
镉氮对台湾桤木和杂交相思生长与光合特性的影响
2018-05-08 03:04:50谭长强郝海坤彭玉华申文辉黄志玲曹艳云
西北农林科技大学学报(自然科学版) 2018年2期
谭长强,郝海坤,彭玉华,申文辉,黄志玲,曹艳云
(1广西林业科学研究院,广西 南宁530002;2广西优良用材林资源培育重点实验室,广西 南宁530002;3国家林业局中南速生材繁育实验室,广西 南宁530002)
镉在土壤中的高度移动性和对植物的严重危害,被视为最具危害性的重金属污染元素之一[1]。镉通过食物链进入人体,对人的健康构成威胁。镉严重抑制植物的生长以及光合作用[2],但抑制程度与镉处理的时间和浓度以及作物种类、生育时期相关。氮是植物生长发育中的重要营养元素,施氮有利于植物对镉胁迫环境的适应[3-4]。
木本植物,特别是生长迅速、生物量大的木本植物,已经显示出用于植物土壤修复的应用价值[5-6]。但目前有关氮肥对木本植物镉胁迫适应性的研究甚少,仅见张帆等[7]研究了施氮对镉胁迫下杨树(Populus)的影响,得出施氮可缓解镉对杨树的毒害,并提高了杨树叶绿素含量和光合速率。已有研究表明,菌根能够促进植物对氮、磷以及其他营养物质和水分的吸收,从而增强植物抵抗环境压力的能力[8]。因此,本试验选择生长迅速,并且均具有固氮菌根的台湾桤木(Alnusformosana)及杂交相思(Acaciamangium×Acaciaauriculiformis)作为试验对象,研究2个树种对镉胁迫环境的适应性及施氮对2个树种解毒机制的影响,为土壤环境镉污染的治理提供更好的途径和方案。
1 材料与方法
1.1 试验材料
台湾桤木是桦木科桤木属(Alnus)树种,原产于台湾地区,在我国福建、四川、广西等地均有种植,有望成为长江以南地区极具潜力的阔叶林树种[9]。杂交相思为马占相思与大叶相思的杂交种,系含羞草科金合欢属(Acacia)速生乔木树种,是南方重要的短周期工业原料林树种[10]。2 树种均具有结瘤固氮的根瘤,并且适应力强,耐贫瘠,是不可多得的速生和兼具改良土壤的树种。试验苗木为扦插0.5 年生营养袋苗,均由广西林业科学研究院苗圃培育。
1.2 试验地及供试土壤
本试验在广西林业科学研究院苗圃进行,属湿润的亚热带季风气候,全年降水天数在160 d左右,具有明显的干湿季节,年平均降雨量1 650 mm,主要集中在5-9 月[11]。供试土壤为红壤,土壤pH值为4.73,土壤有机质、全N、全P、全K含量分别为17.31,0.542,0.344,7.08 g/kg,土壤水解N、速效P、速效K含量分别为66.73,1.26,98.14 mg/kg。
1.3 试验设计
试验采用直径28 cm,高28 cm的塑料盆,每盆装土10 kg。试验设置6个处理(土壤氮肥选用尿素[CO(NH2)2],Cd2+选用CdCl2·2.5H2O),各处理最终土壤施入量分别为:对照(CK),0 mg/kg Cd2+;D1,30 mg/kg Cd2+;D2,60 mg/kg Cd2+;D3,120 mg/kg Cd2+;D1N1,30 mg/kg Cd2++0.4 g/kg CO(NH2)2;D1N2,30 mg/kg Cd2++0.8 g/kg CO(NH2)2,每处理20株。Cd2+以CdCl2·2.5H2O(分析纯)水溶液的形式一次性加入,与土壤混合均匀并在遮雨棚下均衡7 d;氮肥均等分6次施入,其中第1次于2016-03-01施入,以后每15 d施1次。于2016-02-20取预先培养好的成活植株(苗高、地径基本一致,分别约为45.6 cm和 3.25 mm),每盆栽植1株,试验期间保持土壤湿润,并在盆下放置塑料托盘,浇水后将渗出到盘内的水分倒回至盆中,以免镉元素流失。按照常规方法进行松土和除草等处理。
1.4 测定指标及方法
培养90 d后,于5月20日采用LI-6400(LI-COR,USA)便携式光合仪测定光合作用参数,测定时间为上午09:00-11:00,光强、叶温、空气流量、CO2浓度分别设为1 200 μmol/(m2·s)、30 ℃、500 μmol/s和400 μmol/mol,每树种每处理随机抽取6株,每株取完全展开的成熟叶1片测定净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)。处理前1 d以及处理后每隔15 d测量所有苗的苗高、地径。试验结束后,于6月10日,每树种每处理选取6株平均苗木收获植株,水洗除去根部土壤后,105 ℃杀青2 h,80 ℃恒温烘干至恒质量,计算苗木平均生物量。
1.5 数据处理与分析
试验数据运用Excel 2003及DPS 7.0软件进行处理与分析,多重比较采用Duncan新复极差法,运用Origin软件进行制图。
2 结果与分析
2.1 镉、氮处理对台湾桤木及杂交相思苗高和地径增量的影响
由图1可知,台湾桤木及杂交相思苗高、地径增量均随着镉处理添加量的增加呈下降趋势。在镉添加量为30 mg/kg时,台湾桤木苗高、地径较CK分别显著下降了32.64%和47.72%,而杂交相思则分别显著下降了64.81%和52.04%。当镉添加量达到120 mg/kg时,台湾桤木及杂交相思苗高较CK下降幅度分别为61.51%和72.24%,地径下降幅度分别为58.84%和57.94%。
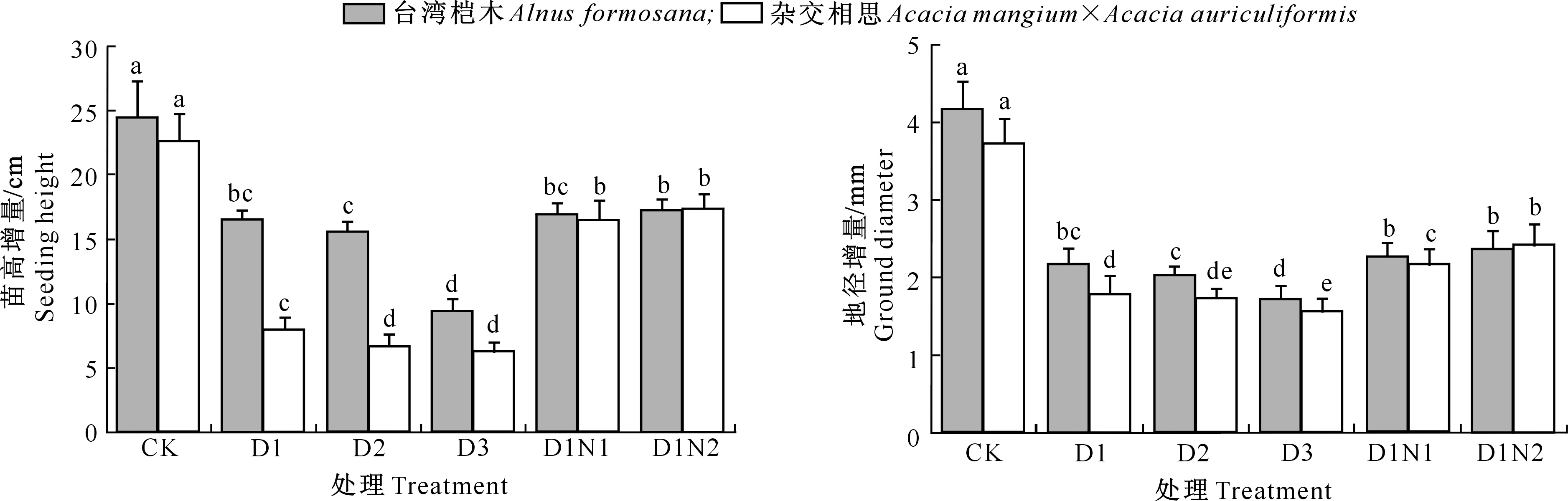
图柱上小写字母表示相同树种不同处理之间差异显著(P<0.05)。下图同 Values followed by different letters are significantly different at the 0.05 level under the same tree species among different treatments.The same below
在30 mg/kg镉胁迫下,随着施氮量的增加,2个树种的苗高均表现出增加的趋势。在高氮处理(D1N2)下,台湾桤木及杂交相思苗高增量比D1分别增加4.86%和118.08%,但台湾桤木D1N1、D1N2与D1之间并未达到显著水平;与D1处理相比,施氮(D1N1、D1N2)显著促进了杂交相思苗高的生长,但高氮(D1N2)与低氮(D1N1)处理之间也未达到显著差异水平。
在30 mg/kg镉处理下,台湾桤木及杂交相思地径随着施氮量的增加而不断上升,并且对杂交相思的影响各处理间达到了显著差异水平,而对台湾桤木的影响并未达到显著差异水平。在施高氮处理(D1N2)下,台湾桤木和杂交相思地径增量分别比D1处理增加了8.71%和35.63%。
2.2 镉、氮处理对台湾桤木及杂交相思幼苗生物量的影响
由表1可知,台湾桤木及杂交相思苗木根、茎、叶生物量均随着镉添加量的增加呈下降趋势。在30 mg/kg镉处理时,台湾桤木及杂交相思苗木根生物量分别比CK下降了44.74%和56.65%,茎生物量分别下降了54.92%和65.73%,叶生物量分别下降了65.88%和62.02%,总生物量分别下降了56.49%和62.26%;在镉添加量达到120 mg/kg时,下降达到最大值,与CK相比均达到了显著差异水平,此时台湾桤木及杂交相思根生物量下降幅度分别为60.23%和64.80%,茎生物量下降幅度分别为67.34%和70.03%,叶生物量下降幅度分别为77.70%和74.54%,总生物量下降幅度分别为69.63%和71.86%。
在30 mg/kg镉处理下,施氮均在一定程度上促进了台湾桤木及杂交相思根、茎、叶生物量的增加,但根生物量表现为随着施氮量的增加呈现先上升后下降趋势。台湾桤木及杂交相思在低氮(D1N1)处理时根生物量较D1分别增加了17.46%和76.20%,并且与处理D1之间均存在显著差异,说明低氮更能促进台湾桤木及杂交相思根的生长。茎、叶及总生物则随着施氮量的增加而上升,其中台湾桤木及杂交相思处理D1N2与D1相比茎生物量分别增加了16.70%和109.68%,叶生物量分别增加了33.96%和74.20%,总生物量分别增加了17.46%和76.20%,并且2种树茎、叶、总生物量处理D1N2与D1之间均存在显著差异,说明对台湾桤木及杂交相思施氮有利于其茎、叶及总生物量的提高。
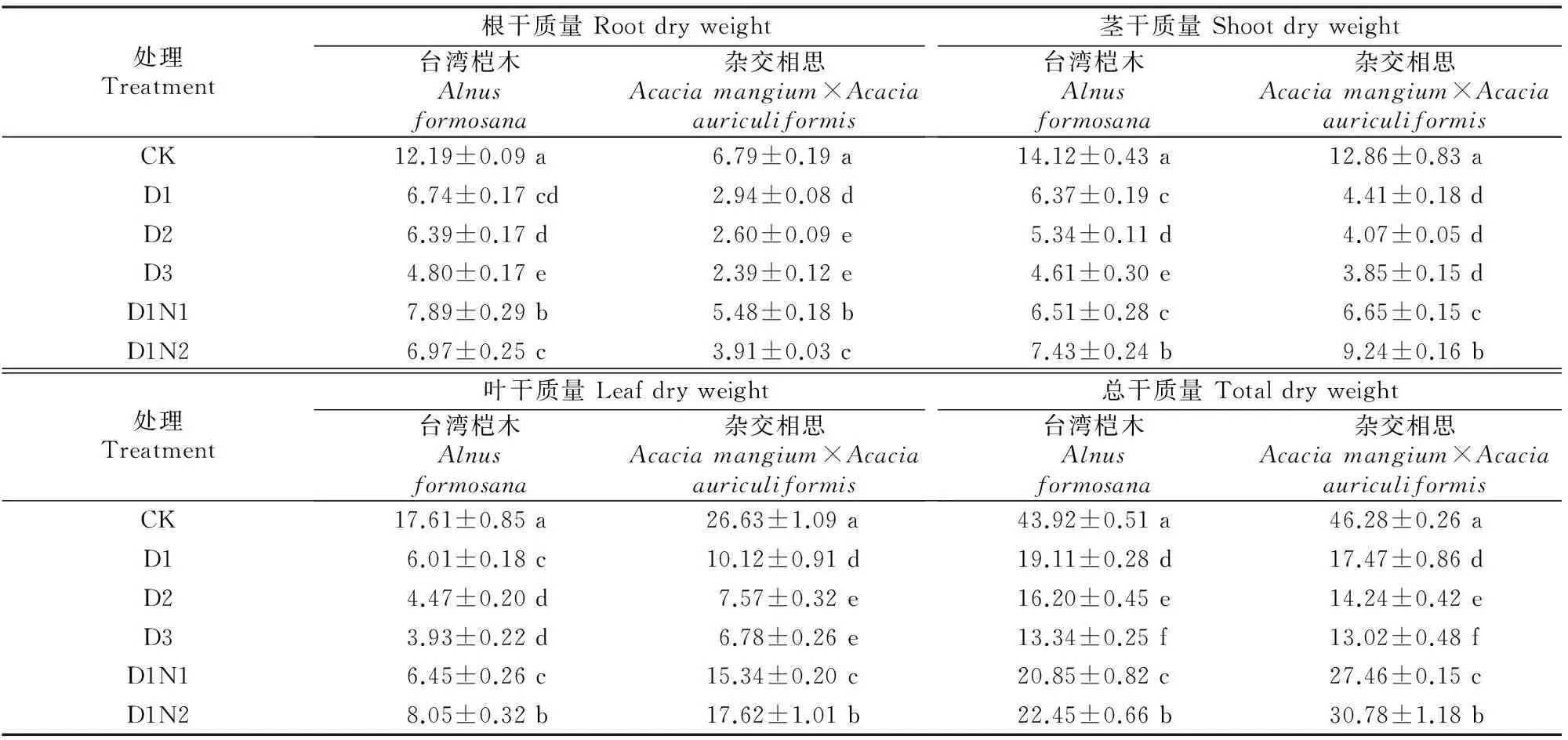
表1 不同镉、氮处理对台湾桤木及杂交相思生物量的影响Table 1 Effects of different Cd and N treatments on dry weight of Alnus formosana and Acacia mangium×Acacia auriculiformis g/株
注:同列数据后标不同小写字母表示不同处理间差异显著(P<0.05)。
Note:Values followed by different letters are significantly different at theP<0.05 level.
2.3 镉、氮对台湾桤木及杂交相思苗木光合特征的影响
2.3.1 净光合速率(Pn) 由图2可知,台湾桤木及杂交相思苗木叶片净光合速率(Pn)随着镉胁迫强度的增加均不断下降,在镉添加量达到30 mg/kg时,台湾桤木及杂交相思Pn下降幅度相差不大,分别为11.90%和12.12%;当镉添加量达到60 mg/kg时,杂交相思Pn下降了50.55%,而台湾桤木则仅下降了17.82%。
在30 mg/kg镉胁迫下,施氮对台湾桤木叶片Pn均有所提高,并且随着施氮量的增加呈现先升高后降低的趋势,但各处理之间并未达到显著差异水平。杂交相思叶片Pn则随着施氮量的增加而不断增加,并且处理D1N2与D1之间达到了显著差异水平。
2.3.2 气孔导度(Gs) 由图3可知,台湾桤木及杂交相思苗木叶片气孔导度(Gs)随着镉添加量的增加均呈下降趋势。在镉添加量达到30 mg/kg时,台湾桤木及杂交相思Gs分别较CK下降了23.22%和14.56%;当镉添加量达到120 mg/kg时,下降幅度分别为45.88%和48.23%,与对应CK相比均达到了显著差异水平。
在30 mg/kg镉胁迫下,台湾桤木和杂交相思Gs均随着施氮量的增加表现为不断上升的趋势,但台湾桤木施氮处理D1N1、D1N2与D1之间并无显著差异,而杂交相思各处理间均达到了显著差异水平。
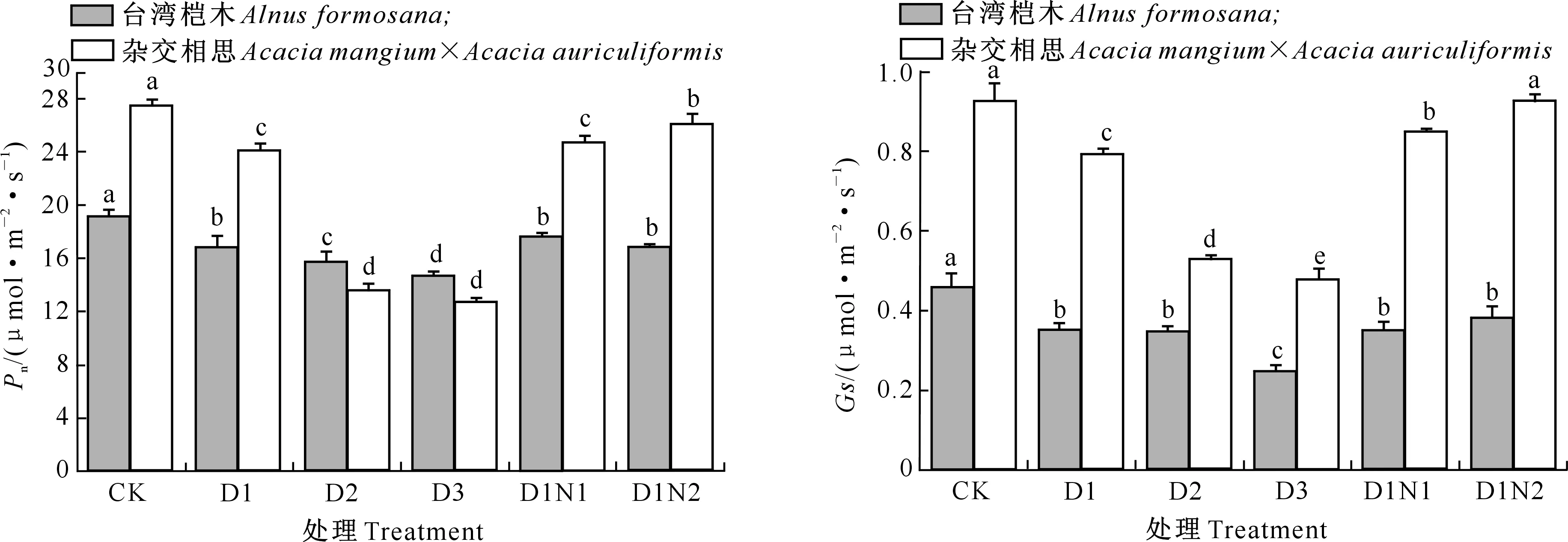
图2 不同镉、氮处理对台湾桤木及杂交相思 苗木净光合速率(Pn)的影响Fig.2 Effects of different Cd and N treatments on net photosynthetic rate (Pn) of Alnus formosana and Acacia mangium×Acacia auriculiformis
2.3.3 胞间CO2浓度(Ci) 由图4可知,台湾桤木及杂交相思苗木叶片胞间CO2浓度(Ci)随着镉处理质量浓度的增加表现为先下降后上升的趋势。在镉添加量为30 mg/kg时,台湾桤木Ci比CK下降了38.13%,而杂交相思仅下降了0.78%;当镉添加量达到60 mg/kg时Ci最低,分别比CK降低40.16%和34.98%,并且与CK相比均达到了显著差异水平;当镉添加量达到120 mg/kg时,Ci值均显著提升,分别仅比CK降低7.92%和9.74%。
在30 mg/kg镉胁迫下,施氮对台湾桤木和杂交相思Ci影响均呈现先增加后降低的趋势。台湾桤木处理D1N1、D1N2与D1之间均达到了显著差异水平,而处理D1N1与D1N2之间无显著差异。杂交相思处理D1N2与D1之间达到显著差异水平,而D1N1与D1之间无显著差异。
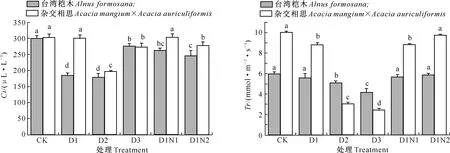
图4 不同镉、氮处理对台湾桤木及杂交相思 苗木胞间CO2浓度(Ci)的影响Fig.4 Effects of different Cd and N treatments on intercellular CO2 concentration (Ci) of Alnus formosana and Acacia mangium×Acacia auriculiformis
2.3.4 蒸腾速率(Tr) 由图5可知,台湾桤木及杂交相思苗木叶片蒸腾速率(Tr)随着镉添加量的增加均呈下降趋势。在30 mg/kg镉胁迫时,台湾桤木及杂交相思Tr较CK下降幅度分别为6.65%和11.97%,与对应CK相比,杂交相思达到了显著差异水平,而台湾桤木并未达到显著差异水平;当镉添加量达到120 mg/kg时,Tr较CK下降幅度分别为30.21%和75.58%,与CK相比均达到了显著差异水平。
在30 mg/kg镉处理下,台湾桤木及杂交相思Tr随着供氮量的增加呈现不断上升趋势,处理D1N2与D1相比分别提升了4.85% 和11.06%,但台湾桤木D1N2与D1之间并未达到显著差异水平,而杂交相思D1N2与D1之间达到了显著差异水平。
3 讨 论
镉胁迫下,多种植物生长受到抑制[2]。本试验结果表明,镉胁迫下台湾桤木及杂交相思苗高、地径、根茎叶及总生物量均随着镉胁迫强度的增加呈下降趋势,但2树种下降趋势存在一定差异。镉胁迫对杂交相思苗高、地径抑制程度要高于台湾桤木。说明镉胁迫下台湾桤木适应性要强于杂交相思。
张帆等[7]研究表明,镉胁迫的同时加入氮可缓解镉胁迫对其生长的抑制。本研究结果表明,30 mg/kg镉胁迫下施氮,台湾桤木及杂交相思苗高、地径、生物量虽然均低于无镉胁迫处理,但较30 mg/kg镉处理均有所提高,并且杂交相思要比台湾桤木提高的更明显,而高氮(0.8 g/kg)会降低氮对台湾桤木及杂交相思根的促进作用。
光合作用是植物生长的基础。Mediavilla等[12]报道,Pn降低主要有两方面的原因:气孔限制和非气孔限制。Farquhar等[13]认为,Ci值的大小是评判气孔限制和非气孔限制的依据。Pn、Gs和Ci同时下降时,Pn的下降为气孔限制,这可能是由于镉胁迫影响了保卫细胞K、Ca的吸收和运输,增加了叶片气孔阻力[14];相反,如果叶片Pn值的降低伴随着Ci值的提高,说明光合作用的限制因素是非气孔限制,这可能是由于严重镉胁迫产生的大量活性氧积累使膜脂过氧化[15],或是镉离子影响了Mg离子的正常运输[16],从而影响了叶绿素的合成,也有可能是原初光化学反应受到了伤害[17-18]。本研究结果表明,不施氮的镉胁迫下台湾桤木及杂交相思Pn、Gs均随土壤镉胁迫强度的增加呈下降趋势,并且在0~60 mg/kg镉胁迫时Ci也下降,说明此时2树种的光合作用主要受气孔限制;而当镉胁迫强度达到120 mg/kg时,Ci突然升高,而Pn及Gs仍然持续下降,说明此时光合作用受到了非气孔因素的限制,镉已经严重影响到了细胞的正常生理代谢。
气孔导度(Gs)的下降一般与叶片水势的降低以及气孔保卫细胞失水有关,并且与植物组织水分状况相关[19]。本研究结果表明,台湾桤木与杂交相思叶片的蒸腾速率(Tr)与气孔导度(Gs)变化趋势相似,即随着镉胁迫强度的增加,Tr下降,说明水分因子可能是影响2树种叶片Gs下降的主要原因,也可能是由于镉引起了叶片脱落酸(ABA)水平增加而导致气孔关闭[20]。而在30 mg/kg镉胁迫下,施氮对2树种产生的影响存在一定差异,低氮(0.4 g/kg)更有利于台湾桤木Pn和Ci的提高,而高氮(0.8 g/kg)更好地促进了杂交相思Pn、Gs和Tr的提高。说明氮可在一定程度上缓解植物镉中毒现象。这与张帆等[7]的研究结果“镉胁迫的同时加入氮能明显提高杨树植株的碳同化能力和新陈代谢速率”相似。本试验未对台湾桤木和杂交相思进行多个镉胁迫梯度下施氮,因此是否其他镉胁迫强度下施氮也会对2个树种产生同样的影响,还有待进一步研究。
4 结 论
镉胁迫下台湾桤木较杂交相思具有更高的适应能力,而施氮对杂交相思适应性的提高则明显高于台湾桤木。因此,在治理土壤镉污染措施上,应针对不同树种采取不同的种植管理措施来提高其对镉污染抵抗的能力。
[参考文献]
[1] Costa G,Morel J L.Efficiency of h+-atpase activity on cadmium uptake by four cultivars of lettuce [J].Journal of Plant Nutrition,1994,17(17):627-637.
[2] 曹 玲,王庆成,崔东海.土壤镉污染对四种阔叶树苗木叶绿素荧光特性和生长的影响 [J].应用生态学报,2006,17(5):769-772.
Cao L,Wang Q C,Cui D H.Impact of soil cadmium contain ration on chlorophyll fluorescence characters and biomass accumulation of four broad-leaved tree species seedlings [J].Chinese Journal of Applied Ecology,2006,17(5):769-772.
[3] Fabian G,Dezsi-Devay M.Ecophysiological studies of the relationship between heavy metal toxicity and nitrogen nutrition in the early development stage of winter wheat [J].Acta Botanica Hungarica,1987,33(3):219-234.
[4] 曹 莹,段 萌,刘玉莲,等.镉胁迫下铵态氮对春小麦光合生理特性的影响 [J].生态环境学报,2011,20(2):359-363.
Cao Y,Duan M,Liu Y L,et al.Effects of NH4+-N on photosynthetic characteristics of spring wheat plant under cadmium [J].Ecology and Environmental Sciences,2011,20(2):359-363.
[5] Pulford I D,Watson C.Phytoremediation of heavy metal-contaminated land by trees:a review [J].Environment International,2003,29(4):529-540.
[6] Yoon J,Cao X,Zhou Q,et al.Accumulation of Pb,Cu,and Zn in native plants growing on a contaminated Florida site [J].Science of the Total Environment,2006,368(2):456-464.
[7] 张 帆,万雪琴,王长亮,等.镉胁迫下增施氮对杨树生长和光合特性的影响 [J].四川农业大学学报,2011,29(3):317-321.
Zhang F,Wan X Q,Wang C L,et al.Effects of nitrogen supplement on photosynthetic characteristic and growth rate of poplar plants under cadmium stress [J].Journal of Sichuan Agricultural University,2011,29(3):317-321.
[8] 童 琳,唐旭利,张 静,等.菌根真菌侵染对植物生物量累积的影响 [J].生态环境学报,2014,23(9):1520-1525.
Tong L,Tang X L,Zhang J,et al.Influence of mycorrhizal colonization on plant biomass accumulation [J].Ecology and Environmental Sciences,2014,23(9):1520-1525.
[9] 王金锡,吴宗兴,龙汉利,等.台湾桤木研究与引种 [M].北京:科学出版社,2013.
Wang J X,Wu Z X,Long H L,et al.Alnusformosanaresearch and introduction [M].Beijing:Science Press,2013.
[10] 王凌晖,吴国欣,施福军,等.不同造林密度对杂交相思生长的影响 [J].南京林业大学学报(自然科学版),2009,33(2):134-136.
Wang L H,Wu G X,Shi F J,et al.Effects of different afforestation density on growth ofAcaciamangium×Acaciaauriculiformis[J].Journal of Nanjing Forestry University(Natural Science Edition),2009,33(2):134-136.
[11] 广西土壤肥料工作站.广西土壤 [M].南宁:广西科学技术出版社,1991.
Soil and Fertilizer Station,Guangxi.Guangxi soil [M].Nanning:Guangxi Science and Technology Press,1991.
[12] Mediavilla S,Santiago H,Escudero A.Stomatal and mesophyll limitations to photosynthesis in one evergreen and one deciduous Mediterranean oak species [J].Photosynthetica,2002,40(4):553-559.
[13] Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis [J].Annual Review of Plant Physiology,1982,33(1):317-345.
[14] 顾颉刚.镉胁迫对杨树无性系植株生长发育及部分生理特性影响的研究 [D].天津:天津师范大学,2008.
Gu J G.The study on Cd effect of growth and some physiological characteristics ofPopulusclones [D].Tianjin:Tianjin Normal University,2008.
[15] 苏 玲,章永松,林咸永,等.维管植物的镉毒和耐性机制 [J].植物营养与肥料学报,2000,6(1):106-112.
Su L,Zhang Y S,Lin X Y,et al.Cadmium toxicity and tolerance in vascular plants [J].Plant Nutrition and Fertilizer Science,2000,6(1):106-112.
[16] 慈敦伟,姜 东,戴廷波,等.镉毒害对小麦幼苗光合及叶绿素荧光特性的影响 [J].麦类作物学报,2005,25(5):88-91.
Ci D W,Jiang D,Dai T B,et al.Effect of Cd toxicity on photosynthesis and chlorophyll fluorescence of wheat seedling [J].Journal of Triticeae Crops,2005,25(5):88-91.
[17] 刘劲松,石 辉,李秧秧.镉胁迫对黄瓜幼苗光合和叶绿素荧光特性的影响 [J].水土保持研究,2011(5):187-190,196.
Liu J S,Shi H,Li Y Y.Effects of Cd2+stress on photosynthesis and chlorophyll fluorescence characteristics of cucumber seedlings [J].Research of Soil and Water Conservation,2011(5):187-190,196.
[18] Malo D D,Schumacher T E,Doolittle J J.Long-term cultivation impacts on selected soil properties in the northern Glains Plains [J].Soil & Tillage Research,2005,81:277-291.
[19] Sun G,Zhu Z,Fang X,et al.Effect of cadmium on photosynthesis and chlorophyll fluorescence of pakchoi [J].Plant Nutrition and Fertilizer Science,2004,11(5):700-703.
[20] Poschenrieder C,Gunse B,Barceló J.Influence of cadmium on water relations, stomatal resistance,and abscisic acid content in expanding bean leaves [J].Plant Physiology,1989,90(4):1365-1371.
猜你喜欢
中国书法(2023年9期)2023-12-26 18:03:59
干旱地区农业研究(2022年1期)2022-01-28 04:51:30
今日农业(2020年19期)2020-12-14 14:16:52
装备制造技术(2019年12期)2019-12-25 03:06:52
科技创新与品牌(2019年12期)2019-02-06 04:03:29
中南林业科技大学学报(2016年1期)2016-12-20 08:09:31
广西林业科学(2016年3期)2016-03-16 05:43:36
中国铸造装备与技术(2015年5期)2015-12-10 10:23:33
中国科技信息(2015年2期)2015-11-16 08:18:32
华东师范大学学报(自然科学版)(2014年1期)2014-04-16 02:54:56
