐岩产能被抑制[14]。北阿尔卑斯山地区三叠系—侏罗系界线层为一套碳酸盐岩含量为9%~15%的泥灰岩,同时泥灰岩是一套典型的三叠系—侏罗系界线沉积物[29]。南阿尔卑斯山地区界线处近物源区一般沉积含Fe的硬灰岩凝缩层;远物源区附近沉积1 m厚的粉砂岩-泥灰岩夹层[45]。在亚平宁山脉北部和中部地区,三叠系—侏罗系界线处也出现了碳酸盐岩产能的下降。亚平宁山脉北部瑞替阶晚期沉积物已经过渡为粉砂岩,界线处沉积夹层于灰岩中的暗色页岩;亚平宁山脉中部界线处沉积一段黑色页岩[46]。
(2)连续沉积的碳酸盐岩剖面[图3(b)]。南阿尔卑斯山地区的东欧在三叠系—侏罗系界线处可能连续沉积碳酸盐岩剖面,多为薄层的半深海生物灰岩或深水碳酸盐岩沉积[47-48]。而在古赤道附近的阿拉伯海穆桑达姆半岛地区,碳同位素值负偏移发生在生物灭绝事件之前,三叠系—侏罗系界线处的沉积物则只是由生物灰岩过渡到鲕粒灰岩,未见碳酸盐岩含量有明显下降[26]。
(3)碎屑岩主导的沉积剖面[图3(c)]。这类剖面大多在东亚地区,如日本西南地区就存在碳酸盐岩含量为0的深水沉积剖面,含放射虫的燧石层为该剖面的三叠系—侏罗系界线层[49]。位于中国西藏地区的格米格剖面也属于这一类型,沉积背景为浅水的硅质海环境,在界线层也未表现出明显的相变[50]。位于中国准噶尔盆地的郝家沟剖面,界线处表现为一套辫状三角洲沉积[51]。中国四川盆地剖面的界线岩性标志则较为复杂,上三叠统为分布广且稳定的灰色及黄灰色长石石英砂岩,夹粉砂质泥岩及粉砂岩,而下侏罗统则有局部差异。四川盆地北部或西北地区沉积石英砾岩或燧石砾岩,南部沉积石英砂岩,重庆万州剖面则沉积泥质粉砂岩或粉砂质泥岩。中国鄂西秭归盆地西部的界线层沉积石英砂岩夹砾岩[52]。
1.6 碳同位素值变化
现今阶段,已有研究报告显示关于三叠系—侏罗系界线的碳同位素值(δ13C)大致可以分为两类:第一类在界线附近,即在瑞替阶晚期及赫塘阶早期初始碳同位素值负偏移(Initial Negative CIE)出现,同时伴有后续的小段碳同位素值正偏移(Positive CIE)及一大段的主要碳同位素值负偏移(Main Negative CIE)[53-55];另一类则是缺少初始碳同位素值负偏移,连续的碳同位素值正偏移被一段主要碳同位素值负偏移打断[29,56](图4)。现阶段研究中,多数学者认为三叠系—侏罗系界线应该与主要碳同位素值负偏移相对应,虽然初期研究成果曾认为碳同位素值正偏移可能与取样精度有关[1,27]。至于部分剖面缺少初始碳同位素值负偏移的原因可以归纳为:①瑞替阶时,海平面下降导致部分碳酸盐岩台地出露,受到剥蚀风化作用[53];②在碳酸盐岩缓坡远端的底流区或该区沉积的风暴岩沉积了凝缩层,并发生铁质矿化导致初始碳同位素值负偏移缺失[45,54]。

LOTF(Last Occurrence of Triassic Fauna)表示三叠系生物最后一次出现的位置;FOJA(First Occurrence of Jurassic Ammonoids)表示侏罗系菊石首次出现的位置;FOJF(First Occurrence of Jurassic Fauna)表示侏罗系生物首次出现的位置;δ13Ccarb为无机碳同位素值;图件引自文献[45]和[54]图4 奥地利Loruns和 Steinernes Meer剖面三叠纪—侏罗纪之交碳同位素值偏移及生物灭绝事件Fig.4 Carbon Isotope Excursions and Relevant Biological Extinction Events of Loruns and Steinernes Meer Profiles from Austria During Triassic-Jurassic
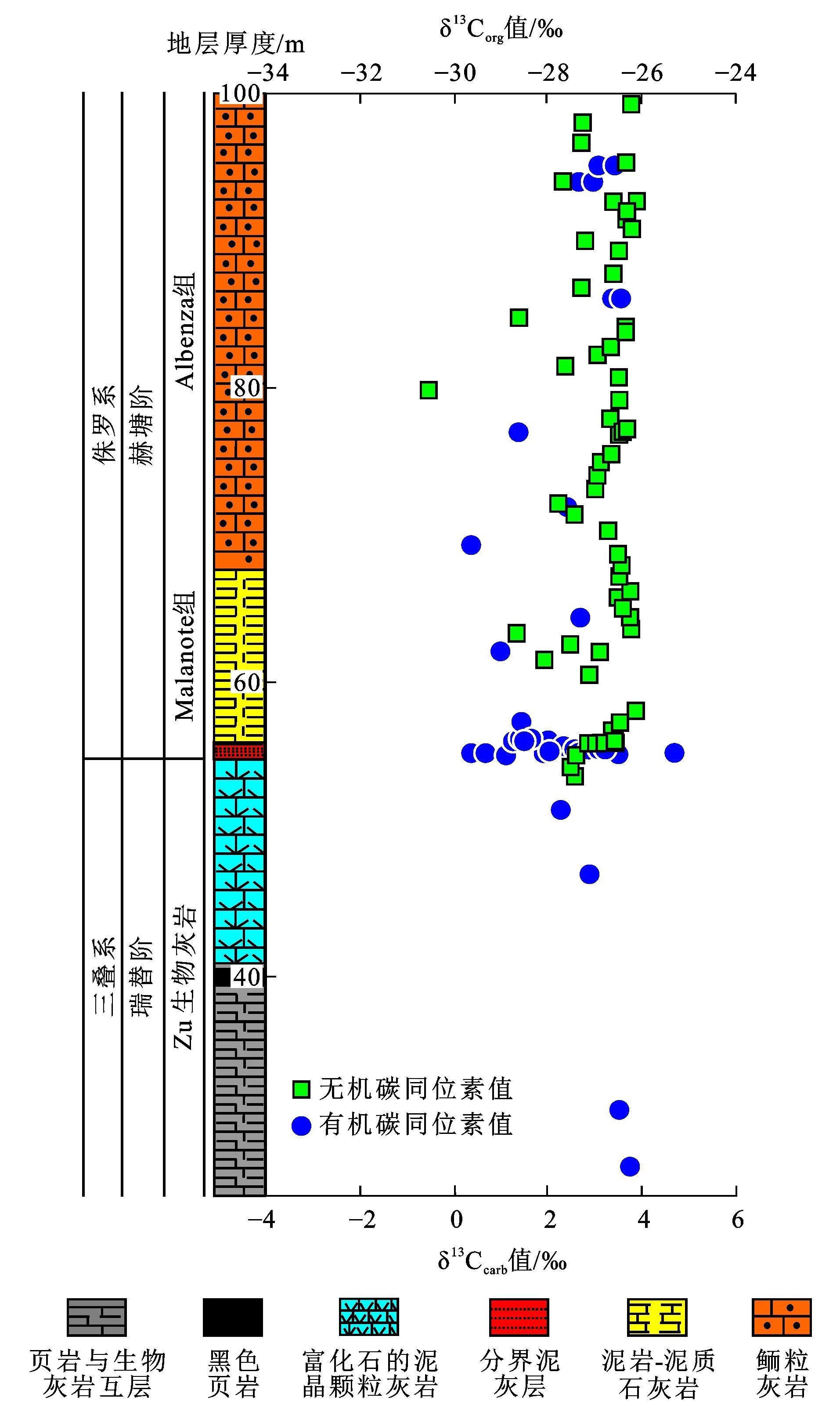
δ13Corg为有机碳同位素值;图件引自文献[57]图5 意大利伦巴第盆地三叠系—侏罗系界线的碳同位素值随深度变化Fig.5 Variation of Carbon Isotope Values with Depth in Triassic-Jurassic Boundary of Lombardy Basin, Italy
三叠系—侏罗系界线的碳同位素值负偏移出现在全球剖面的不同环境中,虽然偏移幅度随地区、沉积物组成和环境发生变化,但在有机碳和无机碳中均有体现,表现为全球性的碳库扰动事件(图5)。同时,McRoberts等研究发现,初始碳同位素值负偏移位置通常与生物灭绝事件相对应,而随后短暂的碳同位素值正偏移及主要碳同位素值负偏移则代表了后灭绝时期生物的恢复阶段[20]。但是,Lindstrom等发现示踪生物化石及双壳类的灭绝发生在初始碳同位素值负偏移之前[58]。
碳同位素值偏移可能与环境变化、成岩作用、有机质组成变化等多种因素有关。例如,海相碳酸盐岩可能会通过与周围液体发生成岩作用(如重结晶作用或新生变形作用)导致碳同位素值发生变化;而有机质的碳同位素值变化则可能由于热解作用产生的烃类替换了原有的碳同位素[59]。关于三叠系—侏罗系界线碳同位素值偏移的成因大致可以分为:①中部大西洋岩浆省火山活动通过喷发大量CO2,岩浆热变质作用及海底甲烷水合物失稳提供大量富含12C的轻质碳至大气-海洋系统中,造成碳同位素值负偏移;②随后出现的无机碳及有机碳同位素值正偏移则可能是因为全球范围内有机碳埋藏速率增加,在来自中部大西洋岩浆省的主要火山活动中,大量额外的CO2再次喷入大气中引起温度的升高及广泛的海洋缺氧环境,造成富含12C的海洋沉积物中有机碳埋藏速率增加,从而提高了大气-海洋系统中的碳同位素值[57,60-64]。只在部分剖面出现的主要碳同位素值负偏移则可能对应了又一次的生物灭绝事件,这说明生物圈通过有机质组成变化也可导致碳同位素值的变化[18]。
2 三叠系—侏罗系界线研究方法
2.1 古生物化石标定法
2.1.1 菊石生物地层学
在三叠纪—侏罗纪海洋生物记录中,菊石通常被认为是可以用来划分地层的最可靠化石。在全球很多剖面研究中,都可以见到以菊石带为基础划分的生物地层,一般包括最后一次出现的晚三叠世菊石Choristocerasmarchi及随后首次出现的早侏罗世菊石Psilocerasspelae。各剖面典型菊石分布见表1。
2.1.2 孢粉生物地层学
孢粉生物地层学的应用非常广泛,可以同时反映海洋生物系统及陆地生物系统的相应变化。通过分析散落在地层中的孢粉成分及统计其体积分数,结合其他地球化学数据(如高精度碳同位素记录)可以帮助划分地层;通过将各孢粉按已知的或可能的母体植物亲缘关系分属种,结合已有记录的母体植物适合生存环境,可以推测古气候、古植被及其他环境变化情况[65]。
Lindstrom等研究发现三叠纪—侏罗纪海洋浮游生物遭受了强烈损失,尤其在沟鞭藻囊孢中的Suessiaceae科,Rhaetogonyaulaxrhaetica就是其中之一,它的消失与三叠纪末期灭绝事件同步,也可能刚好发生在其后不久的时间段内。因此,它的最后普遍出现时间(Last Common)、最后连续出现时间(Last Consistent)以及末现(Last Occurrence)可以被用来作为地层划分标志[23]。其他孢粉类群(包括Granuloperculatipollisrudis、Rhaetipollisgermanicus、Ovalipollisovalis、Limbosporiteslund-bladiae、Semiretisporisgothae、Polypodiisporitespolymicroforatus和Ricciisporitestuberculatus)的末现也可用于地层划分。同时,研究表明,许多孢粉类群的首现(First Occurrence)、首次连续出现(First Consis-tent)及首次普遍出现(First Common)与其他方法所定的三叠系—侏罗系界线时间极为接近。例如,可以用来标志侏罗纪早期的很可能为松柏植物花粉Cerebropollenitesthiergartii的首现及松柏科花粉Pinuspollenitesminimus的首次普遍出现、蕨类孢子Ischyosporitesvariegatus的首现、石松类孢子Kraeuselisporitesreissingeri的首现/连续出现/普遍出现,其均可用来界定三叠系—侏罗系界线[23]。中国陆相地层三叠系—侏罗系界线划分也常用孢粉分析法,早侏罗世蕨类植物孢子数量爆发(Densoisporitesscanicus、Densosporitescrassus、Cyathidites),于界线底部形成高峰值,十分显著,可用于划分地层[66]。
12)注意加强各专业的协调配合。在工程施工中,各专业交叉配合与协调是非常重要的,如果处理的不好,就会出现返工,造成工程投资的极大浪费,影响工期,有的还会影响到建筑物的使用功能,严重的甚至还会带来质量问题和安全隐患。可见工程施工中各专业的协调管理工作,不仅很重要,同时也很必要。作为建设单位,在工程的设计阶段以及施工过程中,应随时注意保持各专业的相互协调配合。
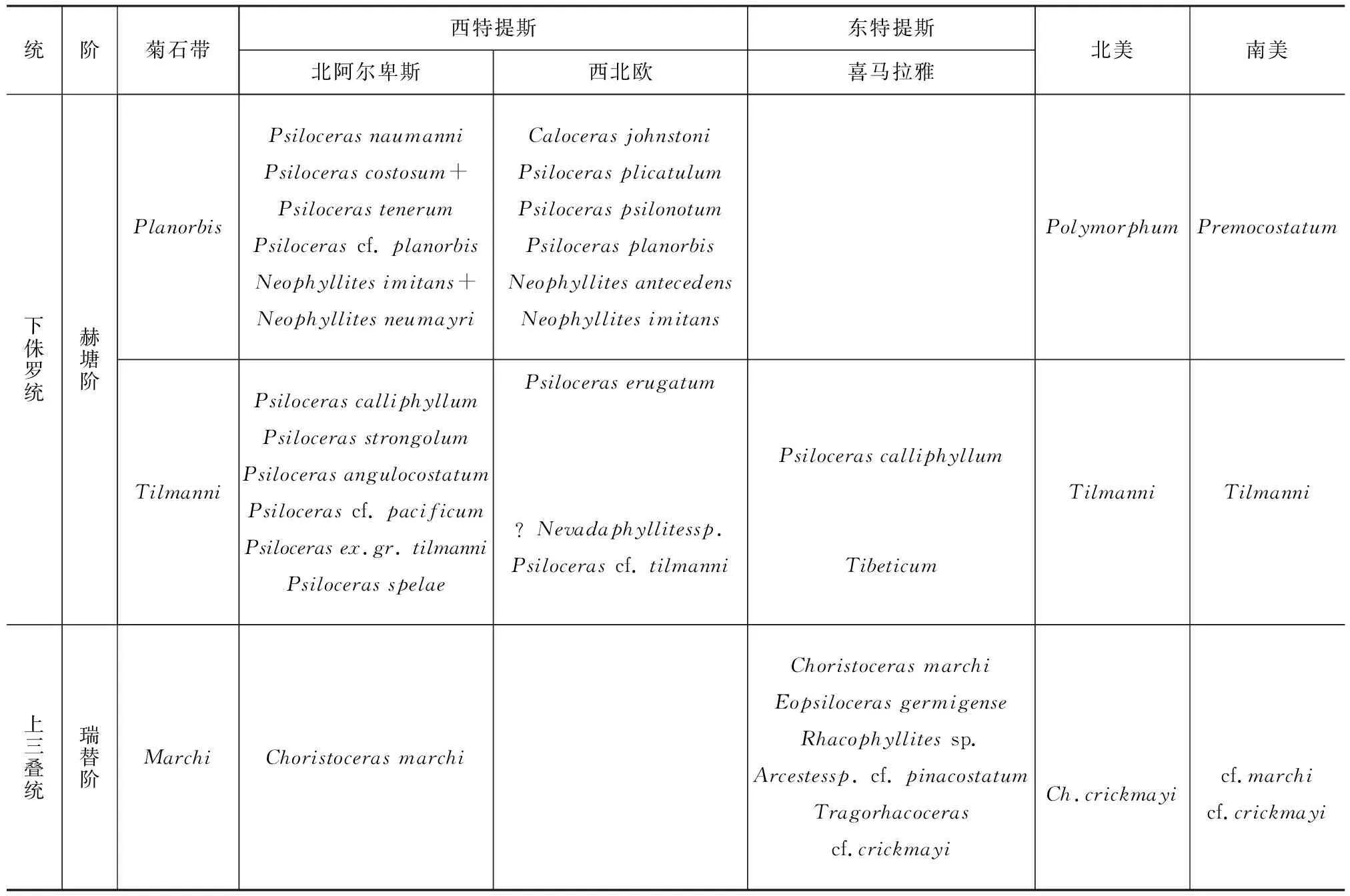
表1 不同地区上三叠统—下侏罗统菊石带比较Tab.1 Comparison of Upper Triassic-Lower Jurassic Ammonite Zones from Different Areas
注:资料引自文献[10]和[23]。
2.2 碳同位素标定法
三叠纪—侏罗纪中部大西洋岩浆省频繁密集的火山爆发喷出大量CO2及甲烷水合物,引发了系列环境变化及生物灭绝事件,这些通常都伴随全球碳循环扰动,在有机碳和无机碳中均有体现。稳定碳同位素法是近年来常用的地球化学手段,主要用于研究大气圈、生物圈、全球气候、陆地风化作用及海洋循环的变化,一般以国际标准物质PDB为参考。
当样品的13C/12C值比国际标准物质PDB高时,δ13C为正值,而δ13C为负值则意味着样品的13C/12C值比国际标准物质PDB低很多。碳酸盐岩中的主要沉积矿物方解石受空气中CO2影响较大,其含有更多氧化碳,无机碳中的δ13C值会偏高;而有机碳受光合作用中的分馏作用而含有更多的还原碳,其δ13C值偏低。因此,无机碳同位素值通常大于有机碳同位素值。然而,碳酸盐岩中的无机碳与有机碳相比更易与周围流体发生成岩作用,导致同位素组分发生变化,因此,有机碳同位素值比无机碳同位素值更稳定[62]。值得注意的是,白垩纪之前,陆相有机质比海相有机质更富13C,因此,大量的陆源输入也会提高海洋沉积物中的有机碳同位素值。
2.3 元素地球化学特征分析法
除了碳同位素标定法以外,近年来很多学者也采用元素地球化学特征分析法来研究三叠系—侏罗系界线及其环境变化。元素含量分布及其相关关系在不同环境下显示不同特征,岩石中各元素含量还受气候变化、物源输入、水体深度及水体介质的化学性质(如盐度、pH值)等因素的影响。吴智平等用不同元素的分布变化反映济源盆地三叠系—侏罗系界线地层沉积相的演变[67]。
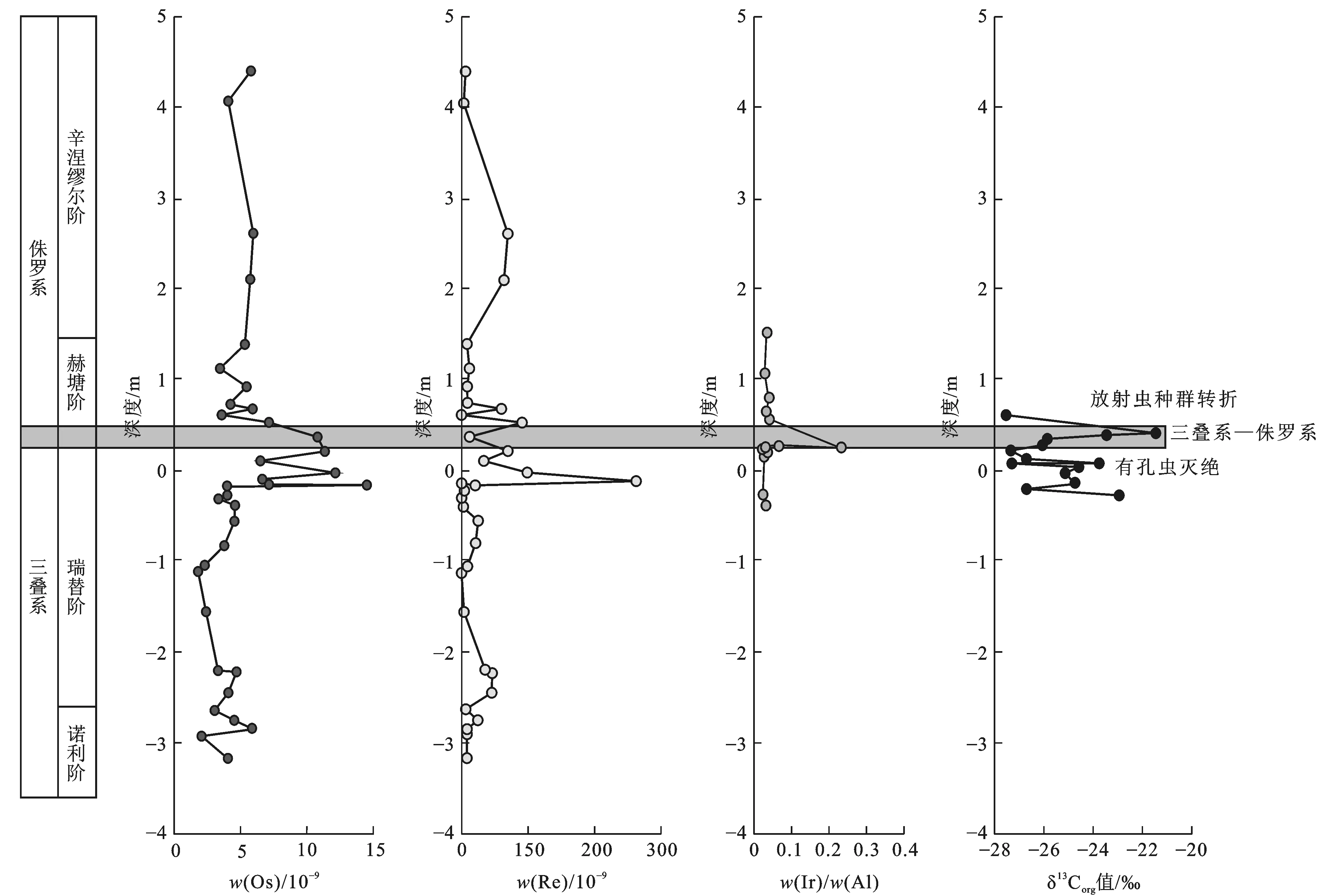
w(·)为元素含量;图件引自文献[49]和[68]图6 日本犬山地区三叠系—侏罗系界线的铂族金属元素异常随深度变化Fig.6 Variation of Platinum Group Elements Anomalies with Depth in Triassic-Jurassic Boundary of Inuyama Area, Japan
三叠纪—侏罗纪中部大西洋岩浆省火山活动的频繁发生会极大地增强陆地风化作用,从而提高海洋沉积物中陆源物质的输入,同时,大量的岩浆喷出侵入也会造成某些特定元素的分布异常,这些都可以反映在沉积物的化学元素特征中。铂族金属元素(Platinum Group Elements)就是可以用来反映该类事件影响的地球化学标志物。Kuroda等通过研究太平洋远洋沉积物,发现三叠系—侏罗系界线附近出现Os和Re分布的异常正偏移,并与该地区有孔虫的灭绝事件相呼应[68];Hori等在研究日本犬山地区盘古大洋深海沉积物层状燧石时也发现在三叠系—侏罗系界线附近出现铂族金属元素的异常高值,并同步于海洋浮游植物的灭绝事件[49](图6);而Tanner等在加拿大芬地盆地三叠系—侏罗系界线的美登组(Blomidon)地层也检测出Ir的异常高值[69]。上述现象表明,铂族金属元素在三叠系—侏罗系界线附近出现异常高值具有全球性特征,可用来帮助厘定三叠系—侏罗系界线的位置。
2.4 综合数据建立地质模型
现阶段的三叠系—侏罗系界线研究方法中,已经不再只采用单一方法,大多综合多种方法,常见的有综合生物化石证据及地球化学指标推测该时期的古气候和古环境。近年来,一些国外学者也利用数学建模的方法对三叠系—侏罗系界线研究进行半定量—定量分析。Kump等对有机碳同位素曲线及无机碳同位素曲线进行量化分析,并提出公式[70]
(1)
式中:Mo表示大气-海洋系统中的无机碳含量;Fw表示来源于风化作用的碳输入;Fvolc表示来源于火山活动的碳输入;Fb,org表示碳输出中沉积物有机碳的埋藏量;Fb,carb表示碳输出中沉积物无机碳的埋藏量;t为时间。
通过改变式(1)中各参数的取值范围,发现增长的有机碳埋藏量会导致大气中CO2浓度降低,与此同时无机碳及有机碳同位素值出现正偏移,虽然有机碳同位素值正偏移出现时间略晚于无机碳同位素值,但是其幅度更大。针对无机碳和有机碳同位素值正偏移幅度不同的情况,Bachan等认为这与有机碳中的可再循环组分有关,并进一步提出公式[57]
(2)

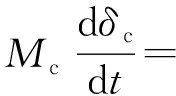
Fw,carb(δw,carb-δc)-Fb,org(Δorg)+
Fextra(δextra-δc)
(3)
式中:Mc代表溶解的无机碳物质的量;δc代表无机碳埋藏量中碳同位素值,因为埋藏无机碳无法进入下一步碳循环,所以计算其他来源的碳同位素值时需要将埋藏量的碳同位素值去除;δvolc为火山活动中的碳同位素值;Fw,org代表岩石风化作用导致的有机碳输入,δw,org为其相应的碳同位素值;Fw,carb代表岩石风化作用导致的无机碳输入,δw,carb为其相应的碳同位素值;Δorg代表两类碳同位素值之间的幅度;Fextra代表其他来源的碳输入,δextra为其相应的碳同位素值。
此外,Jost等在碳循环基础上加入钙循环(即钙同位素数据),提出公式[71]
(4)
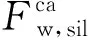
式(4)可以研究中部大西洋岩浆省火山活动释放的大量CO2导致海水酸化对钙循环的影响,从而更全面地了解三叠系—侏罗系界线的情况。
3 三叠系—侏罗系界线的全球关联性
三叠纪—侏罗纪之交引发的环境变化影响范围很广,生物种群变化及碳同位素值偏移已在全球多个剖面中找到。为了较好地呈现各剖面三叠系—侏罗系界线的关联性,本文收集了多地更为稳定的有机碳同位素值与生物种群情况。
Guex等在美国内华达州Nevada剖面发现初始有机碳同位素值负偏移与三叠系菊石的灭绝位置相符,同时赫塘阶底部的碳同位素值负偏移及主要碳同位素值负偏移与大多数陆地植物及菊石、有孔虫的恢复期相对应,三叠系—侏罗系界线因此被确定为侏罗纪菊石(Psilocerasspelae)首次出现的位置,该段主要沉积黑色页岩[18]。在英国St Audrie剖面及丹麦Stenlille剖面中并没有发现明显的生物学证据,但在初始有机碳同位素值负偏移上的正偏移中发现了大量侏罗纪孢粉(P.polymicroforatusabundance)聚集,因此,Lindstrom等将界线位置标志于此[23];该段St Audrie剖面沉积薄层泥岩,偶见灰岩夹层[5],Stenlille剖面沉积灰色粉砂岩[58]。特提斯洋范围内的奥地利Kuhjoch剖面初始碳同位素值负偏移的位置与菊石(Choristocerasmarchii)灭绝位置相对应,在随后的碳同位素值正偏移及主要碳同位素值负偏移中,孢粉种群逐渐恢复;而侏罗纪菊石(Psilocerasspelae)首次出现位置为主要有机碳同位素值负偏移之上的正偏移内,为三叠系—侏罗系界线位置[72]。该剖面也是国际地质科学联合会(IUGS)批准的侏罗系底界全球年代地层单位界线层型剖面和点位(GSSP),该段沉积黏土-粉砂状泥灰岩。意大利伦巴第(Lombardy)盆地Val Adara剖面中菊石证据及孢粉证据均不完善,但三叠系—侏罗系界线处的剖面岩性变化十分明显,在出现初始碳同位素值负偏移之下的瑞替阶主要沉积物为富含化石(尤其珊瑚)的泥晶颗粒灰岩,在碳同位素值负偏移处岩性突变为薄层状泥岩,在随后的赫塘阶连续碳同位素值正偏移中则对应鲕粒灰岩沉积[57]。相同的岩性变化及同位素特征(即鲕粒灰岩与连续碳同位素值正偏移)也出现在阿联酋Wadi Milaha剖面中,但同位素类型为无机碳[26,35](图7)。

图7 全球沉积三叠系—侏罗系界线的典型剖面对比Fig.7 Comparison of Triassic-Jurassic Boundaries from Global Typical Profiles
现阶段中国三叠系—侏罗系界线的研究取得了一些进展,主要以岩性描述及古生物化石研究为主,根据三叠纪—侏罗纪古地理环境可分为海相剖面(西藏地区及广东部分地区)、海陆交互相剖面(湘粤地区)及陆相剖面(除上述地区外的国内大部分地区)。①海相剖面研究取得进展最多。王永胜等利用菊石、珊瑚、腕足类、鱼类及双壳类建立西藏北部多玛地层的晚三叠世—早侏罗世生物地层划分标准[73];阴家润等对西藏格米格剖面进行了详细的菊石研究,并提供了碳氧同位素数据[9-10];Sha等更是在羌塘盆地取得突破性进展,提出了双壳类生物Waagenoperna不仅在本次灭绝事件中存活,同时该生物各亚属聚集分类还可细致区分晚三叠世—早侏罗世各地质时间段,其中卡尼阶—诺利阶Waagenoperna的A类亚属聚集甚至可以同日本及南阿尔卑斯山脉的同类生物相关联[74]。不仅如此,羌塘盆地中三叠世—侏罗纪发生的相关地质现象还可和天文学结合起来,已有部分数据表明该时期剧烈的气候变化可能由行星运行轨道的偏心率周期引起,其与太阳系的混沌扩散有关[16]。②海陆交互相剖面研究近几年来也取得了较大突破。张彦伟等提出了南岭地区的晚三叠世—早侏罗世海相古生物标志带[11];许中杰等利用地球化学元素分析推测华南陆缘于三叠纪—侏罗纪之交出现湿润→短暂干旱→湿润的气候变化及早侏罗世大规模海侵事件具有全球一致性[75]。③陆相剖面研究则更注重利用孢粉类型的变化划分界线及研究相关环境变化。卢远征等利用准噶尔盆地南缘孢粉类型及组合特征判断早侏罗世更为潮湿炎热,同时提供有机碳同位素值,并与全球典型剖面作对比[8];杜建军等则应用地球化学数据推测柴达木盆地早侏罗世环境为贫氧还原环境[76];刘兆生等则对四川盆地晚三叠世孢粉组合及相关古气候和古植物变化进行详细研究,认为沉积相类型也多以河流及湖泊为主[7,77]。
4 结 语
(1)三叠纪—侏罗纪转折期由于受中部大西洋岩浆省火山活动的影响,发生了海平面波动和海水酸化,长期温室效应导致气候炎热,还伴随晚三叠世末的生物灭绝事件,同时碳同位素值也受到影响,这些在全球范围内各剖面上及其地球化学数据中显示出相应的变化。
(2)现阶段划分三叠系—侏罗系界线的方法主要有以菊石及孢粉为主的古生物化石标定法、碳同位素标定法、元素地球化学特征分析法和综合数据建立地质模型等,已经被用于多个国家的三叠系—侏罗系界线研究中。
(3)目前,中国对于三叠系—侏罗系界线的研究尚不全面,究其原因主要有:①中国沉积环境大部分属于陆相沉积环境,不同于现阶段全球研究热点的中、高纬度海相环境中有较为丰富的海相生物化石证据;②东亚地区距离中部大西洋岩浆省大规模频发的火山活动较远,因此,环境变化受其影响相对来说不如欧洲和北美洲等。
(4)中国三叠系—侏罗系界线研究仍缺乏地球化学数据及整合各方面数据进行地质建模等定量化研究。未来对于三叠系—侏罗系地层划分的研究应合理利用半定量的生物化石,结合地球化学数据(如碳同位素、钙同位素、其他重要元素分布特征)等建立地质模型。这不仅可以更精确地厘定三叠系—侏罗系界线,与国际年代地层格架相对应,同时还可用于推测该时期沉积环境的变化,补充全球三叠系—侏罗系界线研究中东亚方面的证据。
参考文献:
References:
[1] HESSELBO S P,MCROBERTS C A,PALFY J.Triassic-Jurassic Boundary Events:Problems,Progress,Possibilities[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2007,244(1/2/3/4):1-10.
[2] HESSELBO S P,ROBINSON S A,SURLYK F,et al.Terrestrial and Marine Extinction at the Triassic-Jurassic Boundary Synchronized with Major Carbon-cycle Perturbation:A Link to Initiation of Massive Volcanism?[J].Geology,2002,30(3):251-254.
[3] MCELWAIN J C,BEERLING D J,WOODWARD F I.Fossil Plants and Global Warming at the Triassic-Jurassic Boundary[J].Science,1999,285:1386-1390.
[4] TANNER L H,LUCAS S G,CHAPMAN M G.Assessing the Record and Causes of Late Triassic Extinctions[J].Earth-science Reviews,2004,65(1/2):103-139.
[5] HESSELBO S P,ROBINSON S A,SURLYK F.Sea-level Change and Facies Development Across Potential Triassic-Jurassic Boundary Horizons,SW Britain[J].Journal of the Geological Society,2004,161(3):365-379.
[6] 刘兆生.塔里木盆地北缘晚三叠世孢粉组合及三叠系—侏罗系界线[J].地层学杂志,1999,23(2):96-106.
LIU Zhao-sheng.Palynological Assemblage of the Late Triassic Tariqike Formation and Triassic-Jurassic Boundary on the Northern Margin of the Tarim Basin,Xinjiang[J].Journal of Stratigraphy,1999,23(2):96-106.
[7] 刘兆生,李丽琴,王永栋.重庆合川地区晚三叠世须家河组孢粉组合[J].古生物学报,2015,54(3):279-304.
LIU Zhao-sheng,LI Li-qin,WANG Yong-dong.Late Triassic Spore-pollen Assemblage from the Xujiahe Formation in Hechuan of Chongqing,China[J].Acta Palaeontologica Sinica,2015,54(3):279-304.
[8] 卢远征,邓胜徽.准噶尔盆地南缘三叠纪—侏罗纪之交的古气候[J].古地理学报,2009,11(6):652-660.
LU Yuan-zheng,DENG Sheng-hui.Palaeoclimate Around the Triassic-Jurassic Boundary in Southern Margin of Junggar Basin[J].Journal of Palaeogeography,2009,11(6):652-660.
[9] 阴家润,FURSICH F T.西藏喜马拉雅地区三叠系—侏罗系界线动物群扩散事件及古环境[J].中国科学:D辑,地球科学,2009,39(9):1232-1238.
YIN Jia-run,FURSICH F T.Dispersal Events of Triassic-Jurassic Boundary Faunas,and Palaeoenvironment of Tibetan Himalaya[J].Science in China:Series D,Earth Sciences,2009,39(9):1232-1238.
[10] 阴家润,蔡华伟,周志广,等.西藏海相三叠系—侏罗系界线及晚三叠世生物绝灭事件研究[J].地学前缘,2006,13(4):244-254.
YIN Jia-run,CAI Hua-wei,ZHOU Zhi-guang,et al.Study of Marine Triassic-Jurassic Boundary Stratigraphy and the Latest Triassic Mass Extinction in Tibet[J].Earth Science Frontiers,2006,13(4):244-254.
[11] 张彦伟,姜宝玉,牛亚卓.南岭地区上三叠统—下侏罗统双壳类生物地层及三叠—侏罗系界线[J].高校地质学报,2014,20(4):570-581.
ZHANG Yan-wei,JIANG Bao-yu,NIU Ya-zhuo.Study on the Upper Triassic-Lower Jurassic Bivalve Biostratigraphy and the Triassic-Jurassic Boundary in the Nanling Region,South China[J].Geological Journal of China Universities,2014,20(4):570-581.
[12] GOLONKA J.Late Triassic and Early Jurassic Palaeo-geography of the World[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2007,244(1/2/3/4):297-307.
[13] GOLONKA J,FORD D.Pangean (Late Carboniferous-Middle Jurassic) Paleoenvironment and Lithofacies[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2000,161(1/2):1-34.
[14] GREENE S E,MARTINDALE R C,RITTERBUSH K A,et al.Recognising Ocean Acidification in Deep Time:An Evaluation of the Evidence for Acidification Across the Triassic-Jurassic Boundary[J].Earth-science Reviews,2012,113(1/2):72-93.
[15] MARZOLI A,BERTRAND H,KNIGHT K,et al.Synchrony of the Central Atlantic Magmatic Province and the Triassic-Jurassic Boundary Climatic and Biotic Crisis[J].Geology,2004,32(11):973-976.
[16] SHA J G,OLSEN P E,PAN Y H,et al.Triassic-Jurassic Climate in Continental High-latitude Asia Was Dominated by Obliquity-paced Variations (Junggar Basin,Urumqi,China)[J].PNAS,2015,112(12):3624-3629.
[17] TROTTER J A,WILLIAMS I S,NICORA A,et al.Long-term Cycles of Triassic Climate Change:A New δ18O Record from Conodont Apatite[J].Earth and Planetary Science Letters,2015,415:165-174.
[18] GUEX J,BARTOLINI A,ATUDOREI V,et al.High-resolution Ammonite and Carbon Isotope Stratigraphy Across the Triassic-Jurassic Boundary at New York Canyon (Nevada)[J].Earth and Planetary Science Letters,2004,225(1/2):29-41.
[19] KORTE C,HESSELBO S P,JENKYNS H C,et al.Palaeoenvironmental Significance of Carbon- and Oxygen-isotope Stratigraphy of Marine Triassic-Jurassic Boundary Sections in SW Britain[J].Journal of the Geological Society,2009,166(3):431-445.
[20] MCROBERTS C A,KRYSTYN L,HAUTMANN M.Macrofaunal Response to the End-Triassic Mass Extinction in the West-Tethyan Kossen Basin,Austria[J].Palaios,2012,27(9/10):607-616.
[21] COHEN K M,FINNEY S C,GIBBARD P L,et al.The ICS International Chronostratigraphic Chart[J].Episodes,2013,36(3):199-204.
[22] HALLAM A,WIGNALL P B.Mass Extinctions and Sea-level Changes[J].Earth-science Reviews,1999,48(4):217-250.
[23] LINDSTROM S,VAN DE SCHOOTBRUGGE B,HANSEN K H,et al.A New Correlation of Triassic-Jurassic Boundary Successions in NW Europe,Nevada and Peru,and the Central Atlantic Magmatic Pro-vince:A Time-line for the End-Triassic Mass Extinction[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2017,478:80-102.
[24] PRETO N,KUSTATSCHER E,WIGNALL P B,et al.Triassic Climates:State of the Art and Perspectives[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2010,290(1/2/3/4):1-10.
[25] HALLAM A.Estimates of the Amount and Rate of Sea-level Change Across the Rhaetian Hettangian and Pliensbachian Toarcian Boundaries (Latest Triassic to Early Jurassic)[J].Journal of the Geological Society,1997,154(5):773-779.
[26] HONIG M R,JOHN C M,MANNING C.Development of an Equatorial Carbonate Platform Across the Triassic-Jurassic Boundary and Links to Global Palaeo-environmental Changes (Musandam Peninsula,UAE/Oman)[J].Gondwana Research,2017,45:100-117.
[27] KUERSCHNER W M,BONIS N R,KRYSTYN L.Carbon-isotope Stratigraphy and Palynostratigraphy of the Triassic-Jurassic Transition in the Tiefengraben Section-northern Calcareous Alps (Austria)[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2007,244(1/2/3/4):257-280.
[28] VAN DE SCHOOTBRUGGE B,TREMOLADA F,>ROSENTHAL Y,et al.End-Triassic Calcification Crisis and Blooms of Organic-walled ‘Disaster Species’[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2007,244(1/2/3/4):126-141.
[29] MCROBERTS C A,FURRER H,JONES D S.Palaeoenvironmental Interpretation of a Triassic-Jurassic Boundary Section from Western Austria Based on Palaeoecological and Geochemical Data[J].Palaeo-geography,Palaeoclimatology,Palaeoecology,1997,136(1/2/3/4):79-95.
[30] BONIS N R,RUHL M,KURSCHNER W M,et al.Climate Change Driven Black Shale Deposition During the End-Triassic in the Western Tethys[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2010,290(1/2/3/4):151-159.
[31] PAYNE J L,VAN DE SCHOOTBRUGGE B.Life in Triassic Oceans:Links Between Planktonic and Benthic Recovery and Radiation[M]∥FALOWSKI P G,KNOLL A H.Evolution of Primary Producers in the Sea.Burlington:Elsevier,2007:165-189.
[32] RUHL M,KURSCHNER W M,KRYSTYN L.Triassic-Jurassic Organic Carbon Isotope Stratigraphy of Key Sections in the Western Tethys Realm (Austria)[J].Earth and Planetary Science Letters,2009,281(3/4):169-187.
[33] HAUTMANN M,STILLER F,CAI H,et al.Extinction-recovery Pattern of Level-bottom Faunas Across the Triassic-Jurassic Boundary in Tibet:Implications for Potential Killing Mechanisms[J].Palaios,2008,23(10):711-718.
[34] KIESSLING W,ABERHAN M,BRENNEIS B,et al.Extinction Trajectories of Benthic Organisms Across the Triassic-Jurassic Boundary[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2007,244(1/2/3/4):201-222.
[35] AL-SUWAIDI A H,STEUBER T,SUAREZ M B.The Triassic-Jurassic Boundary Event from an Equatorial Carbonate Platform (Ghalilah Formation,United Arab Emirates)[J].Journal of the Geological Society,2016,173(6):949-953.
[36] PALFY J,DEMENY A,HAAS J,et al.Carbon Isotope Anomaly and Other Geochemical Changes at the Triassic-Jurassic Boundary from a Marine Section in Hungary[J].Geology,2001,29(1):1047-1050.
[37] WHITESIDE J H,OLSEN P E,EGLINTON T,et al.Compound-specific Carbon Isotopes from Earth’s Largest Flood Basalt Eruptions Directly Linked to the End-Triassic Mass Extinction[J].PNAS,2010,107(15):6721-6725.
[38] SCHALTEGGER U,GUEX J,BARTOLINI A,et al.Precise U-Pb Age Constraints for End-Triassic Mass Extinction,Its Correlation to Volcanism and Hettan-gian Post-extinction Recovery[J].Earth and Planetary Science Letters,2008,267(1/2):266-275.
[39] WIGNALL P B,ZONNEVELD J P,NEWTON R J,et al.The End Triassic Mass Extinction Record of Williston Lake,British Columbia[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2007,253(3/4):385-406.
[40] HAUTMANN M.Effect of End-Triassic CO2Maximum on Carbonate Sedimentation and Marine Mass Extinction[J].Facies,2004,50(2):257-261.
[41] QUAN T M,VAN DE SCHOOTBRUGGE B,FIELD M P,et al.Nitrogen Isotope and Trace Metal Analyses from the Mingolsheim Core (Germany):Evidence for Redox Variations Across the Triassic-Jurassic Boundary[J].Global Biogeochemical Cycles,2008,22(2):GB2014.
[42] KASPRAK A H,SEPULVEDA J,PRICE-WALDMAN R,et al.Episodic Photic Zone Euxinia in the Northeastern Panthalassic Ocean During the End-Triassic Extinction[J].Geology,2015,43(4):307-310.
[43] TRECALLI A,SPANGENBERG J,ADATTE T,et al.Carbonate Platform Evidence of Ocean Acidification at the Onset of the Early Toarcian Oceanic Anoxic Event[J].Earth and Planetary Science Letters,2012,357/358:214-225.
[44] RIES B J.Aragonite Production in Calcite Seas:Effect of Seawater Mg/Ca Ratio on the Calcification and Growth of the Calcareous Alga Penicillus Capitatus[J].Paleobiology,2005,31(3):445-458.
[45] GALLI M T,JADOUL F,BERNASCONI S M,et al.Stratigraphy and Palaeoenvironmental Analysis of the Triassic-Jurassic Transition in the Western Southern Alps (Northern Italy)[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2007,244(1/2/3/4):52-70.
[46] CIARAPICA G.Regional and Global Changes Around the Triassic-Jurassic Boundary Reflected in the Late Norian-Hettangian History of the Apennine Basins[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2007,244(1/2/3/4):34-51.
[47] ROZIC B,KOLAR-JURKOVSEK T,SMUC A.Late Triassic Sedimentary Evolution of Slovenian Basin (Eastern Southern Alps):Description and Correlation of the Slatnik Formation[J].Facies,2009,55(1):137-155.
[48] ROZIC B.Upper Triassic-Lower Jurassic Limestones from Mt Kobla in the Northern Tolmin Basin:Tectonically Repeated or Continuous Succession?[J].RMZ-materials and Geoenvironment,2008,55(3):345-362.
[49] HORI R S,FUJIKI T,INOUE E,et al.Platinum Group Element Anomalies and Bioevents in the Triassic-Jurassic Deep-sea Sediments of Panthalassa[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2007,244(1/2/3/4):391-406.
[50] HALLAM A,WIGNALL P B,YIN J R,et al.An Investigation into Possible Facies Changes Across the Triassic-Jurassic Boundary in Southern Tibet[J].Sedi-mentary Geology,2000,137(3/4):101-106.
[51] 贺振建.准噶尔盆地四棵树剖面孢粉组合及三叠系—侏罗系界线[J].地层学杂志,2007,31(增2):513-520.
HE Zhen-jian.Sporopollen Assemblages and the Triassic-Jurassic Boundary at the Sikeshu Section of the Junggar Basin,Xinjiang[J].Journal of Stratigraphy,2007,31(S2):513-520.
[52] 孟繁松,陈辉明,李旭兵.四川盆地非海相三叠系—侏罗系界线研究[J].地层学杂志,2005,29(增1):565-572.
MENG Fan-song,CHEN Hui-ming,LI Xu-bing.Study of the Non-marine Triassic-Jurassic Boundary in the Sichuan Basin[J].Journal of Stratigraphy,2005,29(S1):565-572.
[53] FELBER R,WEISSERT H J,FURRER H,et al.The Triassic-Jurassic Boundary in the Shallow-water Marine Carbonates from the Western Northern Calcareous Alps (Austria)[J].Swiss Journal of Geosciences,2015,108(2/3):213-224.
[54] GALLI M T,JADOUL F,BERNASCONI S M,et al.Anomalies in Global Carbon Cycling and Extinction at the Triassic/Jurassic Boundary:Evidence from a Marine C-isotope Record[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2005,216(3/4):203-214.
[55] WILLIFORD K H,WARD P D,GARRISON G H,et al.An Extended Organic Carbon-isotope Record Across the Triassic-Jurassic Boundary in the Queen Charlotte Islands,British Columbia,Canada[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2007,244(1/2/3/4):290-296.
[56] PALFY J,DEMENY A,HAAS J,et al.Triassic-Jurassic Boundary Events Inferred from Integrated Stratigraphy of the Csovar Section,Hungary[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2007,244(1/2/3/4):11-33.
[57] BACHAN A,VAN DE SCHOOTBRUGGE B,FIEBIG J,et al.Carbon Cycle Dynamics Following the End-Triassic Mass Extinction:Constraints from Paired δ13Ccarband δ13CorgRecords[J].Geochemistry,Geophysics,Geosystems,2012,13(9):Q09008.
[58] LINDSTROM S,VAN DE SCHOOTBRUGGE B,DYBKJAER K,et al.No Causal Link Between Terrestrial Ecosystem Change and Methane Release During the End-Triassic Mass-extinction[J].Geology,2012,40(6):531-534.
[59] LEHMANN M F,BERNASCONI S M,BARBIERI A,et al.Preservation of Organic Matter and Alteration of Its Carbon and Nitrogen Isotope Composition During Simulated and In-situ Early Sedimentary Diagenesis[J].Geochimica et Cosmochimica Acta,2002,66(20):3573-3584.
[60] BACHAN A,PAYNE J L.Modelling the Impact of Pulsed CAMP Volcanism onpCO2and δ13C Across the Triassic-Jurassic Transition[J].Geological Magazine,2016,153(2):252-270.
[61] DAL CORSO J,MARZOLI A,TATEO F,et al.The Dawn of CAMP Volcanism and Its Bearing on the End-Triassic Carbon Cycle Disruption[J].Journal of the Geological Society,2014,42(3):235-238.
[62] FAURE G,MENSING T M.Isotopes:Principles and Applications[M].3rd ed.Hoboken:John Wiley and Sons Inc.,2005.
[63] RUHL M,KURSCHNER W M.Multiple Phases of Carbon Cycle Disturbance from Large Igneous Pro-vince Formation at the Triassic-Jurassic Transition[J].Geology,2011,39(5):431-434.
[64] RUHL M,BONIS N R,REICHART G J,et al.Atmospheric Carbon Injection Linked to End-Triassic Mass Extinction[J].Science,2011,333:430-434.
[65] PETERSEN H I,LINDSTROM S.Synchronous Wildfire Activity Rise and Mire Deforestation at the Triassic-Jurassic Boundary[J].Plos One,2012,7(10):e47236.
[66] 邓胜徽,卢远征,樊 茹,等.中国陆相三叠系—侏罗系界线[J].地层学杂志,2013,37(4):582-584.
DENG Sheng-hui,LU Yuan-zheng,FAN Ru,et al.Triassic-Jurassic Boundary of China Terrestrial Sections[J].Journal of Stratigraphy,2013,37(4):582-584.
[67] 吴智平,马在平,周瑶琪.济源盆地三叠系与侏罗系界线地层沉积相及元素地球化学特征[J].石油大学学报:自然科学版,2002,26(3):20-25.
WU Zhi-ping,MA Zai-ping,ZHOU Yao-qi.Element Geochemical Characters of Sedimentary Facies in the Triassic-Jurassic Boundary Section of Jiyuan Basin[J].Journal of the University of Petroleum,China:Natural Science Edition,2002,26(3):20-25.
[68] KURODA J,HORI R S,SUZUKI K,et al.Marin Osmium Isotope Record Across the Triassic-Jurassic Boundary from a Pacific Pelagic Site[J].Geology,2010,38(12):1095-1098.
[69] TANNER L H,KYTE F T.Anomalous Iridium Enrichment at the Triassic-Jurassic Boundary,Blomidon Formation,Fundy Basin,Canada[J].Earth and Planetary Science Letters,2005,240(3/4):634-641.
[70] KUMP L R,ARTHUR M A.Interpreting Carbon-isotope Excursions:Carbonates and Organic Matter[J].Chemical Geology,1999,161(1/2/3):181-198.
[71] JOST A B,BACHAN A,VAN DE SCHOOTBRUGGE B,et al.Additive Effects of Acidification and Minera-logy on Calcium Isotopes in Triassic/Jurassic Boundary Limestones[J].Geochemistry,Geophysics,Geosystems,2017,18(1):113-124.
[72] HILLEBRANDT A V,KRYSTYN L,KURSCHNER W M,et al.The Global Stratotype Sections and Point (GSSP) for the Base of the Jurassic System at Kuhjoch (Karwendel Mountains,Northern Calcareous Alps,Tyrol,Austria)[J].Episodes,2013,36(3):162-198.
[73] 王永胜,郑春子.藏北色哇地区索布查组、曲色组岩石地层、层序地层、生物地层特征及三叠系与侏罗系界线[J].地层学杂志,2007,31(4):377-384.
WANG Yong-sheng,ZHENG Chun-zi.Lithostratigraphy,Sequence Stratigraphy,and Biostratigraphy of the Suobucha and Quse Formations and the Triassic-Jurassic Boundary in the Sewa Area on the South Margin of the Qiangtang Basin,Northern Tibet[J].Journal of Stratigraphy,2007,31(4):377-384.
[74] SHA J G,WANG Y Q,PAN Y H,et al.Temporal and Spatial Distribution Patterns of the Marine-brackish-water BivalveWaagenopernain China and Its Implications for Climate and Palaeogeography Through the Triassic-Jurassic Transition[J].Palaeogeography,Palaeoclimatology,Palaeoecology,2016,464:43-50.
[75] 许中杰,程日辉,沈艳杰,等.闽西南地区晚三叠—早侏罗世温湿—干热气候转变的沉积记录[J].中国矿业大学学报,2012,41(5):783-792.
XU Zhong-jie,CHENG Ri-hui,SHEN Yan-jie,et al.Sedimentary Records of the Climatic Transition from Warm and Humid to Dry and Hot During Late Triassic-Early Jurassic in Southwestern Fujian[J].Journal of China University of Mining and Technology,2012,41(5):783-792.
[76] 杜建军,张士安,肖伟峰,等.柴达木盆地北缘中—下侏罗统碎屑岩地球化学特征及其地质意义[J].地球科学与环境学报,2017,39(6):721-734.
DU Jian-jun,ZHANG Shi-an,XIAO Wei-feng,et al.Geochemistry Characteristics of Middle-Lower Jurassic Clastic Rocks in the Northern Margin of Qaidam Basin and Their Geological Significance[J].Journal of Earth Sciences and Environment,2017,39(6):721-734.
[77] 刘兆生,李丽琴,王永栋.四川宣汉晚三叠世孢粉组合及其古植被与古气候意义[J].微体古生物学报,2015,32(1):43-62.
LIU Zhao-sheng,LI Li-qin,WANG Yong-dong.Late Traissic Spore-pollen Assemblage from Xuanhan of Sichuan,China[J].Acta Micropalaeontologica Sinica,2015,32(1):43-62.
猜你喜欢
小资CHIC!ELEGANCE(2021年40期)2021-11-08
汉语世界(The World of Chinese)(2021年1期)2021-02-22
分析化学(2019年3期)2019-03-30
分析化学(2017年5期)2017-06-21
学苑创造·A版(2017年5期)2017-05-09
中学生理科应试(2017年2期)2017-04-01
分析化学(2016年9期)2016-05-14
海峡姐妹(2016年7期)2016-02-27
中学政史地·初中(2008年8期)2008-10-18
