
滇西北石卡雪山2个针叶树种森林上限径向生长对温度和降水的响应
2018-05-16 08:40尹定财李丽萍张卫国
生态学报 2018年7期
张 贇,尹定财,孙 梅,李丽萍,田 昆,张卫国
西南林业大学国家高原湿地研究中心, 昆明 650224
全球变化对人类生存环境和地球生态系统的影响,已成为世界关注的热点。气候变化作为全球变化的重要组成部分,已经显著影响陆地生态系统,尤其是森林生态系统的结构和功能,森林生态系统对气候变化的响应已成为重要的科学问题[1]。树木年轮学作为研究气候变化的重要手段[2],在研究森林生态系统对气候变化响应中发挥着重要作用[3]。气候变化对森林的影响直接作用于树木的生理过程,树木每一个年轮的形成都是多个生态因子综合作用的结果[4]。在去除树木自身生长趋势和其他非气候因素的影响后,年轮指数将保留大量过去气候变化的信息[5],因此开展树木年轮与气候因子关系的研究,对揭示影响树木径向生长的关键气候因子、加强森林生态系统的保护与管理具有重要意义。
森林上限及纬度分布北界均属于气候变化敏感区,森林上限的树木处在环境胁迫的临界状态,因而该区域树木径向生长对气候变化的响应研究受到高度关注[6]。通常认为,森林上限树木径向生长主要受温度的限制[7],然而,由于受地区气候差异影响,这一规律并不是在所有地区都适用[8- 9]。即使同一地区,不同树种径向生长与气候变化的关系也存在差异[10]。因此,探讨同一地区不同树种径向生长对气候变化的响应差异,能够帮助我们深入理解树木生长对不同气候因子的响应机制,对预测未来气候变化如何影响树木生长提供科学依据。
滇西北高原位于青藏高原的东南缘,对全球气候变化敏感,森林资源丰富且保存完好,是树木年轮学研究的理想场所[11- 12]。近年来,我国学者在该区域利用树木年轮开展了气候重建研究,Fan等通过研究该区域长苞冷杉(Abiesgeorgei)和麦吊云杉(Piceabrachytyla)树木年轮,发现温度,特别是冬季温度是影响树线附近树木生长的主要气候因子,并利用线性回归的方法重建了该区域过去250年的年平均温度[13];Bi等利用丽江云杉(Picealikiangensis)树轮宽度标准化年表重建了玉龙雪山地区春季帕尔默干旱指数(PDSI)[14];Guo等发现玉龙雪山的丽江云杉和云南铁杉(Tsugadumosa)的径向生长对气候变化的响应存在很大差异,并重建了该区域5—6月的帕尔默干旱指数[15]。此外,玉龙雪山及普达措国家公园相继开展了不同树种径向生长对气候因子响应差异研究[10,16],研究结果表明树木生长受温度和降水的共同作用。香格里拉地区的石卡雪山尚未见有关树木年轮学的研究报道,采样点的增加及研究树种的扩充有利于补充和完善该地区的树木年轮学知识,从而进一步揭示影响滇西北树木生长的主要气候因子。
本研究拟以石卡雪山森林上限丽江云杉和高山松(Pinusdensata)为研究对象,上述2个树种为香格里拉地区森林组成的主要树种(代表性),且生物习性各异(差异性),主要(1)建立2个树种树木年轮年表并分析径向生长特征;(2)利用响应函数分析2个树种径向生长与温度和降水的响应关系及差异;(3)运用冗余分析(redundancy analysis, RDA)验证树木径向生长与气候因子的关系。进而补充和完善滇西北高原树木年轮资料,探讨不同树种对气候变化的响应差异,以揭示影响该地区树木生长的主要气候因子,从而为有效应对未来气候变化对该区域树木生长及森林生态系统影响提供理论依据。
1 材料与方法
1.1 研究区概况
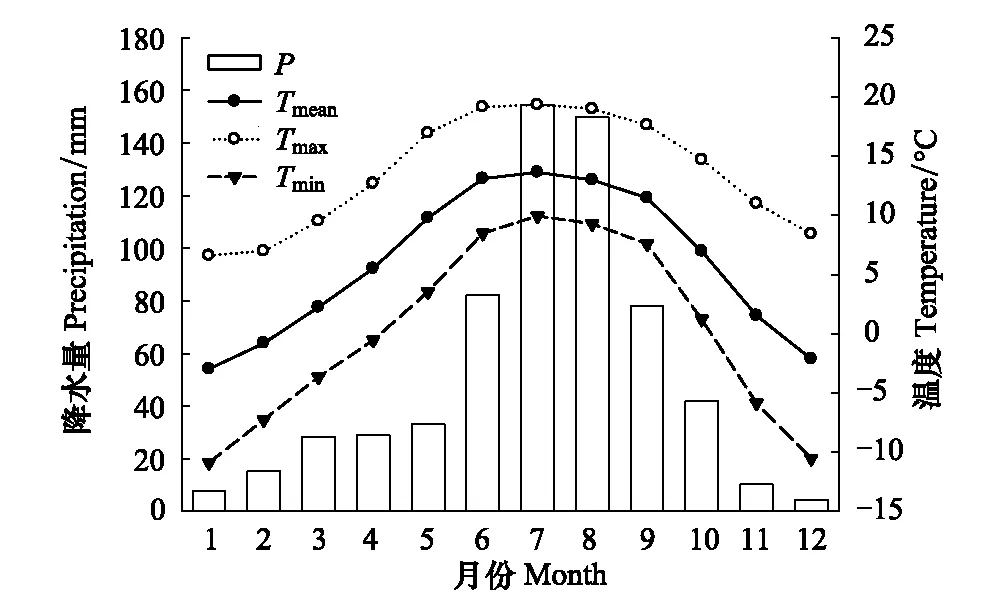
图1 香格里拉气象站1960—2011年气候资料 Fig.1 Climate data from the Shangri-La meteorological station (1960—2011)Tmean:月平均温 Monthly mean temperature;Tmax:月平均最高温 Monthly mean maximum temperature;Tmin:月平均最低温 Monthly mean minimum temperature;P:月降水量 Monthly total precipitation
石卡雪山位于香格里拉市区西南部,区域内植被垂直分布明显,物种资源丰富,森林保存完整,以丽江云杉、高山松、长苞冷杉和大果红杉(Larixpotaniniivar.macrocarpa)等针叶树种为主。丽江云杉为浅根性树种,侧根较为发达,主要分布在海拔3100—3810m之间,具有很强的耐荫能力[17];高山松为深根性树种,喜阳、耐干燥贫瘠的土壤[17],在滇西北的海拔分布范围一般多在2700—3700m之间[18]。
石卡雪山位于我国西南部季风气候区,干湿季明显,昼夜温差大,气候具有雨热同期的特点。根据香格里拉气象站1960—2011年的气候数据资料,其年平均温为5.9℃,7月最热,为13.6℃,1月最冷,为-3.0℃;年降水量为634mm,时间分配不均,主要集中在6—9月,占全年降水量的73%(图1)。
1.2 样品采集与年表建立
2015年8月和2016年4月,在石卡雪山分别采集丽江云杉和高山松森林上限的树木年轮样本。取样时,尽量选择树龄较大、生长良好的优势木,在树高1.3m处,从不同方位采集1—2个样芯。同时,为保证样本所含气候信息的一致性,同一树种的采样点高差被控制在10m以内。将取到的样芯放入塑料吸管内,并编号。2个树种共采集52棵树,101个样芯(表1)。

表1 树木年轮采样点概况
*分子数据代表株树,分母数据代表芯数
样品带回实验室,按照Stokes等阐述的基本方法对样品进行预处理[19]。经自然风干、固定、打磨处理后,将样品放在双筒显微镜下初步定年,然后放于EPSON Scan(Expression 11000XL)扫描仪上进行扫描,扫描参数设置为图像类型24-位全彩,分辨率3200dpi,扫描的年轮图片用软件CDendro and CooRecorder ver. 7.3[20]测量年轮宽度,该系统精度为0.001mm。最后利用COFECHA程序[21]交叉定年和对测量结果进行检验,并剔除与主序列相关性低的样芯,最终剩余49棵树,85个样芯进入主序列。运用ARSTAN程序[22]采用67%的样条函数进行拟合,去除树木自身遗传因子所产生的生长趋势,最终建立了石卡雪山丽江云杉和高山松的标准化年表(STD)、差值年表(RES,图2)和自回归年表(ARS)。从年表统计特征值可知,丽江云杉和高山松差值年表的统计特征值普遍高于标准化年表(表2),因此,本研究利用差值年表与气候因子进行响应分析。
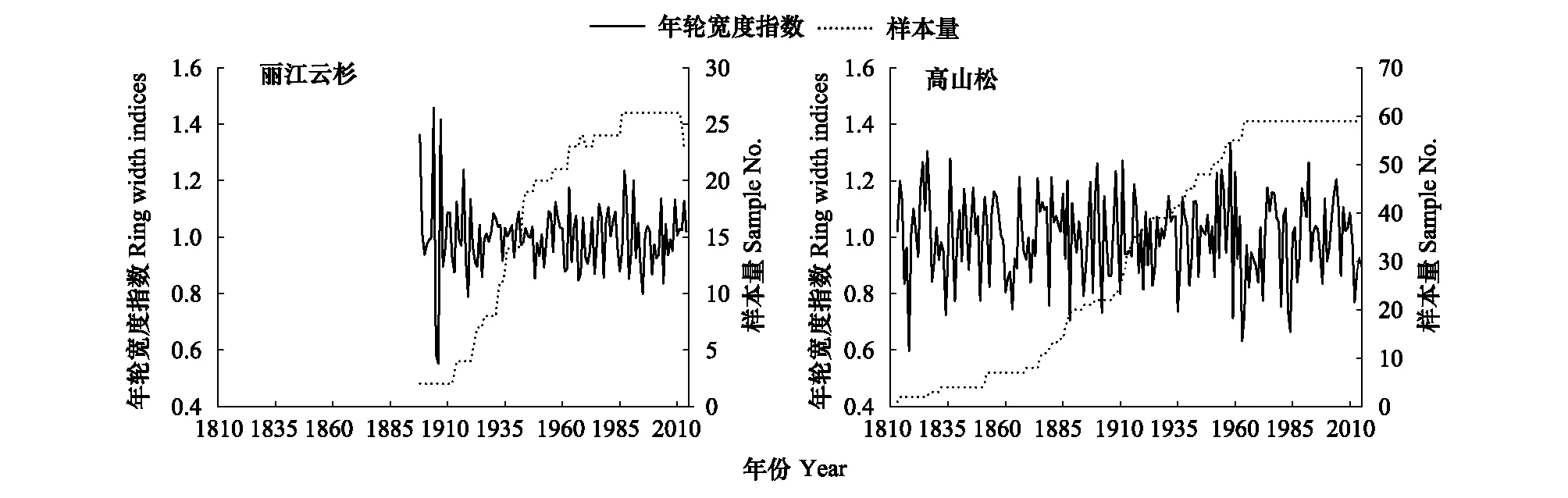
图2 差值年表(实线)和样本量(虚线)Fig.2 Residual tree-ring chronology (solid line) and sample size (dotted line)
1.3 气象数据采集与处理
气象数据选取距采样点最近的香格里拉气象站(99°42′E,27°50′N,海拔3276.7m,中国气象科学数据共享服务网:http://cdc.cma.gov.cn)气象数据资料。气候变量选取香格里拉气象站1960—2011年的月平均温(Tmean)、月平均最高温(Tmax)、月平均最低温(Tmin)和月降水量(P),从上年7月至当年10月,共计64个气候变量。为了检验气候数据的均一性,采用Mann-Kendall法[23]对气象数据进行突变检验,结果显示气象数据通过置信度为0.05的显著性水平,检验结果表明,气象数据稳定可靠,可以满足气候因子与年轮指数的响应分析和冗余分析。
1.4 数据处理
利用树木年轮学的专业软件DendroClim2002[24],运用响应函数分析年轮指数与香格里拉气象站1960—2011年的气候数据(温度和降水)的相关关系。响应函数是一种相关性分析,通过主成分分析去除气候因子的自相关,再与年表做逐步回归,并将各主成分量的回归系数转换成对应原始气候资料的回归系数,以回归系数的大小和正负表示树木生长对气候因子的响应程度和关系,常用于树木径向生长与气候因子相关分析中[6]。树木径向生长与气候因子的动态关系由DendroClim2002中的Evolutionary and Moving Response and Correlation模块完成。采用冗余分析进一步检测树木径向生长与气候因子的关系。冗余分析是多变量环境梯度分析,通过年表与气候变量的回归和主成分分析来评价树木径向生长与气候因子的关系[25]。冗余分析用软件CANOCO 4.5完成[26]。绘图用SigmaPlot 10.0完成。
2 结果
2.1 年表特征分析
由2个树种的年表特征和公共区间统计特征分析(表2)可知,高山松差值年表的平均敏感度、第一特征向量百分比、信噪比和样本总体代表性均高于丽江云杉差值年表,说明高山松差值年表包含更多的环境信息。2个树种差值年表的样本总体代表性均超过85%,说明2个年表都能较好地代表研究区内丽江云杉和高山松径向生长的基本特征,适合树木年轮学研究。
2.2 径向生长与温度和降水的响应分析
石卡雪山丽江云杉差值年表与温度和降水的响应分析表明,温度和降水显著影响其径向生长(P<0.05),其中与上年10月平均最低温呈显著负相关,与上年11月平均最高温以及当年7月平均温、平均最高温和平均最低温呈显著正相关;与上年8月降水量和当年5月降水量呈显著负相关(图3a)。
石卡雪山高山松径向生长仅与温度达到显著相关,其中与上年10月平均温和平均最高温以及11月平均温呈显著正相关,与当年7月平均温和平均最高温呈显著负相关,而与降水未达到显著相关水平(图3b)。
2.3 径向生长与气候因子的动态关系
通过窗口为32年的滑动相关来分析丽江云杉和高山松径向生长与气候因子的动态关系。在滑动区间(1992—2011年)内,丽江云杉径向生长与上年11月平均最高温、当年7月平均温及上年8月降水相关性的稳定性较好,分析的所有年份达到显著相关,与当年5月降水、当年7月平均最高温和平均最低温在部分年份达到显著相关,与上年10月平均最低温相关性较差,仅在2010—2011年达到显著相关(表3)。高山松径向生长与上年10月平均温、当年7月平均温和平均最高温相关性的稳定性较好,分析的所有年份达到显著相关,与上年10月平均最高温在部分年份达到显著相关,和上年11月平均温只在2011年达到显著相关(表3)。
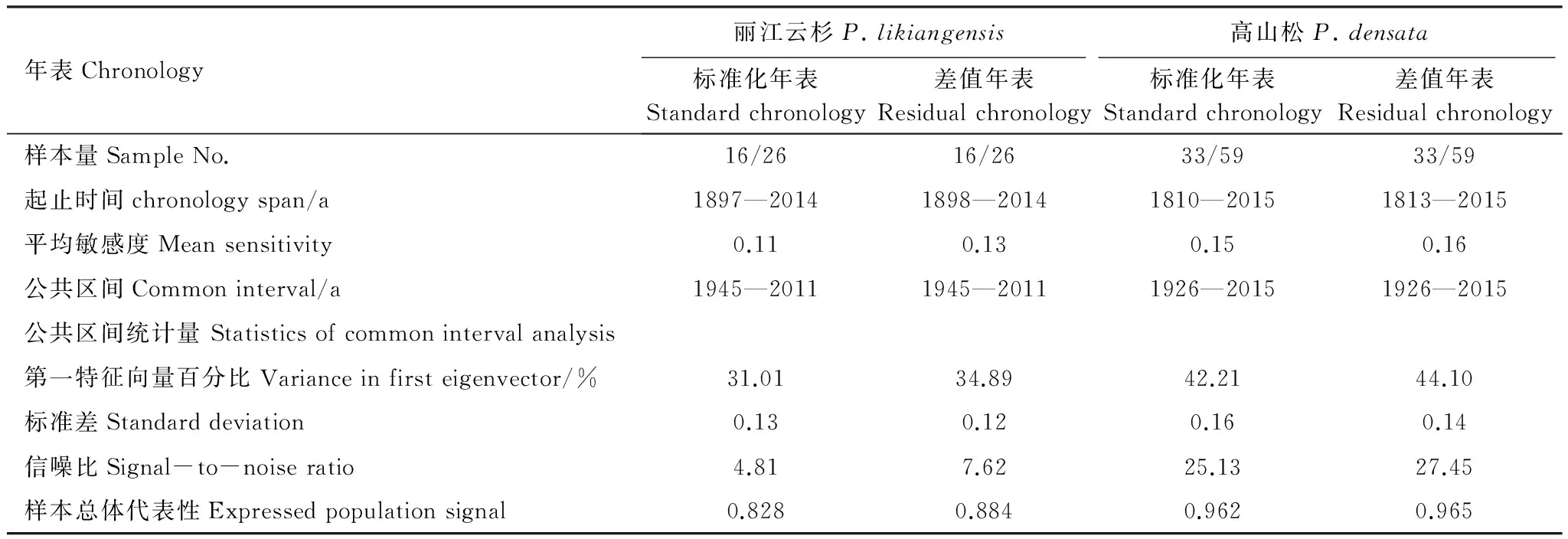
表2 树轮年表及公共区间统计量
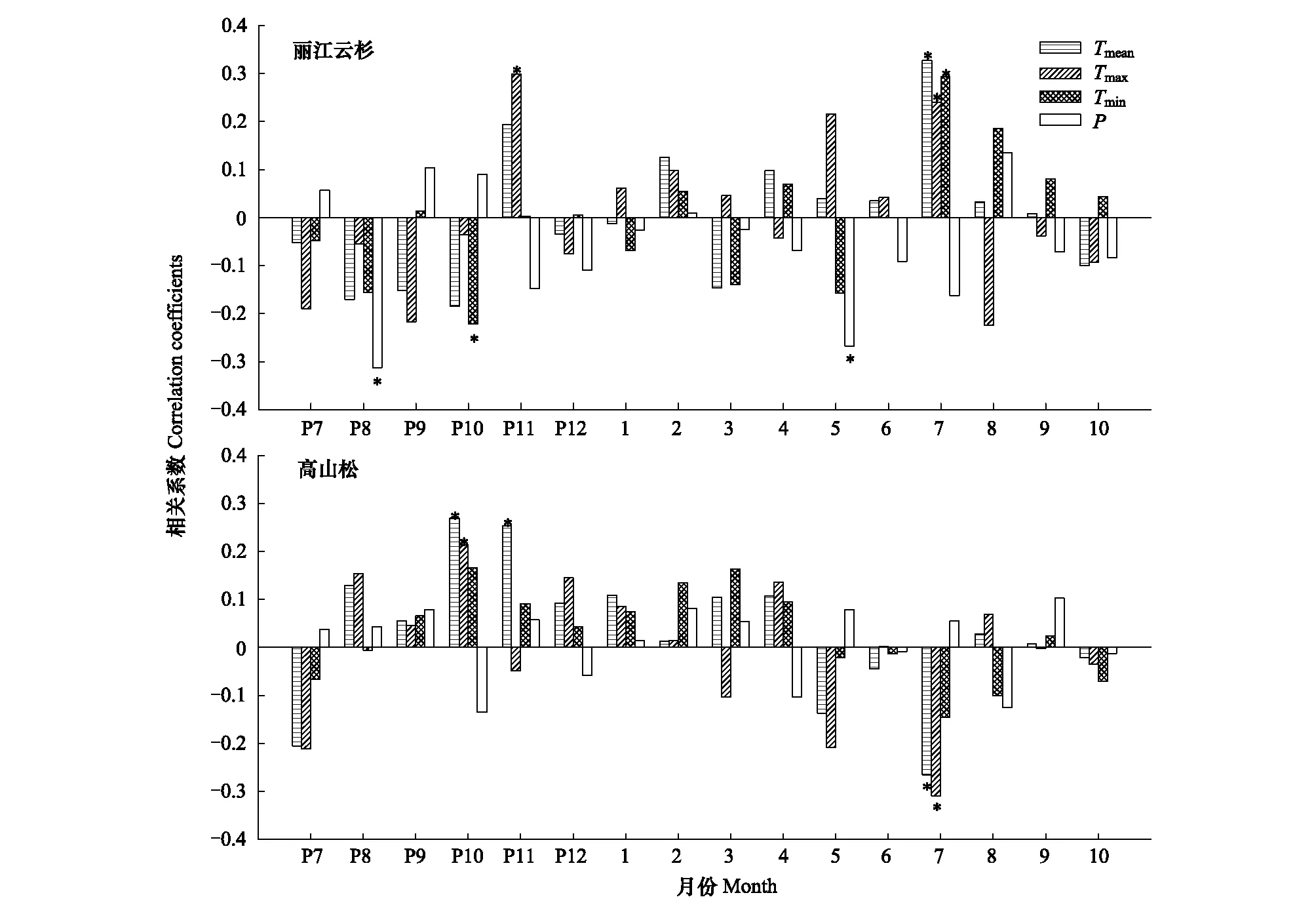
图3 差值年表与逐月气候因子的相关分析Fig.3 Correlation analyses of response function between the residual chronologies and the monthly climatic factorsP表示上一年, *表示达到0.05水平的显著相关
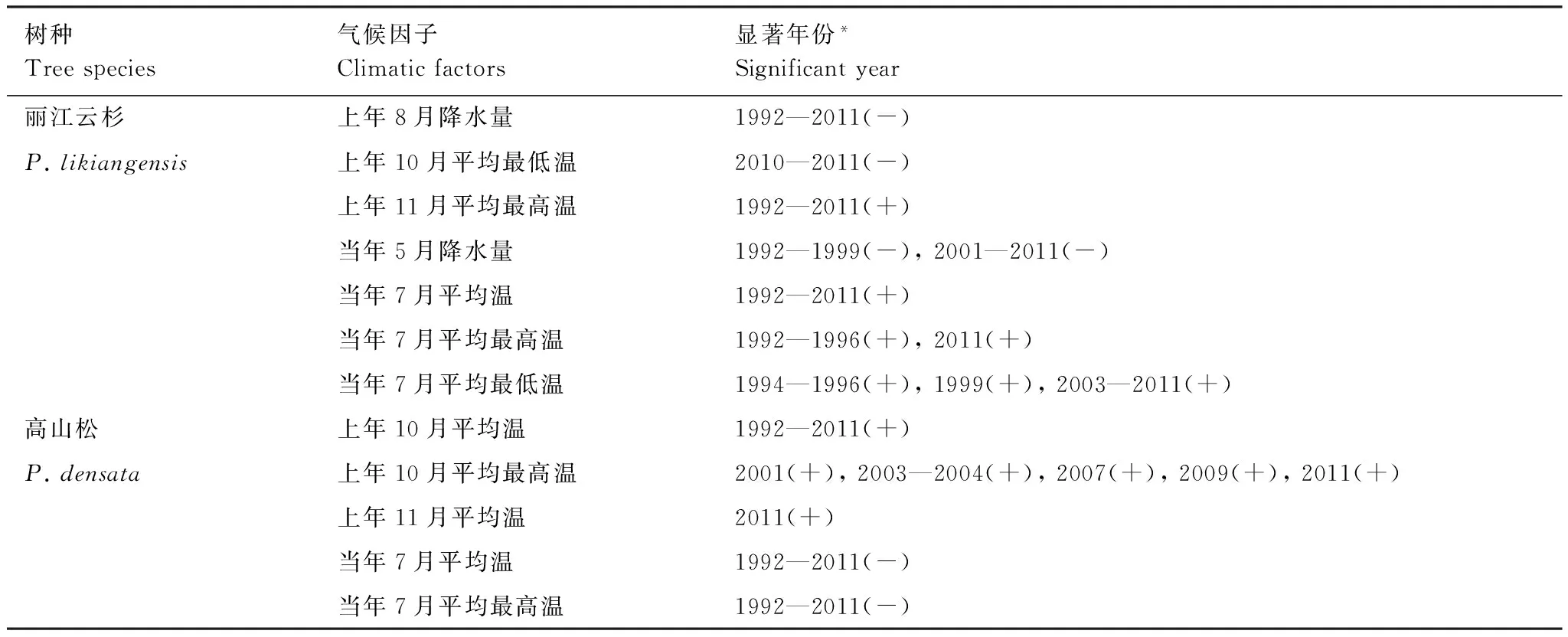
树种Treespecies气候因子Climaticfactors显著年份*Significantyear丽江云杉上年8月降水量1992—2011(-)P.likiangensis上年10月平均最低温2010—2011(-)上年11月平均最高温1992—2011(+)当年5月降水量1992—1999(-),2001—2011(-)当年7月平均温1992—2011(+)当年7月平均最高温1992—1996(+),2011(+)当年7月平均最低温1994—1996(+),1999(+),2003—2011(+)高山松上年10月平均温1992—2011(+)P.densata上年10月平均最高温2001(+),2003—2004(+),2007(+),2009(+),2011(+)上年11月平均温2011(+)当年7月平均温1992—2011(-)当年7月平均最高温1992—2011(-)
(-)表示相关关系为显著负相关;(+)表示相关关系为显著正相关;*表示达到0.05水平的显著相关
2.4 径向生长与气候因子的冗余分析
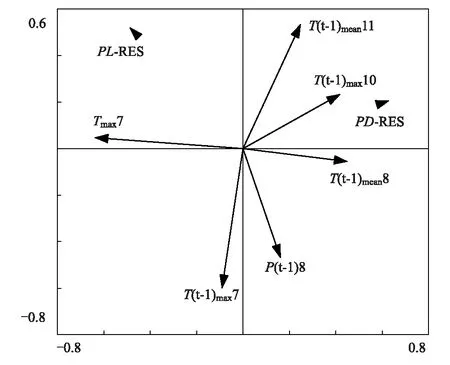
图4 差值年表与气候因子的冗余分析 Fig.4 The redundancy analysis between two residual chronologies and the climatic factors只选取显著相关(P<0.05)的气候因子;气候因子向量越长表示贡献率越大,年表点与向量(本身或延长线)垂线越短表明二者相关性越高,二者方向相同表明具有正相关关系,相反则为负相关关系;P:降水量,precipitation;Tmean:平均温,mean temperature;Tmax:平均最高温,mean maximum temperature;数字(Number)):月份,month;(t-1):前一年,the previous year;PL-RES:丽江云杉差值年表,residual chronology of Picea likiangensi;PD-RES:高山松差值年表,residual chronology of Pinus densata
丽江云杉和高山松差值年表与气候因子冗余分析结果与响应函数分析结果基本一致,进一步验证了丽江云杉和高山松径向生长与气候因子的关系。在64个气候变量中,有6个变量对石卡雪山丽江云杉和高山松径向生长的影响达到显著水平(P<0.05)。综合向量长度以及年表点到向量垂线长度可知,当年7月平均最高温(正相关)和上年8月降水(负相关)与丽江云杉年表有着很强的相关性,上年11月平均温和7月平均最高温分别对丽江云杉起着一定的促进与抑制作用。高山松径向生长主要受当年7月平均最高温(负相关)和上年10月平均最高温(正相关)影响,其次上年11月与8月平均温对其有着促进作用,上年8月降水和上年7月平均最高温对其影响相对较弱(图4)。
3 讨论
3.1 2个树种径向生长对气候响应的趋同
石卡雪山丽江云杉和高山松径向生长更多的与上年气候因子呈显著相关,这在生理学上被称为“滞后效应”[27],说明上年气候因子对该区域树木径向生长的重要作用,在相邻的小中甸[28]、普达措国家公园[16]也有温度对树木生长滞后效应的报道。上年11月的高温(平均温、平均最高温)对丽江云杉和高山松径向生长具有显著促进作用。11月的香格里拉温度较低(图1),树木已基本停止生长,但仍有较弱的生理活动,此时温度升高能够促进光合作用,有利于营养物质积累,为下一年的树木生长创造有利条件[29- 30]。
3.2 2个树种径向生长对温度响应的差异
当年7月的温度是影响2个树种径向生长的主要气候因子,但作用却相反,这与丽江云杉和高山松的生境差异有关。7月正值生长盛期,高温提高了树木的光合作用速率,制造大量营养物质,从而满足树木生长的需要[31]。但同时需要消耗大量水分,以满足植物光合作用的正常运行,若此时水分供应不足,高温反而会抑制树木的生长[32]。丽江云杉具有很强的耐荫能力,生长在阴坡、半阴坡,其土壤蒸腾蒸发较弱,水分散失小,水分能够满足丽江云杉进行光合作用。在相邻的川西色达、马尔康等地区,7月温度对川西云杉(Piceabalfourian)的促进作用也有报道[33- 34]。而高山松喜阳,生长在阳坡、半阳坡,光照充足而导致蒸发较强,高温会进一步加剧水分缺失,从而限制高山松的生长[35]。
另外,上年10月温度升高对2个树种径向生长的作用也相反, 丽江云杉表现为夜间光合产物的消耗不利于来年的生长,而高山松则表现为白天营养物质的积累促进来年生长。丽江云杉在生长季末期(10月)主要受夜间温度(最低温)影响,夜晚温度偏高,则呼吸作用加剧,消耗累积光合产物,从而限制了来年树木的生长[36]。高山松则主要受白天温度(最高温)影响,白天温度越高,光合作用越强,形成的营养物质更多[37],且生长季末期的高温在一定程度上延长了生长季[29],有利于营养物质的积累,促进来年树木的生长。
3.3 2个树种径向生长对降水响应的差异
丽江云杉与上年8月和当年5月降水呈显著负相关,而高山松与各月降水均未达到显著相关水平,这主要与丽江云杉和高山松的根系特性有关。8月降水较多,且经过前期降水的积累(图1),土壤含水量过于饱和,会造成树木根部的无氧呼吸,消耗过多的光合产物,进而不利于树木来年生长[38- 39]。而5月温度快速上升(图1),冰雪大量融化,若这一时期降水增加,容易导致林中积水,对树木根系产生不利影响而抑制树木的生长[39]。丽江云杉为浅根性树种,对土壤含水率变化更为敏感,因此表现出与降水的显著相关关系。而高山松的根系较深,受到的影响相对较小,且其主要生长在阳坡、半阳坡,土壤水分蒸发量较大,减小了土壤含水量饱和对其生长的抑制作用。
3.4 冗余分析
丽江云杉和高山松年表与气候因子之间的冗余分析与传统的响应函数分析结果基本一致,上年11月、10月、当年7月温度以及上年8月降水是影响该地区树木生长的主要气候因子,进一步证明了该方法能够有效量化树木径向生长与气候因子的关系。冗余分析与响应函数分析之间也存在一定差异,比如响应函数分析还检测到当年5月降水对丽江云杉径向生长的影响,而冗余分析检测到的是上年7月和8月温度对树木生长的影响,这种与响应函数方法的差异在前人研究中同样有所发现[40],两者的综合应用可以更好的揭示影响树木生长的主要气候因子。
综上,石卡雪山2个树种径向生长对气候响应存在差异,丽江云杉受温度和降水共同作用,高山松仅与温度表现出显著相关性,这与其生长环境和生物学习性差异有关。另外,气候因子对树木生长的滞后效应在该区域表现明显。本研究的结果有助于阐明影响石卡雪山针叶树种径向生长的主要气候因子,为其他地区不同树种径向生长对气候响应差异的研究提供参考。
参考文献(References):
[1] 王叶, 延晓冬. 全球气候变化对中国森林生态系统的影响. 大气科学, 2006, 30(5): 1009- 1018.
[2] 于大炮, 周莉, 代力民, 王庆礼, 刘明国. 树木年轮分析在全球变化研究中的应用. 生态学杂志, 2003, 22(6): 91- 96.
[3] Zhang Q B, Hebda R J. Variation in radial growth patterns ofPseudotsugamenziesiion the central coast of British Columbia, Canada. Canadian Journal of Forest Research, 2004, 34(9): 1946- 1954.
[4] 刘敏, 毛子军, 厉悦, 孙涛, 李兴欢, 黄唯, 刘瑞鹏, 李元昊. 不同纬度阔叶红松林红松径向生长对气候因子的响应. 应用生态学报, 2016, 27(5): 1341- 1352.
[5] Fritts H C. Tree Rings and Climate. London: Academic Press, 1976.
[6] 李广起, 白帆, 桑卫国. 长白山红松和鱼鳞云杉在分布上限的径向生长对气候变暖的不同响应. 植物生态学报, 2011, 35(5): 500- 511.
[7] 徐宁, 王晓春, 张远东, 刘世荣. 川西米亚罗林区不同海拔岷江冷杉生长对气候变化的响应. 生态学报, 2013, 33(12): 3742- 3751.
[8] 袁玉江, 李江风. 天山西部树轮年表的响应函数. 冰川冻土, 1995, 17(2): 170- 177.
[9] 张文涛, 江源, 王明昌, 张凌楠, 董满宇. 芦芽山阳坡不同海拔华北落叶松径向生长对气候变化的响应. 生态学报, 2015, 35(19): 6481- 6488.
[10] 张卫国, 肖德荣, 田昆, 陈广磊, 和荣华, 张贇. 玉龙雪山3个针叶树种在海拔上限的径向生长及气候响应. 生态学报, 2017, 37(11): 3796- 3804.
[11] 牛贺文, 何元庆. 玉龙雪山地区大气降水中粉尘颗粒物特征研究. 冰川冻土, 2014, 36(1): 71- 79.
[12] 杨文云, 李昆. 云南森林树种种质资源保存策略初探. 林业科学研究, 2002, 15(6): 706- 711.
[13] Fan Z X, Bräuning A, Cao K F. Annual temperature reconstruction in the central Hengduan Mountains, China, as deduced from tree rings. Dendrochronologia, 2008, 26(2): 97- 107.
[14] Bi Y F, Xu J C, Gebrekirstos A, Guo L, Zhao M X, Liang E Y, Yang X F. Assessing drought variability since 1650 AD from tree-rings on the Jade Dragon Snow Mountain, Southwest China. International Journal of Climatology, 2015, 35(14): 4057- 4065.
[15] Guo G A, Li Z S, Zhang Q B, Ma K P, Mu C L. Dendroclimatological studies ofPicealikiangensisandTsugadumosain Lijiang, China. IAWA Journal, 2009, 30(4): 435- 441.
[16] 余佳霖, 张卫国, 田昆, 松卫红, 李秋平, 杨荣, 张贇. 普达措国家公园海拔上限3个针叶树种径向生长对气候变化的响应. 北京林业大学学报, 2017, 39(1): 43- 51.
[17] 云南植被编写组. 云南植被. 北京: 科学出版社, 1987.
[18] 刘中天. 滇西北高山针叶林区高山松的发展趋势. 林业资源管理, 1981, (1): 32- 34.
[19] Stokes M A, Smiley T L. An Introduction to Tree-Ring Dating. Chicago: University of Chicago Press, 1968.
[20] Larsson L A. CDendro v. 7.3. Cybis Elektronik & Data AB. Sweden: Saltsjöbaden, 2010.
[21] Holmes R L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin, 1983, 43(1): 69- 78.
[22] Cook E R, Holmes R L. Users Manual for Program ARSTAN. Tucson: Laboratory of Tree-Ring Research, University of Arizona, 1986.
[23] Kendall M G, Gibbons J D. Rank Correlation Methods. 5th ed. London: Edward Arnold, 1990.
[24] Biondi F, Waikul K. DENDROCLIM2002: a C++ program for statistical calibration of climate signals in tree-ring chronologies. Computers & Geosciences, 2004, 30(3): 303- 311.
[25] ter Braak C J F. Canonical community ordination. Part I: basic theory and linear methods. Écoscience, 1994, 1(2): 127- 140.
[26] ter Braak C J F, Smilauer P. CANOCO Reference Manual and CanoDraw for Windows User′s Guide: Software for Canonical Community Ordination. Version 4.5. Ithaca, New York: Micro-computer Power, 2002.
[27] 李江风, 袁玉江, 由希尧. 树木年轮水文学研究与应用. 北京: 科学出版社, 2000.
[28] 赵志江, 谭留夷, 康东伟, 刘琪璟, 刘俊清. 云南小中甸地区丽江云杉径向生长对气候变化的响应. 应用生态学报, 2012, 23(3): 603- 609.
[29] 彭剑锋, 勾晓华, 陈发虎, 刘普幸, 张永, 方克艳. 阿尼玛卿山地不同海拔青海云杉(Piceacrassifolia)树轮生长特性及其对气候的响应. 生态学报, 2007, 27(8): 3268- 3276.
[30] Fritts H C, Dean J S. Dendrochronological modeling of the effects of climatic change on tree-ring width chronologies from the Chaco Canyon area, southwestern United States. Tree-Ring Bulletin, 1992, 52: 31- 58.
[31] 李宗善, 刘国华, 张齐兵, 胡婵娟, 罗淑政, 刘兴良, 何飞. 利用树木年轮宽度资料重建川西卧龙地区过去159年夏季温度的变化. 植物生态学报, 2010, 34(6): 628- 641.
[32] Szeicz J M, MacDonald G M. A 930-year ring- width chronology from moisture- sensitive white spruce (PiceaglaucaMoench) in northwestern Canada. The Holocene, 1996, 6(3): 345- 351.
[33] 肖丁木, 秦宁生, 李金健, 李媛媛. 树轮宽度指示的川西色达7月平均最高气温变化. 中国沙漠, 2013, 33(5): 1536- 1543.
[34] 喻树龙, 袁玉江, 魏文寿, 张同文, 尚华明, 陈峰. 川西马尔康7月平均气温的重建及其气候周期信号检测. 高原气象, 2012, 31(1): 193- 200.
[35] 彭剑锋, 勾晓华, 陈发虎, 张永香, 刘普幸, 张永, 方克艳. 阿尼玛卿山地祁连圆柏径向生长对气候的响应. 地理学报, 2007, 62(7): 742- 752.
[36] 赵志江, 康东伟, 李俊清. 川西亚高山不同年龄紫果云杉径向生长对气候因子的响应. 生态学报, 2016, 36(1): 173- 179.
[37] 陈力, 尹云鹤, 赵东升, 苑全治, 吴绍洪. 长白山不同海拔树木生长对气候变化的响应差异. 生态学报, 2014, 34(6): 1568- 1574.
[38] 李腾, 何兴元, 陈振举. 东北南部蒙古栎径向生长对气候变化的响应——以千山为例. 应用生态学报, 2014, 25(7): 1841- 1848.
[39] 高露双, 王晓明, 赵秀海. 长白山过渡带红松和鱼鳞云杉径向生长对气候因子的响应. 植物生态学报, 2011, 35(1): 27- 34.
[40] 王晓春, 宋来萍, 张远东. 大兴安岭北部樟子松树木生长与气候因子的关系. 植物生态学报, 2011, 35(3): 294- 302.
猜你喜欢
世界建筑(2022年10期)2022-10-25
景德镇陶瓷(2021年1期)2021-03-24
现代农村科技(2020年7期)2020-12-19
艺术品鉴(2019年12期)2020-01-18
乡村科技(2019年14期)2019-02-21
启迪与智慧·教育版(2018年12期)2018-02-26
江苏农业科学(2017年5期)2017-04-15
甘肃林业(2016年2期)2016-11-07
科技与创新(2016年18期)2016-11-04
湖北农业科学(2014年21期)2014-12-22
