
洪水径流对种子传播及种群更新的影响
——以黄土丘陵沟壑区旱柳为例
2018-05-05 08:44:48刘姝彤焦菊英吴多洋
水土保持研究 2018年1期
刘姝彤, 焦菊英, 胡 澍, 吴多洋, 邓 娜
(1.西北农林科技大学 林学院, 陕西 杨凌 712100; 2.西北农林科技大学 水土保持研究所, 陕西 杨凌 712100)
种子扩散是指种子远离母体,到达另一个生境的动态过程,是多数植物实现更新的主要途径[1],是种群在空间上扩展分布的重要基础[2]。种子传播和扩散所形成的空间格局从根本上影响幼苗分布与生长的潜在范围,并最终决定种群空间分布格局[3]。另外,种子扩散可以有效阻止种群的灭绝[4],有利于种群数量和分布区的拓展[5],增强种群的抗干扰能力,维持生态系统的物种多样性和景观多样性[6]。影响种子分布格局和幼苗建成的关键因素主要是种子的二次扩散,即通过风媒、水媒、动物媒介等进行的扩散。目前已有较多研究集中在特定植物的种子雨[7-8]和种子分布格局研究[9-10],动物媒介(如鸟类、啮齿类动物等)对种子扩散的影响[11-15],以及种子的风传散等方面[16-17]。另外,有学者对黄土丘陵沟壑区人工降雨条件下土壤侵蚀对种子迁移和流失的影响做了相关研究[18-19],Yu等[20]研究表示侵蚀性降雨会造成黄土丘陵沟壑区撂荒坡的种子流失、种子二次扩散和种子聚集。但仍然缺乏干旱与半干旱地区在自然降雨条件下水媒对种子扩散及种群更新影响的研究。
本课题组在安塞县坊塌小流域进行野外调查时发现,有两个相邻淤地坝的坝地内生长了高密度的旱柳(Salixmatsudana)幼树,而坝地在2013年之前一直作为农田耕种玉米,由于2013年暴雨洪水而导致撂荒。向当地居民了解时被告知,坝地所生长的旱柳并非人工种植,而是由于2013年暴雨洪水将种子冲至坝体前所致。究竟是否如此呢?为此,本文调查此处两个淤地坝控制小流域范围内旱柳成树及幼树的生长分布特征,旨在解释坝地突然生长大面积密集旱柳的现象,验证此现象是由于洪水促进种子扩散,还是坝地为幼苗生长提供了有利条件,亦或是二者共同作用下改变了种群幼苗分布格局。以期为干旱半干旱地区自然降雨所产生的地表径流对种子二次扩散的作用以及对植物种群更新格局影响的相关研究提供依据。
1 材料与方法
本研究区为延安市安塞县坊塌流域两个相邻的淤地坝控制小流域,地处109°14′42″—109°17′02″E,36°47′30″—36°49′02″N,面积4.98 km2。坊塌流域属于黄土丘陵沟壑区,中温带大陆性半干旱季风气候,年平均气温8.8℃,年平均降水量505.3 mm,降雨主要集中在每年7—9月且多暴雨,无霜期157 d左右。
本研究对象为两个淤地坝控制小流域内分布的旱柳成树和幼树。旱柳为杨柳科柳属落叶乔木,因常见于江边河畔等湿润地区,故又名河柳、江柳。于2016年7—8月对2个坝控流域内的旱柳进行野外实地调查与测量。其中位于西侧淤地坝称为1号淤地坝,东侧称为2号淤地坝(图1)。
(1) 对于淤地坝控制流域内成片生长的旱柳幼树,通过挖掘观察其根部有无串根生长现象,以此判定其为实生苗或萌生苗。并使用GPS记录其成片生长边界的坐标。每个坝地沿沟道方向等距离取3个点,分别代表坝前、坝中和坝后的幼树生长情况;并在每个点在垂直于淤地坝沟道方向布设3个5 m×5 m样方;淤地坝控制流域内其余地区成片生长的幼树,使用GPS对其边界进行定位并记录,视其生长面积取5 m×5 m样方(共15个,调查面积共3.77 hm2),调查幼树的基径、树高和生长密度。对于单株生长的幼树,记录坐标并测量树高、基径。
(2) 对淤地坝控制流域内成片生长的旱柳成树,使用GPS对成片旱柳林的边界进行定位并记录,并视其生长面积取10 m×10 m样方(共229个,调查面积共11.36 hm2),进行每木检尺,记录成树树高、胸径,样方内株数,计算生长密度。对于单株生长的成树,记录坐标并进行每木检尺,记录树高、胸径。
采用Excel 2013,SPSS 16.0统计分析旱柳幼树成树生长数据;运用ArcMap勾绘坝控流域内旱柳的空间分布,其余所有图均在Origin 8.1中完成。
2 结果与分析
2.1 旱柳成树生长与分布特征
旱柳成树生长多分布于流域沟道内。1号淤地坝控制流域内旱柳成树共948株,分布范围较广,几乎遍布淤地坝上游每条沟道,因此其分布的斑块形状也多为沿沟道分布的长条状;而下游少有大面积旱柳成树分布,仅有几处小面积成树生长;1号坝成树多为成林分布,生长密度不等,低至26.06株/hm2,高可达338.84株/hm2,但生长密度高的旱柳成树均分布于较深的沟谷内,较浅的支沟内成树生长密度不高于50株/hm2;胸径多在30~40 cm范围内;偶有单株分布于路边及靠近沟谷处的位置。2号淤地坝控制流域内旱柳成树少,共94株,多为零星分布,但也均分布于沟道内,胸径大多小于10 cm,无胸径大于30 cm的成树(图2)。1号淤地坝控制流域内成树平均树高和平均胸径均高于2号坝内成树,且存在极显著差异(p<0.0001)(图3)。
2.2 旱柳幼树的生长与分布特征
两个淤地坝控制流域内幼树多集中于坝地上,均为两年生实生苗,1,2号淤地坝坝地上的幼树生长面积分别为1.41 hm2和1.55 hm2,总体上1号淤地坝坝地上旱柳幼树生长密度和株高均高于2号坝,生长状况较好。

图1两淤地坝控制流域内旱柳生长分布
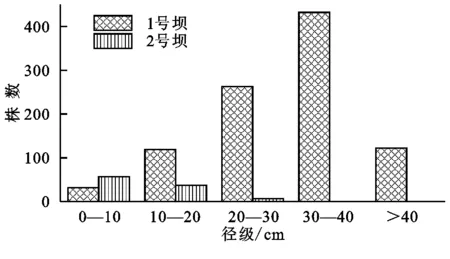
图2旱柳成树各径级数量分布

图3两淤地坝控制流域内旱柳成树生长特征比较
两个淤地坝密度最大处分别为坝后(12.88株/m2)和坝前处(6.77株/m2);偶有幼树分布于淤地坝内成树所分布的支沟沟口处,存在种子繁殖和无性繁殖两种繁殖方式,但无性繁殖较多。1号坝幼树生长密度从坝前至坝后总体呈现增大趋势且存在显著差异(p<0.0001),密度最大可达12.88株/m2;最低4.44株/m2。平均株高(p=0.009)及平均基径(p=0.016)均存在显著差异,且呈现减小趋势,其中株高最高达161.23 cm,最低为119.84 cm;基径则由1.38 cm递减到0.70 cm。2号坝则相反,生长密度由坝前至坝后存在显著差异(p=0.001),但呈减小趋势,生长密度最高处为6.80株/m2,远远小于1号坝生长密度;树高(p=0.004)及基径(p=0.005)均存在显著差异,且呈增加趋势,树高最高达143.07 cm,基径最大为1.62 cm(图4)。总体来看,1号坝坝地的旱柳幼树林生长更密集,株高也高于2号坝内植株株高,但因生长过密导致其基径较小。
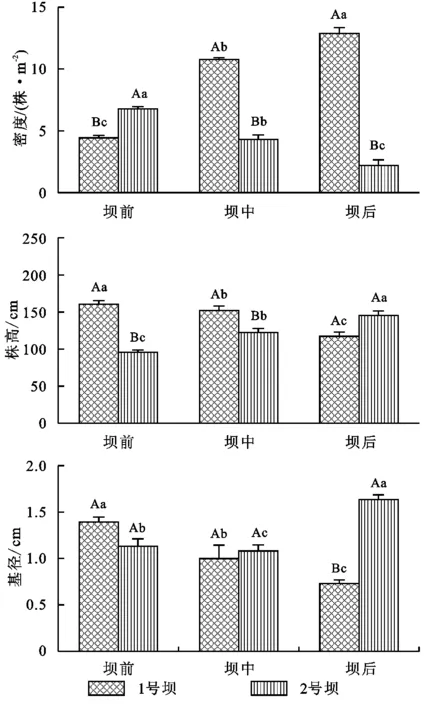
注:大写字母仅表示两淤地坝间差异,小写字母仅表示淤地坝内三个不同位置间差异。
图4两淤地坝坝地旱柳幼树生长特征
3 讨论与结论
3.1 旱柳的种子特性及生长习性
旱柳,又名河柳、江柳,为杨柳科柳属落叶乔木。柳属植物具备生长迅速(特别是种子萌发期和幼苗期)、光合速率高、根系对营养元素吸收和利用率高、喜光耐寒、喜湿耐旱、容易成活、抗性强等特点[21]。同样,旱柳幼苗生长过程中对土壤要求不严,生长速度快,萌芽力强,根系发达,极易成林,但在水分条件较好的土壤中生长较好,且多生长于海拔1 600 m以下的平原、山地溪流两侧,水分条件较好的沙丘及丘间低洼地带[22],东北、华北平原、西北黄土高原,西至甘肃、青海,南至淮河流域以及浙江、江苏均有分布,为平原地区常见树种[23],是中国温带地区分布广泛且较为常见的造林和绿化树种之一。因此淤地坝沟道及坝地此类水分条件良好的环境适宜其种子萌发及幼苗生长,旱柳花期早而长,为早春主要蜜源,其蒴果4—5月成熟,种子细小,基部具白色长毛,大多依靠风媒进行传播,可传播较远距离传播。繁殖能力较强,种子、扦插和埋条等方式均能完成繁殖,但扦插繁殖较易[24]。但有学者指出,种子植物如果长期进行扦插繁殖,会对种群整体的生存能力及遗传多样性造成影响[25];Rice等[26]也认为,植物通过种子繁殖及有性繁殖方式进行种群更新,可以显著提高其遗传多样性。
杨柳科植物种子成熟后没有休眠期,种子寿命短,通常被认为是“短寿种子”。通常认为寿命3 a或更短的为短寿种子,寿命3~15 a的为中寿种子,寿命15~100 a以上的为长寿种子[27]。“种子寿命”是指从种子完全成熟到种子丧失生活力所经历的时间[28]。但也有学者表示,通常被定义为短寿的种子,只在不适宜其存活的条件下突出表现为短命,当所处环境条件适宜时,种子寿命可延长[29]。目前并无具体研究表明旱柳种子寿命,但有研究指出同属植物蒙古柳(Salixlinearistipularis)种子寿命在常温条件下为45 d左右,在-80℃条件下其寿命可达10个月以上[30];高山柳(Salixcupularis)种子寿命为225 d左右[31]。因此推断旱柳种子在常温状态下可由种子成熟期(每年4—5月)存活至11月,而在经历冬季低温状态时,种子寿命可延长至次年春季。
3.2 洪水径流对旱柳种子二次扩散的作用
植物种子的扩散受种子数量[32]、种子特性、母树高度等植物自身的传播特性以及地形条件等因素的影响[17, 33]。在淤地坝此类特殊的立地条件下,由于自然降雨带来较大地表径流以及坝体对径流的拦截作用,水媒可对种子传播及种子分布格局产生重要影响。
旱柳种子细小,可通过风媒传播较远距离,而本研究区淤地坝控制流域内成树已生长多年,但坝地在2013年洪水之前由于耕种玉米并无旱柳的幼苗更新现象,2013年洪水之后耕地撂荒,2015年发现坝地成片生长旱柳幼树,且均为种子繁殖,因此推测由于2013年7—9月份安塞出现了7次大雨,3次暴雨,降雨总量达735.1 mm,造成土壤水分极度饱和,可能并不适宜种子萌发;2014年同时期出现了5次大雨,1次暴雨,降雨量相对减少(501.9 mm),但仍高于多年平均降雨量,仍然起到了对当年种子的扩散作用和淤积泥沙的作用,而2015年相对干旱(年降雨仅310.2 mm)[34],因此经过水分蒸发和入渗,2015年时土壤水分已降低了很多,达到适宜种子萌发的条件,致使了大量旱柳种子的萌发。可见,该淤地坝内或相邻区域旱柳成树产生的种子通过风媒传播至淤地坝控制流域范围内,再由洪水产生的地表径流进行了种子的二次传播,将整个淤地坝径流集水区内地表与土壤表层中存活的旱柳种子携带至淤地坝体前的坝地上;且地表径流被坝体拦截,导致径流携带泥沙最终都淤积在坝地上,对旱柳种子起到了聚集的作用,从而出现了旱柳幼树高密度生长的现象。
1号淤地坝上游存在较多较深、地势低洼的沟谷,形成了土壤水分条件良好的环境,适宜旱柳生长;因此1号淤地坝内旱柳成树数量多,生长好;而2号坝内沟谷较少,且沟谷多较浅,因此2号淤地坝内成树分布零星,生长状况不如前者。同时,1号淤地坝内旱柳幼树生长状况较好可能是由于1号坝内成树分布广,数量多,可产生种子数量多;且1号坝内成树较高,因此种子传播距离较远,但此区域内旱柳幼树并不完全来源于1号坝母树的种子扩散;而2号坝内虽然成树数量少,树高较低,导致种子数量少且传播距离有限,但由于风媒作用,可将相邻区域旱柳母树产生的种子传播至此,再由洪水将种子扩散至坝体前密集生长。
3.3 洪水径流对旱柳种子萌发与幼苗更新的影响
除传播和聚集种子的作用外,地表径流同时携带泥沙淤积在坝地内,改善了坝地的土壤水分条件和壤肥力状况。有研究指出淤地坝可改善农业生产条件,坝地地区的粮食生产量为梯田的2~3倍,坡耕地的4~6倍[35];胡建军等[36]的研究也发现淤地坝将荒沟变为高产稳产田。这都充分体现了淤地坝提高坝地土壤水分条件和肥力状况的作用。
杨柳科的植物种子没有休眠期,在适宜条件下即会萌发[37],且幼苗生长迅速。坊塌流域2013年7月的暴雨产生了大量洪水径流,2014年降雨量也高于多年平均值,致使2013年土壤水分极度饱和,2014年水分含量也较高;且降雨产生了较大的泥沙淤积量,而使研究区域两个淤地坝的坝地土壤肥力在原本就作为农田耕种施肥的基础上大大提高,在水分条件变得适宜时,便导致了淤地坝坝地旱柳种子萌发,幼苗快速生长的现象。
总而言之,从本研究观察到的现象表明,虽然自然降雨产生的地表径流会造成水土流失,但也会促进植物种子随之传播和扩散,并将种子携带至水分充足、适宜种子萌发和幼苗建成的生境,从而扩大植物种群分布,影响种群更新格局。可见,在干旱与半干旱地区,降雨径流在传播和扩散植物种子的同时,携带泥沙淤积于地表,提高了土壤含水量和土壤质量,恰巧为种子提供了充足的水分和肥力,创造了适宜种子萌发的生境,从而对植物种群更新起到了促进作用。因此,干旱与半干旱地区地表径流对种子的二次传播及其对植被更新的影响值得进一步加强研究。
参考文献:
[1] 常罡,邰发道.季节变化对锐齿栎种子扩散的影响[J].生态学杂志,2010,30(1):189-192.
[2] Russell S K, Schupp E W. Effects of microhabitat patchiness on patterns of seed dispersal and seed predation ofCercocarpusledifolius(Rosaceae)[J]. Oikos, 1998,81:434-443.
[3] Nathan R, Muller-Landau H C. Spatial patterns of seed dispersal, their determinants and consequences for recruitment[J]. Trends in Ecology & Evolution, 2000,15(7):278-285.
[4] 朱丽,卢剑波,余林.复合种群中扩散的研究进展[J].生态学杂志,2010,29(5):1008-1013.
[5] 龙翠玲,余世孝.茂兰喀斯特森林林隙种子雨、种子库空间变异[J].植物分类与资源学报,2007,29(3):327-332.
[6] 唐勇,曹敏,张建侯,等.西双版纳白背桐次生林土壤种子库、种子雨研究[J].植物生态学报,1998,22(6):505-512.
[7] 杜彦君,马克平.森林种子雨研究进展与展望[J].生物多样性,2012,20(1):94-107.
[8] 张健,郝占庆,李步杭,等.长白山阔叶红松(Pinuskoraiensis)林种子雨组成及其季节动态[J].生态学报,2008,28(6):2445-2454.
[9] 于飞.秦岭松栎混交林建群种种子扩散过程及天然更新格局研究[D].陕西杨凌:西北农林科技大学,2014.
[10] 张亚娟.黄土高原马栏林区落叶阔叶林种子雨动态及主要物种幼苗空间格局变化[D].西安:陕西师范大学,2015.
[11] 李宏俊,张知彬.动物与植物种子更新的关系Ⅱ.动物对种子的捕食、扩散、贮藏及与幼苗建成的关系[J].生物多样性,2001,9(1):25-37.
[12] 马庆亮,赵雪峰,孙明洋,等.啮齿动物作用下退耕地山杏种子扩散与贮藏的季节变化[J].应用生态学报,2010,21(5):1238-1243.
[13] 饶晓东,陈德祥,王代平,等.鸟类与植物种子扩散关系的研究进展[J].安徽农业科学,2010,38(26):14209-14212.
[14] 姜明敏,曹林,肖治术,等.都江堰林区取食樱桃果实(种子)的鸟类及其种子扩散作用[J].动物学杂志,2010,45(1):27-34.
[15] 曹林,郭聪.啮齿动物对印度栲种子扩散的效率[J].兽类学报,2011,31(4):323-329.
[16] 陈玲玲,林振山,何亮.风传草本植物种子空间传播新模型[J].生态学报,2010,30(17):4643-4651.
[17] 诸葛晓龙,朱敏,季璐,等.入侵杂草小飞蓬和钻形紫菀种子风传扩散生物学特性研究[J].农业环境科学学报,2011,30(10):1978-1984.
[18] 雷东,韩鲁艳,焦菊英,等.模拟降雨条件下黄土丘陵区主要植物种子的流失特征[J].水土保持研究,2011,18(1):1-6.
[19] Han L Y, Jiao J J, Wang N, et al. Seed removal on loess slopes in relation to runoff and sediment yield[J]. Catena, 2011,85:12-21.
[20] Yu W J, Jiao J J, Chen Y, et al. Seed removal due to overland flow on abandoned slops in the chinese hilly-gullied loess plateau region[J]. Land Degradation & Development, 2017,28:274-282.
[21] 隋德宗.盐胁迫对柳树无性系幼苗生长影响的研究[D].南京:南京林业大学,2006.
[22] 华北树木志编写组.华北树木志[M].北京:中国林业出版社,1984.
[23] 中国科学院中国植物志编辑委员会.中国植物志[M].北京:科学出版社,1990.
[24] 中国科学院西北植物研究所.秦岭植物志[M].北京:科学出版社,1974.
[25] Tomimatsu H, Ohara M. Genetic diversity and local population structure of fragmented populations of Trillium camschatcense(Trilliaceae)[J]. Biological Conservation, 2003,109:249-258.
[26] Rice K J, Gordon D R, Hardison J L, et al. Phenotypic variation in seedlings of a “keystone” tree species(Quercusdouglasii):the interactive effects of acorn source and competitive environment[J]. Oecologia, 1993,96:537-547.
[27] 美国农业部要务局,李霆.美国木本植物种子手册[M].北京:中国林业出版社,1984.
[28] 赵国余.蔬菜种子学[M].北京:北京农业大学出版社,1989.
[29] 傅家瑞.种子生理[M].北京:农业出版社,1992.
[30] 冯爽.蒙古柳(Salixlinearistipularis)有性繁殖特性研究[D].哈尔滨:东北林业大学,2013.
[31] 余道平,向丽,杨楠,等.高山柳种子萌发特性[J].西北农业学报,2013,22(3):148-151.
[32] Dechaine J M, Burger J C, Burke J M. Ecological patterns and genetic analysis of post-dispersal seed predation in sunflower(Helianthusannuus)crop-wild hybrids[J]. Molecular Ecology, 2010,19:3477-3488.
[33] 王妍,卢琦,程立岩,等.呼伦贝尔沙地天然樟子松种子扩散机制研究[J].生态科学,2010,29(5):403-410.
[34] 曹斌挺.黄土丘陵沟壑区退耕坡面不同植物群落的土壤侵蚀特征[D].陕西杨凌:西北农林科技大学,2016.
[35] 康俊生.淤地坝在治理水土流失改善农业生产条件中的作用[M].北京:北京农业出版社,2013.
[36] 胡建军,牛萍,曹炜.浅谈黄河上中游地区水土保持淤地坝工程的作用[J].水资源与水工程学报,2002,13(2):28-31.
[37] 张小单.珍稀濒危植物钻天柳的种子萌发和组织培养特性研究[D].哈尔滨:东北林业大学,2015.
猜你喜欢
中国水土保持(2023年1期)2023-03-06 01:57:12
河北果树(2020年1期)2020-02-09 12:31:18
中国水土保持(2019年11期)2019-11-13 09:27:36
天津师范大学学报(自然科学版)(2019年4期)2019-09-17 00:59:10
山东林业科技(2018年4期)2018-09-01 08:15:20
作文周刊·小学四年级版(2018年12期)2018-04-10 12:07:56
现代园艺(2017年11期)2017-06-28 11:32:57
现代农业科技(2016年21期)2017-03-06 00:31:54
土壤与作物(2015年3期)2015-12-08 00:46:58
现代农业(2015年1期)2015-02-28 18:40:08
