
糙皮侧耳(Pleurotus ostreatus)615 对小麦秸秆木质纤维素降解效果的研究
2018-04-24 02:32:15刘红菊屯妮萨麦提赛伊迪张仕琦杨开伦
中国饲料 2018年5期
刘红菊, 屯妮萨·麦提赛伊迪, 张仕琦, 杨开伦
(新疆农业大学动物科学学院,新疆肉乳用草食动物营养重点实验室,新疆乌鲁木齐830052)
我国小麦种植面积大,每年小麦秸秆达到了1.4 亿吨,(高振华和邸明伟,2008;李坚,2008)。但大量的小麦秸秆以燃烧以及填埋的方式处理,造成环境严重污染。将小麦秸秆作为粗饲料饲喂反刍动物,不仅提高了小麦秸秆的有效利用率,还可以解决畜牧业饲草料紧缺的问题(Wei等,2009)。但是,小麦秸秆纤维含量高,纤维素达到了41.00%,半纤维素达到了32.51%,木质素达到了15.40%(Lv等,2016),作为反刍动物粗饲料,其适口性差,消化率、利用率低,导致其潜能未被开发出来(王炫清等,2017)。因此,如何降低小麦秸秆中木质纤维素含量,提高反刍动物对小麦秸秆的利用率具有重要意义。目前,小麦秸秆作为粗饲料的主要处理方法有物理法、化学法和微生物发酵法。物理处理时间短、无污染且可提高小麦秸秆适口性,但耗能较大,成本较高,不适于大规模处理(卢松,2010)。化学法处理时间短,秸秆利用率高,但成本高且易造成二次环境污染。微生物处理小麦秸秆是在秸秆中加入有益微生物,微生物在代谢过程中产生漆酶、锰过氧化物酶和木质素过氧化物酶,其相互作用,可以降低粗纤维的含量,从而可以提高其作为粗饲料的利用价值。研究显示,秸秆经食用真菌发酵后,对秸秆的木质纤维具有较好的降解能力,可有效降解木质素,保留纤维素并降低干物质损失率。张荣等(2015)研究表明,利用Trichoderma asperellum1285发酵麦秸10 d后,半纤维素和木质素的损失率分别为23.22%、18.01%。糙皮侧耳菌株是担子菌门下伞菌目侧耳科一种类,是我国广泛栽培的食用菌,以农作物秸秆(棉花秸秆、稻草、小麦秸秆等)为底物培养食用菌,采摘子实体后测得秸秆中纤维素、半纤维素以及木质素含量降低,氮含量增加。本试验首先液体培养糙皮侧耳(Pleurotus ostreatus)615 菌株,研究其产漆酶、锰过氧化物酶的能力;其次利用菌液固体发酵经1.00%CaO处理的小麦秸秆,测定发酵后小麦秸秆中粗蛋白质、纤维素、半纤维素及木质素的含量变化,探究糙皮侧耳615对小麦秸秆木质纤维素的降解效果,为丰富小麦秸秆微生物处理技术的研究与应用提供参考。
1 材料与方法
1.1 培养基 马铃薯葡萄糖培养基(PD)的配制:去皮马铃薯200.00 g切成薄片放入装有1.00 L去离子水的烧杯中煮沸30 min,用8层纱布将马铃薯淀粉液滤出,加入葡萄糖20.00 g、KH2PO43.00 g、MgSO4·7H2O 1.50 g、 维生素 B11.00 g 搅拌,溶解,将配制好的PD培养液分装到三角锥瓶中,塞棉塞,并包报纸2层,线绳扎紧,进行121℃高压灭菌20 min,冷却后测得pH为6.0,备用。
1.2 菌种来源 糙皮侧耳 (Pleurotus ostreatus)615菌株购于新疆农业科学院微生物研究所。
1.3 种子液的制备 将备用PD培养液分别倒入3个250 mL三角锥瓶中各100.00 mL,并接入糙皮侧耳615菌株1.00 g,放入水浴摇床恒温振荡器,温度设为25℃,转速为150 r/min,培养 5 d,每天观察菌球生长情况。制得种子液,放4℃冰箱,备用。
1.4 粗酶液的制备 将100.00 mL发酵培养基装入250 mL三角瓶中,121℃灭菌20 min,冷却后接入5 d菌龄的糙皮侧耳615菌株种子液10.00 mL,做3个重复,放入水浴摇床恒温振荡器中,温度设为25℃,转速为150 r/min,培养10 d。每天取培养液2.00 mL至离心管中,离心(4℃,6000 r/min)10 min,用移液枪吸取上清液于离心管中冷冻(-20℃),保存,直至取到第10天为止。该粗酶液用于测定漆酶、锰过氧化物酶1~10 d的酶活力变化。
1.5 漆酶活力测定 用ABTS的氧化来表示漆酶的活性。其1.50 mL反应体系为:吸取浓度为2.50 mmol/L的2,2-联氮-二 (3-乙基-苯并噻唑-6-磺酸)二铵盐(ABTS)500.00μL和浓度为0.10 mol/L的醋酸钠缓冲液 (pH=4.5)900.00μL,加入100.00μL粗酶液,启动反应,测定反应5 min内420 nm处吸光度的变化。以转化1.00μmol ABTS所需的酶量来表示1个酶活力单位(1.00μmol/min)。
1.6 锰过氧化物酶活力测定 用MnSO4的氧化来表示锰过氧化物酶的活性。其1.41 mL反应体系为:吸取浓度为1.43 mmol/L的硫酸锰溶液(即用浓度为 50.00 mmol/L,pH=4.5酸-乳酸钠盐缓冲液配置MnSO4溶液)1.35 mL和浓度为0.08 mol/L的H2O2溶液0.05 mL,加入0.05 mL粗酶液,启动反应,测定反应5 min内270 nm处吸光度的变化。每分钟使1.00 mmol/L的Mn2+转化为Mn3+所需的锰过氧化物酶量来表示1个酶活力单位(1.00 μmol/min)。
1.7 糙皮侧耳615菌株固体发酵小麦秸秆 称取100.00 g小麦秸秆原料(粉碎至40目)放入烧杯中,在烧杯中加入1.00%的CaO及100.00 mL去离子水,用玻璃棒搅拌均匀后静置24 h。将静置24 h的小麦秸秆装袋并放入121℃高压灭菌锅中灭菌20 min,灭菌结束后放在超净工作台中冷却,待接种。试验分为三组分别为空白组、对照组和试验组,空白组为未处理的小麦秸秆原样,对照组为经1.00%CaO处理静置24 h的小麦秸秆,试验组为经1.00%CaO处理静置24 h后接种10%糙皮侧耳615菌株种子液的小麦秸秆。
在超净工作台中利用糙皮侧耳615菌株种子液接种冷却后的小麦秸秆样品,接种量为10.00%,将接种后的小麦秸秆样品25℃发酵30 d,每日观察发酵情况。
1.8 糙皮侧耳615菌株发酵小麦秸秆30 d后饲料常规成分的测定 未处理的小麦秸秆原样、经1.00%CaO处理静置24 h的小麦秸秆、糙皮侧耳615菌株发酵小麦秸秆样品干物质的测定采用GB/T6435-2006饲料中水分和其他挥发性物质含量的测定方法进行测定。中性洗涤纤维(NDF)和酸性洗涤纤维(ADF)含量采用美国ANKOM纤维分析仪进行测定,并计算出纤维素、半纤维素、木质素含量及降解率,蛋白质采用快速定氮仪测定。
1.9 扫描电镜分析 将待扫描小麦秸秆样品用导电胶带固定在铜台上,喷金后用日立新型高分辨场发射扫描电镜SU8010扫描电子显微镜观察。扫描电镜分析小麦秸秆试样为未粉碎样。
1.10 数据计算及处理 试验结果均以“平均值±标准差”表示,试验数据采用SPSS18.0软件的单因素方差分析(One-way ANOVA)程序进行方差分析,各组间平均值的多重比较采用Duncan法进行。
2 结果与分析
2.1 糙皮侧耳615菌株种子液形态与显微观察糙皮侧耳615菌株液体培养5 d后出现小米粒大小的淡黄色小菌球。糙皮侧耳615菌株显微镜下菌丝的结构有横隔和分枝,有锁状联合,菌丝粗细较均匀,较细胞狭长(王茂成,2013)。
2.2 漆酶活力测定 由图1可知,漆酶活力从第1天~第4天不断上升,并在第4天达到最大值1428.00 U/L,漆酶活力从第5天开始下降,且在第10天下降到399.00 U/L。

图1 1~10 d漆酶活力变化
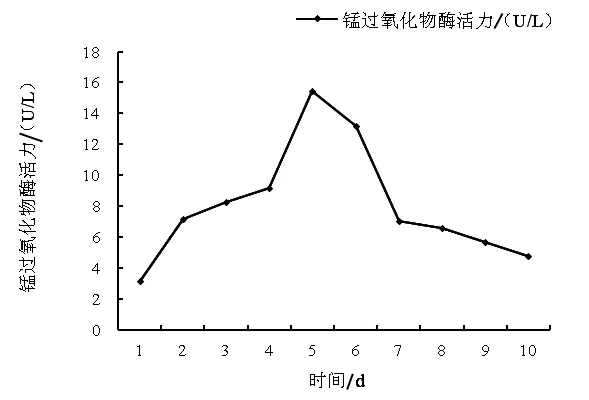
图2 1~10 d锰过氧化物酶活力变化
2.3 锰过氧化物酶活力测定 由图2可知,锰过氧化物酶活力从第1天至第5天不断上升,并在第5天达到最大值15.45 U/L,锰过氧化物酶活力从第6天开始下降,且在第10天下降到4.74 U/L。2.4 菌株发酵小麦秸秆30 d后饲料常规成分的含量 由表1可知,在中性洗涤纤维方面,试验组显著低于对照组和空白组(P<0.05),且分别低了4.23%和4.08,对照组与空白组无显著性差异(P>0.05);在酸性洗涤纤维方面,试验组显著高于对照组(P < 0.05),极显著高于空白组(P < 0.01),且分别高了4.19%和7.24%,对照组显著高于空白组(P<0.05),且高了2.93%;在纤维素方面,试验组极显著高于对照组与空白组 (P<0.01),且分别高了37.17%、38.87%,对照组与空白组无显著性差异(P>0.05);在半纤维素方面,试验组极显著低于对照组与空白组 (P<0.01),且分别降低了19.87%、20.89%,对照组与空白组无显著性差异;在木质素方面,试验组极显著低于对照组与空白组(P < 0.01),且分别降低了 43.75%、57.01%,对照组与空白组差异显著 (P<0.05),且比空白组低23.57%;在粗蛋白质方面,试验组极显著高于对照组与空白组 (P<0.01),且分别提高了4.14%、3.91%,对照组与空白组无显著性差异(P>0.05)。
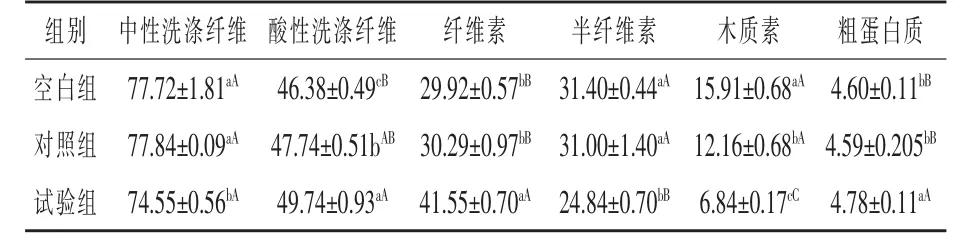
表1 糙皮侧耳615菌株发酵小麦秸秆30 d后对饲料常规成分含量的影响(干物质基础)%
2.5 小麦秸秆扫描电镜结构变化分析 小麦秸秆原样、1.00%CaO处理后小麦秸秆、糙皮侧耳615菌株发酵后小麦秸秆扫描电镜图分别见图3、图4,图5。图3所示在放大5000倍的情况下可以清楚地显示小麦秸秆原料表面光滑平整,结构致密有规则。图4所示经1.00%CaO处理后的小麦秸秆表面蜡质层遭到破坏,使得小麦秸秆表面出现裂痕。图5所示糙皮侧耳(Pleurotus ostreatus)615菌株发酵后小麦秸秆表面出现大量密集孔洞、细丝及膨松状态片状结构。

图3 小麦秸秆原样扫描电镜图(左:外侧 右:内侧5000×)

图4 1.00%CaO处理后小麦秸秆扫描电镜图(左:外侧 右:内侧5000×)

图5 糙皮侧耳(Pleurotus ostreatus)615菌株固态发酵30 d的小麦秸秆扫描电镜图(左:外侧 右:内侧 5000×)
3 讨论
3.1 糙皮侧耳615菌株液体培养1~10 d过程中漆酶、锰过氧化物酶活力的变化 漆酶和锰过氧化物酶是秸秆中粗纤维降解的关键酶,主要产生于真菌。在真菌菌丝生长过程中会分泌大量的漆酶与锰过氧化物酶。木质素的降解主要由漆酶与锰过氧化物酶参与完成。首先,木质素结构中的苯环在酶的催化作用下发生单电子氧化反应形成苯氧自由基;其次,木质素的降解产物被菌丝体吸收,进一步被氧化成二氧化碳和水,从而实现对木质素的降解。因此,提高漆酶与锰过氧化物酶的活力,是实现木质素降解的主要手段。本试验使用液体培养基培养糙皮侧耳615菌株,培养液中漆酶和锰过氧化物酶活性随着培养时间先升后降。漆酶在培养的第4天活性达到最高,活力为1428.00 U/L,此后漆酶的活力呈下降趋势,至第10天活力与培养初接近,与范文霞等(2008)以毛云芝菌为菌种进行液体培养(PD)的结果一致,其研究培养到第4天时漆酶的活力达到最高水平,为582 U/mL,且随着培养时间的延长,其酶活逐渐降低。本试验中漆酶活力比范文霞等试验结果中漆酶活力高59.24%,可能因为菌种不同其产酶能力也不同。
池玉杰等(2009)研究表明,在红平菇降解木质素试验中以木屑为产酶底物,测得锰过氧化物酶在第11天达到最高,之后随时间延长逐渐下降。而范鹏珍等(2011)研究也表明,在含Mn2+的LNAS培养基中不加底物条件下培养菌株可检测到含量较低的锰过氧化物酶,但产酶最佳时间为第13天,出现延后。本试验锰过氧化物酶在培养的第5天活性达到最高,活力为15.45 U/L,此后酶活开始降低。与池玉杰、范鹏珍等结果相比较,本试验锰过氧化物酶活性到达最高值的时间明显缩短,出现这种结果的原因可能与试验培养基、添加的底物以及菌种有关。池玉杰(2009)在培养菌种过程中以木屑为底物,可能因为菌种生长过程中产生锰过氧化物酶用来降解木屑中的粗纤维而被消耗,直至粗纤维降解结束,才使得锰过氧化物酶活性在第13天达到最大值。在未添加底物条件下,范鹏珍(2011)研究利用低氮天冬酰胺-琥珀酸培养基 (LNAS)培养杏鲍菇和乳白耙齿菌,LNAS培养基除了含有KH2PO4和MgSO4以外,还含有CaCl2等矿物质溶液,该培养基pH为4.5,本试验PD培养基主要含有马铃薯淀粉、葡糖糖、蔗糖、KH2PO4、MgSO4等,且 pH 为 6.0。 因此,培养基的成分不同、pH不同导致范鹏珍等研究产酶时间与本试验产酶时间不同。在本试验培养条件下,使用糙皮侧耳615菌株作为菌种,以PD作为产酶培养基可以缩短产生锰过氧化物酶的时间。研究表明,在偏碱性培养基中真菌产锰过氧化物酶的活力更高(李旭东等,2006)。本试验在PD培养基内添加维生素B1,维生素B1可提高培养基pH,可能是使锰过氧化物酶活力达最高值时间缩短的原因。
3.2 糙皮侧耳615菌株发酵小麦秸秆30 d后对其营养物质变化的影响 小麦秸秆粗纤维含量高(60%以上),粗蛋白质含量低(3% ~6%)(魏敏,2002)且适口性差,作为粗饲料不能被完全消化,利用率低。目前小麦秸秆作为粗饲料饲喂家畜的方式为直接饲喂或加工成颗粒饲喂,颗粒饲喂方法虽然提高了采食量,但利用率仍然很低(哈丽代·热合木江等,2014)。因此,小麦秸秆作为粗饲料的有效利用,一是要提高动物对小麦秸秆的采食量,二是要提高小麦秸秆中木质纤维素的消化率。
提高小麦秸秆中木质纤维素 (特别是木质素)的降解率是提高小麦秸秆消化利用的关键。木质纤维素的降解主要是通过多种纤维降解酶协同作用来实现的。研究显示,将真菌接种到棉花秸秆(Shi等,2008)、玉米秸秆(侯进等,2010)、稻草秸秆(李燕荣,2010)),其分泌的超纤维氧化酶可溶解秸秆表面的蜡质,并吸附在木质纤维素的端部,然后,菌丝由端部向内延伸,并产生纤维素酶、半纤维素酶、内切聚糖酶和外切聚糖酶将秸秆降解为小分子碳源和半纤维素,为菌丝的生长和木质素的降解提供碳源和能量(邓缘,2005;Hadar等,1992)。
张荣等 (2015)研究发现,T.asperellum1285菌株发酵麦秸8 d后纤维素、半纤维素分别降低了9.94%、18.58%。T.asperellum1285菌株发酵麦秸10 d后半纤维素和木质素的分别降低了23.22%、18.01%。本试验糙皮侧耳615菌株固态发酵小麦秸秆30 d后其中性洗涤纤维、半纤维素和木质素分别降低了4.23%、19.87%和43.75%,而小麦秸秆酸酸性洗涤纤维和纤维素含量分别升高了4.19%和37.17%。本试验半纤维素、木质素降解率明显高于张荣等(2015)研究结果,纤维素降解率低于其结果。可能原因是张荣等发酵麦秸时间与本试验不同,本试验发酵时间较张荣等发酵时间长,使得半纤维素、木质素被降解的彻底。侯进等(2010)应用白腐真菌发酵曲种发酵玉米秸秆结果显示,中性洗涤纤维含量升高,与本试验研究结果相同。由于菌种不同导致其对纤维素降解能力也不同,且小麦秸秆品种和收获时期等不同,中性洗涤纤维、酸性洗涤纤维含量有差异 (朱顺国等,2001)。Hadar等(1992)研究发现,平菇发酵棉籽壳三周时,降解木质素作用深入到密集组织中;随后,植物的木髓部分逐渐被分离出来,导管处的细胞也开始被降解,第四周发现木质素在体外的降解速率逐渐加快,含量明显降低,所以本试验中半纤维素及木质素比张荣等第8天、第10天的降解率高。此外,通过电镜扫描经糙皮侧耳615菌株发酵后的小麦秸秆,发现其表面覆盖的一层蜡质层遭到破坏,菌丝直接进入小麦秸秆内部分泌漆酶、锰过氧化物酶,这两种酶相互作用对内部纤维素、半纤维素及木质素进行降解,使得小麦秸秆被分解成膨松状态的片状结构。此结果进一步证明了糙皮侧耳615菌株可降低小麦秸秆中纤维含量。Arora(1995)和Asiegbu等(1996)研究表明,由于白腐真菌不同菌种的生理差异,具有不同的木质素降解酶系,对粗纤维的降解能力也不同。李野等(2008)研究发现,高效降解秸秆木质素的4株侧耳属食用菌类固体发酵小麦秸秆30 d后小麦秸秆中蛋白质含量显著增加了11.22%。本试验通过糙皮侧耳615菌株发酵小麦秸秆30 d后粗蛋白质增加了4.14%,蛋白质的增加量显著低于李野等研究结果。可能与处理所用的菌种、发酵条件等有关,因为菌种不同、发酵条件不同,发酵菌的生长量也不同。
4 结论
4.1 糙皮侧耳615菌株液体培养过程中,漆酶在第4天达到最大值1428.00 U/L,锰过氧化物酶第5天达到最大值15.45 U/L。
4.2 小麦秸秆经糙皮侧耳615菌株固体发酵30 d后,小麦秸秆中的木质素含量、半纤维素含量均显著降低,粗蛋白质含量显著升高。
[1]池玉杰,闫洪波.红平菇木质素降解酶系统漆酶、锰过氧化物酶及木质素过氧化物酶的检测[J].林业科学,2009,45(12):154 ~158.
[2]邓缘.浅谈秸秆木质素的生物降解[J].江西饲料,2005,5:29~31.
[3]范鹏珍,池玉杰,惠非琼,等.杏鲍菇和乳白耙齿菌锰过氧化物酶活性规律的研究[J].中国林副特产,2011,29(1):1 ~ 4.
[4]范文霞,蔡友华,刘学铭,等.毛云芝菌产漆酶液体培养条件的优化[J].食品与生物技术学报,2008,27(3):88 ~ 93.
[5]高振华,邸明伟.生物质材料及应用[M].北京:化学工业出版社,2008.
[6]哈丽代·热合木江,阿依古丽·达噶尔别克,扎依尔·艾山,等.粉碎或颗粒化棉花秸秆饲喂绵羊对增质量的影响[J].饲料研究,2014,13:52 ~ 53.
[7]侯进,李婷,李杰.白腐真菌降解玉米秸秆的研究[J].饲料博览,2010,1:4 ~ 7.
[8]侯进,李婷,吕文龙,等.应用白腐真菌发酵曲种发酵玉米秸秆的研究[J].中国奶牛,2010,7:25 ~ 27.
[9]李坚.生物质复合材料学[M].北京:科学出版社,2008.
[10]卢松.微生物处理玉米秸秆的腐解特征研究:[硕士学位论文][D].重庆:西南大学,2010.
[11]李旭东,荚荣,吕飒音,等.不同缓冲体系下pH值对裂褶菌F17产锰过氧化物酶的影响 [J].安徽大学学报 (自然科学版),2006,30(5):78 ~ 82.
[12]李燕荣.双孢蘑菇高效栽培及其对秸秆生物降解的研究:[硕士学位论文][D].湖南长沙:中南林业科技大学,2010.
[13]李野,梁军锋,张洪生,等.4株食用菌发酵小麦秸秆粗蛋白含量研究[J].饲料研究,2008,9:41 ~ 44.
[14]王炫清,苗嘉曦,陈兴,等.高效木质纤维素分解菌Aspergillus fumigatus Z5原位分解小麦秸秆的研究 [J].南京农业大学学报,2017,40(4):671 ~ 680.
[15]王茂成.木质素降解真菌的筛选鉴定及相关酶活性研究:[硕士学位论文][D].重庆:西南大学,2013.
[16]魏敏.棉花秸杆对绵羊饲用价值的初步研究:[硕士学位论文][D].乌鲁木齐:新疆农业大学,2002.
[17]张荣,奚永兰,叶小梅,等.棘孢木霉1285对麦秸的降解及厌氧发酵的影响[J].江苏农业学报,2015,31(2):328 ~ 333.
[18]朱顺国,邢壮,张微,等.玉米秸秆NDF与ADF含量变化规律的研究[J].中国奶牛,2001,1:24 ~ 26.
[19]Arora D S.Biodelignification of wheat straw by different fungal associations[J].Biodegradation,1995,6(1):57 ~ 60.
[20]Asiegbu F O,Paterson A,Smith J E.The effects of co-fungal cultures and supplementation with carbohydrate adjuncts on lignin biodegradation and substrate digestibility[J].World Journal of Microbiology&Biotechnology,1996,12(3):273 ~ 279.
[21]Hadar Y,Kerem Z,Gorodecki B,et al.Utilization of lignocellulosic waste by the edible mushroom,Pleurotus[J].Biodegradation,1992,3(2 ~ 3):189 ~ 205.
[22]Shi J,Chinn M S,Sharmashivappa R R.Microbial pretreatment of cotton stalks by solid state cultivation of Phanerochaete chrysosporium.[J].Bioresource Technology,2008,99(14):6556.
[23]Lv E,Xia W,Tang M,et al.Preparation of an efficient oil-spill adsorbent based on wheat straw[J].BioResources,2016,12(1):296 ~315.
[24]Wei Xiang W,Hao Hao L,Wei L,et al.Decomposition of Bacillusthuringiensis (Bt) transgenic criceresidues(straw and roots) in paddy fields[J].Journal of Soilsand Sediments,2009,10(9):457 ~ 467.
猜你喜欢
黑龙江大学自然科学学报(2022年4期)2022-11-17 08:07:52
安庆师范大学学报(自然科学版)(2022年3期)2022-09-20 07:10:38
河北科技师范学院学报(2022年2期)2022-08-26 08:55:46
江西农业大学学报(2020年4期)2021-01-27 01:06:02
上海包装(2019年8期)2019-11-11 12:16:14
食用菌(2019年4期)2019-08-27 06:35:00
天津造纸(2016年1期)2017-01-15 14:03:29
中国塑料(2016年3期)2016-06-15 20:30:00
西南农业学报(2016年5期)2016-05-17 05:42:45
中国造纸学报(2015年1期)2015-12-16 19:35:25
