
武夷山常绿阔叶林空间结构参数分布特征
2018-04-19 01:21陈婷婷陈水飞葛晓敏方炎明
生态学报 2018年5期
陈婷婷, 徐 辉, 杨 青, 陈水飞, 葛晓敏, 吴 军, 崔 鹏, 方炎明, 丁 晖,*
1 南京信息工程大学应用气象学院, 南京 210044 2 环境保护部南京环境科学研究所自然保护与生物多样性研究中心/国家环境保护武夷山生态环境科学观测研究站/国家环境保护生物安全重点实验室, 南京 210042 3 福建省武夷山生物研究所, 武夷山 354300 4 南京林业大学南方现代林业协同创新中心/生物与环境学院, 南京 210037
植物种群的空间分布特征一直是植物群落生态研究的重点之一[1]。林木空间结构一般是指林木个体在森林群落中的分布格局及其属性在空间上的排列方式,即林木之间树种、大小、分布等空间关系[2],它是森林群落最显著的特征。林木个体的空间布局不仅影响森林群落对光的截获、幼苗存活和更新等,还潜在地影响着群落内部物种的共存和维持机制的稳定[3]。分析植物群落内物种的空间结构和分布可以使我们在了解群落内物种分布的同时,认识到群落内不同物种的生理特性及其与环境因子的相互关系[4]。传统格局分布的研究手段多采用样方法、Ripley点格局分析法、双向轨迹方差分析和χ2检验等分析方法,然而因受研究尺度和分析方法的限制作用,这些方法均存在不同程度的局限性,无法全面客观地反映物种空间分布的特征[5]。本文采用基于最近4株相邻木的空间结构参数研究武夷山常绿阔叶林的空间结构特征摆脱了传统格局分析中尺度效应的影响,测量方法也十分便捷,只需调查相邻木的夹角大小便可获得准确的分布信息[3,6]。此外在一个群落中,优势种作为群落演替的内在驱动力,最能体现一个群落内部的空间结构和格局的形成过程[7],本研究通过分析优势种群的空间结构特征,使我们能够更加深刻地认识整个群落的发展过程和演变趋势[6-8]。
受长期人为干扰等因素的影响,福建武夷山地区的常绿阔叶林原生林已消失殆尽,现存多为天然次生林[8- 9]。为了保护这一宝贵的森林资源,对武夷山常绿阔叶林种群结构和更新的研究迫在眉睫。本研究以福建武夷山9.6hm2常绿阔叶林样地为研究对象,采用角尺度、混交度和大小比数来分别分析样地中乔木的水平分布格局,树种空间隔离程度及林木的生长优劣程度[10- 11]。同时,在此基础上采用结构参数的二元分布更为立体地展现武夷山常绿阔叶林的空间结构特征[7- 8,10]。最后,通过深入探讨乔木层中优势树种的空间结构特征,判断常绿阔叶林的群落演替水平和趋势[12],为今后深入研究森林种群格局和维持物种多样性奠定基础。
1 研究区概况
武夷山作为全球生物多样性保护的关键区之一,保存有世界同纬度带上最完整、最典型、面积最大的中亚热带原生性森林生态系统[13]。武夷山境内海拔1800m以上的山峰多达三十余座,由此形成一道天然屏障,冬季可阻挡和削弱北方冷空气的入侵,使该区具有降水量多,湿度大,垂直变化显著等特点[14]。武夷山年平均气温一般在8.5—18℃范围内,最热月(7月)均温为26.7℃,最冷月(1月)均温为8.3℃,相对湿度78%—84%,年均降水量一般为1486—2150mm,局部地方高达3000mm以上,年蒸发量为1000mm左右,有雾日超过60d[15]。该区域多低山丘陵,属红色砂砾岩分布区,地层构造属中生代白垩纪、为第三纪系沉积的“赤石群碎屑岩”地层[16]。武夷山属于典型的亚热带湿润季风气候,四季交替明显,水热条件良好,空气湿度较大,其群落外貌终年常绿,育有壳斗科、樟科、山茶科、木兰科和金缕梅科等常绿树种。同时,阔叶林的林冠层以下有藤本植物和少量附生植物等层间植物分布期间,结构较为整齐[17- 18]。
2 研究方法
2.1 样地的设置与调查
调查小组于2013年10至12月期间,在武夷山市星村镇四新村附近(地理坐标为27°35′24″N,117°45′55″E)选取并设立一处9.6hm2较具代表性的(东北-西南方向长400m,西北-东南长240m)常绿阔叶林永久固定监测样地,并对整个样地进行了全面踏查[18]。其中,样地所处的主山脊呈东北-西南走向,与长边基本平行,而西北-东南方向则由主山脊向两侧山脚延伸[19]。
首先利用TOPCON全站仪将整个样地划分为240个20m×20m大样方,并结合实时动态控制系统(Real-Time Kinematic, RTK)测定每个大样方四个角的高程[20]。再将每个大样方分成16个5m×5m的小样方,详细记录乔木层中所有胸径≥1cm林木的植物编号、树种名称、树高、胸径、冠幅、坐标等信息,并挂牌作标记。首次调查结果显示,该常绿阔叶林样地内包含有64973株独立乔木个体(包括分枝和萌枝),共171种,隶属于44科86属[19]。
2.2 数据处理方法
基于4株相邻木的角尺度等空间结构参数的应用已较为成熟,它们能准确分析森林群落内的林木个体空间分布特征[10]。角尺度等结构参数均是以一株中心树和围绕在它四周的4株最近相邻木为基本结构单元计算,具体操作方法如下:
(1)角尺度(Uniform angle index):将任意两个邻接最近相邻木的夹角的小角设为α,再比较α与标准角α0(最近相邻木均匀分布时的夹角)的大小关系,角尺度等于4个α小于α0所占的比例,展现了一个基本空间结构单元内的林木个体分布情况[7,21]。此外,我们还可以通过该样地所有乔木树种角尺度的平均值来判定一个大样地中的群落空间分布特征,即根据平均角尺度是否在随机分布置信区间[0.475,0.517]内来(区间内为随机分布;小于0.475为均匀分布;超出0.517则为团状分布)判别[22]。
(2)混交度(Mingling degree):通过最近4株相邻木中与参照树为不同树种所占的比例,描述了群落中乔木的树种隔离程度[7],可划分为零度、弱度、中度、强度和极强度混交5个等级[21]。
(3)胸径大小比数(Size ratio):比较4株最近相邻木大于参照树所占的比例[7],比例值越小,中心参照树的优势程度越高。最终,树种优势度可分为优势、亚优势、中庸、劣势、绝对劣势5个等级[21]。
为避免边缘效应的影响,研究首先在样地四周预设出5m的缓冲区,且缓冲区的乔木只作为相邻木来使用[23]。采用林分空间结构分析软件Winkelmass 1.0计算出该9.6hm2常绿阔叶林样地内所有乔木的角尺度、混交度和大小比数值。通过分析它们的一元、二元频率分布图,我们能够深层次地挖掘出乔木层中所隐藏的潜在空间异质性信息[24],二元分布图更是利用结构参数表达结构上的频率优势[25],充分直观地展示了结构参数组合的分布状况和数量特征[26]。本文采用Excel建立数据透视表,分别得出任意2个结构参数指标取值组合(Xi, Yj)的相对频率,并用Sigmaplot 12.5绘制Xi-Yj的二元分布图[27]。同时,通过R软件分析了角尺度、大小比数、混交度、平均胸径和平均树高的相关性,计算出它们之间的皮尔森相关系数(R)和P值[28]。最后,还对重要值前10位的优势树种的空间结构参数进行深入的分析,进一步了解现阶段武夷山常绿阔叶林群落的内部特征。
3 研究结果
3.1 武夷山常绿阔叶林的空间结构参数分布
3.1.1 武夷山常绿阔叶林3种空间结构参数的一元分布
样地的角尺度频率分布随着角尺度等级的增加(Uniform angle index=0.00→1.00)先增加后减小,右端频率值高于左端,呈负正态分布(图1)。整个样地的平均角尺度0.5707,因此该样地属于轻度聚集分布。其次,在所有混交度等级上,低混交度所占比例极少,且随着混交度等级的增加(Mingling degree=0.00→1.00),高混交度等级上的频率值迅速增大。整个样地的平均混交度为0.8359,表明样地中树木的混交程度较高,群落异质性很强(图1)。5个大小比数等级(Size ratio=0.00→1.00)上的频率值均在0.20上下,其平均大小比数为0.5057(图1),说明样地内乔木层的生长状况整体处于中庸状态。
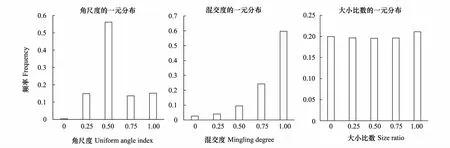
图1 武夷山常绿阔叶林角尺度、大小比数和混交度的一元分布Fig.1 The univariate distribution of the uniform angle index, size ratio and mingling degree of the evergreen broad leaved forest in Wuyi Mountains, Fujian, China
3.1.2 武夷山常绿阔叶林3种空间结构参数的二元分布
在角尺度和混交度的二元分布图中,多数相对频率值(约56%)分布在随机分布轴(Uniform angle index=0.50)上[25],且最高频率值出现在结构组合(0.50, 1.00)上,呈随机分布的结构单元在样地中占有最大比例,约为33.98%(图2)。同一角尺度等级上的频率值都随着混交度等级的降低而减少,并在混交度为零时几乎逼近零值。同时,角尺度等级越靠近随机分布轴,其频率值的减幅就越大。其中,每列混交度相对应的频率值随着角尺度等级的增加呈先增大后减小的趋势,且在高混交度等级上达到最大值。角尺度-大小比数二元分布图(图2)以随机分布轴和中庸轴为中心两侧大致呈对称分布,同一大小比数等级上的频率值均随角尺度的增大先增大后减小,在随机分布轴上取得最大值,进一步表明这些乔木整体上处于随机分布状态。当在同一角尺度等级上时,各大小比数上的频率值极为接近,说明角尺度对大小比数并没有直接的影响。混交度-大小比数的二元分布图(图2)中绝大部分值分布于中高混交等级轴(Mingling degree=0.50→1.00)上。同一大小比数等级上的频率值随着混交度等级的减少(Mingling degree=1.00→0.00)而迅速下降直至趋于零。不同优劣状态的林木在同一混交度等级上频率值较相近。
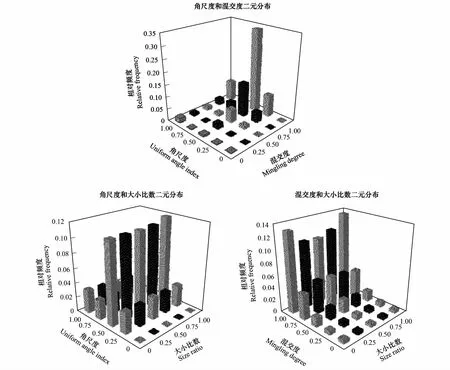
图2 武夷山常绿阔叶林角尺度、大小比数和混交度的二元分布Fig.2 The binary distribution of the uniform angle index, size ratio and mingling degree of the evergreen broad leaved forest in Wuyi Mountains, Fujian, China
3.1.3 武夷山常绿阔叶林空间结构参数的相关性
将样地内171个树种的3种空间结构参数与胸径、高度之间进行相关性分析可知(表1),大小比数作为衡量树种优势度高低程度的参数,与胸径(R=-0.81,P<0.01)和树高(R=-0.83,P<0.01)呈极显著负相关。混交度与树高之间呈现显著正相关性,这在一定程度上说明树种越高,其周围分布有其他树种的可能性越大。此外,在3种空间结构参数之间,只有角尺度和混交度之间存在极显著负相关性。

表1 空间结构参数与树高、胸径的相关性分析
3.2 武夷山常绿阔叶林中若干常见种的空间结构特征
前10个优势树种的角尺度集中分布在区间[0.55, 0.59]内,说明它们在该样地中均为轻度聚集分布(表2)。其中,毛锥、矩叶鼠刺等的角尺度数值较大,聚集程度也较高。再由大小比数可知,优势树种之间的优势度差异较大,最小为0.258,最大可达0.630。罗浮锥、木荷、甜槠、米槠等树种的大小比数值最小,在该常绿阔叶林中具有较大优势,且这些树种的大小比数与树高线性相关。同时,10个树种的混交程度均较高,都呈高度混交(或极高度)混交状态。综上所述,优势种中罗浮锥、木荷和米槠的优势程度高,其周围树种的混交度也极高,在整个样地内具有一定的生长优势。赤楠、格药柃、矩叶鼠刺和弯蒴杜鹃虽个体数量多,但它们的大小比数较大,在群落内的优势程度低,多处于乔木下层。受光照等环境因素的影响,该常绿阔叶林内占据优势的树种多为阳生树种,如木荷、甜槠等树种;而赤楠等喜阴树种则多处于乔木下层。
表2武夷山常绿阔叶林前10个优势树种的3种空间结构参数
Table2Thespatialstructureparametersofthetop10dominanttreespeciesintheevergreenbroadleavedforestintheWuyiMountains,Fujian,China
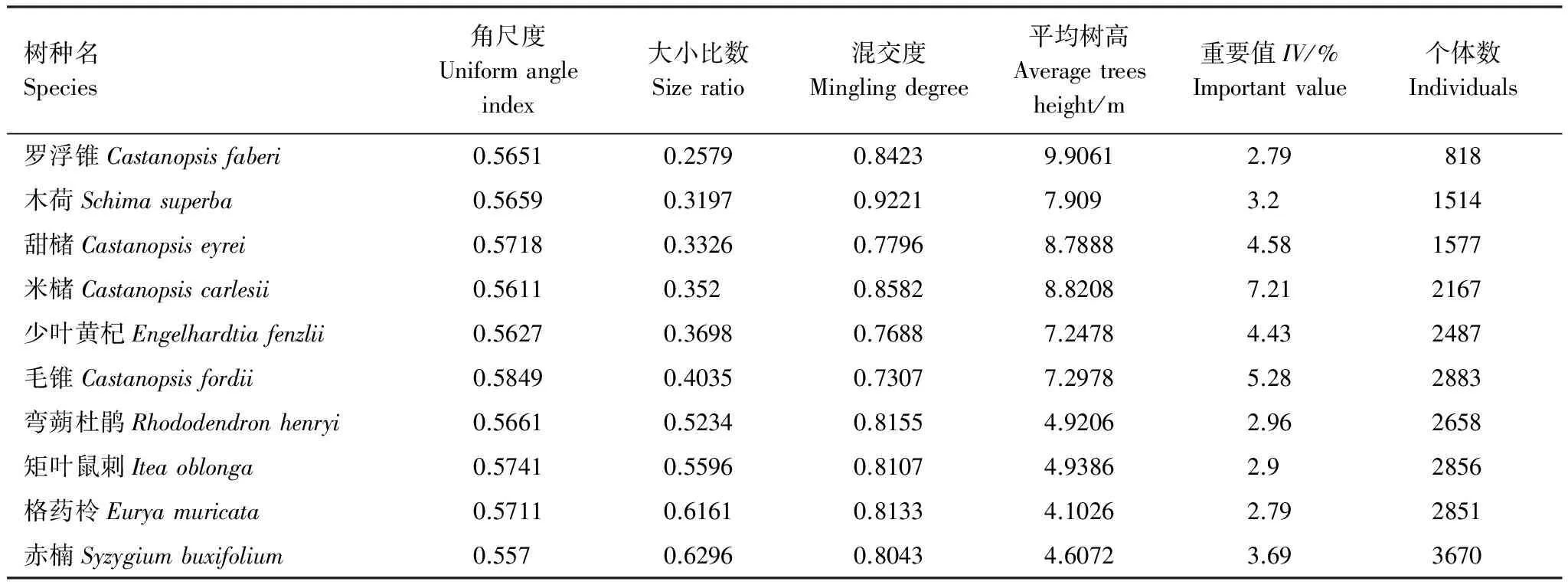
树种名Species角尺度Uniformangleindex大小比数Sizeratio混交度Minglingdegree平均树高Averagetreesheight/m重要值IV/%Importantvalue个体数Individuals罗浮锥Castanopsisfaberi0.56510.25790.84239.90612.79818木荷Schimasuperba0.56590.31970.92217.9093.21514甜槠Castanopsiseyrei0.57180.33260.77968.78884.581577米槠Castanopsiscarlesii0.56110.3520.85828.82087.212167少叶黄杞Engelhardtiafenzlii0.56270.36980.76887.24784.432487毛锥Castanopsisfordii0.58490.40350.73077.29785.282883弯蒴杜鹃Rhododendronhenryi0.56610.52340.81554.92062.962658矩叶鼠刺Iteaoblonga0.57410.55960.81074.93862.92856格药柃Euryamuricata0.57110.61610.81334.10262.792851赤楠Syzygiumbuxifolium0.5570.62960.80434.60723.693670
4 讨论
4.1 武夷山常绿阔叶林3种空间结构参数
4.1.1 武夷山常绿阔叶林的空间分布特征
森林是区域生态系统的重要组成部分,研究森林群落内部的空间结构分布情况对优化区域整体结构、发挥区域生态功能价值具有非常重要意义[29- 30]。一个树种的空间分布是其受多种环境因子共同作用后的综合反映,如当个体间存在激烈竞争时,林木个体多呈均匀分布;个体共存的空间关系多表现为聚集分布;而当个体互相独立时则呈现随机分布[31]。很多生态学者证实自然环境中大多数物种表现为聚集分布,森林物种分布的优势格局多为聚集格局[5],如浙江古田山[32],广东鼎湖山[33],云南哀牢山[34],甘肃小陇山[8],巴拿马热带雨林[35]等。森林演替一般以自然入侵更新为主,易受树种自身生长特性、区系和人为(或自然)干扰因素的影响,树种聚集分布在一起分布可以提高自身的生存几率[1,5,36]。本研究由结构参数的一元分布获得武夷山常绿阔叶林的宏观空间分布特征,样地以随机分布为主,聚集分布为辅,聚集分布结构单元明显多于均匀分布;样地内树种的混交程度极高,具有很强的物种异质性;同时,树种的优劣程度参差不齐,整体处于中庸的生长状态。刘韶辉在对湖南会同亚热带次生阔叶林的群落特征分析时发现,群落优势树种随着演替的加剧会通过种内聚集来缓解种间竞争,使系统向稳定共存条件转化,其次生阔叶林中的落叶树种演替到后期会变成常绿阔叶树种[37]。而康迪对黄土高原丘陵沟壑区典型恢复植被群落的研究也表明,群落种间空间相关性在演替初期往往趋向随机,没有明显的种间关联,随着演替的深化,具有相似生活型或相互有利的物种会相对集聚,演替至顶级群落时所保存的物种保持互利共存的关联性,以保证群落的相对稳定[38]。本研究中的武夷山常绿阔叶林样地呈强度混交的轻度聚集分布格局,具有较高的树种异质性,因此,该常绿阔叶林的物种演替已达到较高的水平,可能已达到演替中后期,群落内大量物种共存,继续向顶级群落演替。
然而,一元分布在表达结果的形式上较为单一,无法满足多样性分析的需求[39]。图2中结构参数的二元分布分别将角尺度、混交度和大小比数进行两两联合,以三维空间的形式很好地展示武夷山常绿阔叶林群落的空间结构特征。结果表明,该样地中高混交度、随机分布结构组合的比例远远高于其他任何一个组合,即绝大多数林木个体处于强度混交、随机分布状态;无论树种的优劣程度如何,绝大多数林木个体以与其他树种共生的形式生存,而乔木层明显的垂直结构分化可能是其主要原因[40],不同的树种因其树高的差异,分别在不同的高度利用着光照资源,得以在水平方向的较小范围内共存;不同优势度等级上的角尺度分布均呈现相似的规律,同一分布类型上的林木优劣程度也相差不大,可见树种的优劣程度与其分布类型不存在直接关系。上述结果进一步印证了该常绿阔叶林具有高度异质性的空间结构,且有大量树种共生演替。一般地,随着演替的不断增强,群落内的物种组成和结构变得越来越复杂多变[41]。该样地乔木层内的树种隔离程度虽然较高,但其树种优势程度普遍不高,当前未有明显的优势建群种出现,与丁晖等在该样地所做的研究结果相一致[19]。
4.1.2 武夷山常绿阔叶林空间结构参数的相关性
相关性分析结果表明,树种的混交度会与其平均树高呈显著正相关。一方面,树种的胸径和树高对其优势度有显著影响,树种的胸高断面积越大,树高越高,代表其生物量的积累越多,那么其在样地内的优劣程度也就越大。另一方面,根据He和Duncan所提出的负密度制约理论[42]可知,许多树种在幼年时会都聚集分布在一起,随着年龄的增长,其种群内部竞争开始愈发激烈,受自疏作用的影响,只有少数个体存活并长成大树。在此期间,随着群落内部微环境的改变,其他树种渐渐侵入,在提高该区域物种混交度的同时,大大增加了其物种多样性,有助于群落形成较为稳定的动态结构[43]。再看3个空间结构参数的空间相关性可知,只有角尺度和混交度之间存在极显著负相关性,即角尺度越大(邻近木越聚集),其周围物种的混交程度就越低(树种周围为不同种树的可能性越小)。表明树种的多样性越高,异质性越高,其分布也就越均匀。受物种特性和环境等众多因素的影响,许多树种会采取忍耐生存策略(其树种分布多为聚集分布))来提高自身生存机率,此时树种的混交程度通常会较低,其群落内的物种多样性也不高,反之亦然。
4.2 常绿阔叶林中优势树种的空间分布及其演替规律
为探讨武夷山常绿阔叶林内部结构的动态变化特征和维持机制,本研究对乔木层中重要值排前10个优势树种的角尺度、混交度和大小比数进行研究。王志泰等在研究东祁连山高寒地区柳灌丛群落种间关联性时曾指出,树种间的竞争、排斥等相互作用对维持森林结构和功能的稳定非常有益[44]。而上述结果表明,武夷山常绿阔叶林乔木层现阶段的竞争较为激烈,林内林木也以幼、中龄林为主,属于典型的恢复性植被。10个优势种不仅隔离程度都极高,树种的优劣程度差距也较大。其中,木荷、米槠和罗浮锥等喜阳树种呈混交度高,其优劣程度较大的状态,更新状况良好,在整个群落内具有一定的生长优势,占据着乔木上层阳光充足的地方;而赤楠、格药柃和矩叶鼠刺等喜阴树种个体数量虽多,但普遍不高,平均树高均在5m以下,广布于乔木下层,与此同时,它们树种的优势程度较低,大小比数值均超出0.50,可见赤楠等多处于乔木亚层,在该群落内生长优势并不显著。Loucks在威斯康辛南部森林[45]和Bazzaz在美国伊利诺斯州南部落叶阔叶林区弃耕地[46]的研究结果表明,演替一开始,喜阳植物首先侵入,并在期间占据有利的生长优势;接着就会有喜阴植物不断涌入,并在喜阳植物下广布繁殖;最后,当喜阳和喜阴植物的同时出现而使得样地内的物种多样性达到一定程度时,该区域已到达演替中后期。由此可以推断该武夷山常绿阔叶林当前可能处于演替中后期,未来我们将对该9.6hm2样地进行持续的动态观测,以期更加充分地证实本研究所得结论,并进一步为探索武夷山常绿阔叶林的更新方式和演替规律提供科学依据。
致谢:武夷山世界遗产监测中心提供试验场地,特此致谢。
参考文献(References):
[1] 尤海舟, 刘兴良, 缪宁, 何飞, 马钦彦. 川滇高山栎种群不同海拔空间格局的尺度效应及个体间空间关联. 生态学报, 2010, 30(15): 4004- 4011.
[2] 龚直文, 亢新刚, 顾丽, 赵俊卉, 郑焰锋, 杨华. 天然林林分结构研究方法综述. 浙江林学院学报, 2009, 26(3): 434- 443.
[3] 惠刚盈, 李丽, 赵中华, 党普兴. 林木空间分布格局分析方法. 生态学报, 2007, 27(11): 4717- 4728.
[4] 陶冶, 张元明, 吴小波. 植物种群的多尺度多参数空间分布格局——以白茎绢蒿和准噶尔沙蒿为例 // 中国植物学会第十五届会员代表大会暨八十周年学术年会论文集. 南昌: 中国植物学会, 2013: 134- 135.
[5] 宋厚娟, 叶吉, 蔺菲, 原作强, 郝占庆, 王绪高. 取样面积对森林木本植物空间分布格局分析的影响. 科学通报, 2014, 59(24): 2388- 2395.
[6] 惠刚盈, Gadow K V, 胡艳波, 陈伯望. 林木分布格局类型的角尺度均值分析方法. 生态学报, 2004, 24(6): 1225- 1229.
[7] 张岗岗, 王得祥, 柴宗政, 张丛珊, 刘文桢, 张宋智. 小陇山2种典型天然林空间结构参数分布特征. 林业科学研究, 2015, 28(4): 531- 537.
[8] 曹小玉, 李际平, 封尧, 胡园杰, 张彩彩, 房晓娜, 邓超. 杉木生态公益林林分空间结构分析及评价. 林业科学, 2015, 51(7): 37- 48.
[9] 周娟, 陈丽华, 郭文体, 李胜男, 高洪翠, 李长暄, 李海燕. 辽东山地山杨天然次生林空间结构分析. 西北农林科技大学学报: 自然科学版, 2014, 42(8): 79- 85, 98- 98.
[10] 惠刚盈. 基于相邻木关系的林分空间结构参数应用研究. 北京林业大学学报, 2013, 35(4): 1- 8.
[11] 惠刚盈, 胡艳波, 徐海. 森林空间结构的量化分析方法 // 2005年中国科协学术年会26分会场论文集. 北京: 中国科学技术协会, 2005: 47- 50.
[12] 董灵波, 刘兆刚, 马妍, 倪宝龙, 李元. 天然林林分空间结构综合指数的研究. 北京林业大学学报, 2013, 35(1): 16- 22.
[13] 陈晓. 武夷山常绿阔叶林若干优势树种的生态位分析. 森林与环境学报, 2016, 36(3): 337- 341.
[14] 李荣禄. 为世罕见的模式标本产地——武夷山生物圈保护区. 中国生物圈保护区, 1999, (2): 3- 3.
[15] 胡砚秋, 李文斌, 崔佳玉, 苏志尧. 亚热带常绿阔叶林优势种个体及生物量的点格局分析. 生态学报, 2016, 36(4): 1066- 1072.
[16] 武夷山市志编纂委员会. 武夷山市志. 北京: 中国统计出版社, 1994: 103- 259.
[17] 陈婷婷, 徐辉, 马方舟, 陈水飞, 徐海根, 方炎明, 杨青, 徐鲜钧, 丁晖. 武夷山亚热带常绿阔叶林物种多样性的尺度效应. 生态与农村环境学报, 2016, 32(5): 750- 756.
[18] 丁晖, 杨云方, 徐海根, 方炎明, 陈晓, 杨青, 伊贤贵, 徐辉, 温小荣, 徐鲜钧. 武夷山典型常绿阔叶林物种组成与群落结构. 生态学报, 2015, 35(4): 1142- 1154.
[19] 丁晖, 方炎明, 杨青, 陈晓, 袁发银, 徐辉, 何立恒, 严靖, 陈婷婷, 余朝健, 徐海根. 武夷山中亚热带常绿阔叶林样地的群落特征. 生物多样性, 2015, 23(4): 479- 492.
[20] 吕寻, 杨双宝, 刘文桢, 郭小龙, 李安民, 袁一超. 小陇山锐齿栎原始群落林木径阶空间结构特征. 北京林业大学学报, 2015, 37(5): 11- 18.
[21] 仇建习, 汤孟平, 沈利芬, 娄明华, 庞春梅. 近自然毛竹林空间结构动态变化. 生态学报, 2014, 34(6): 1444- 1450.
[22] 胡艳波, 惠刚盈, 王宏翔, 李远发, 赵中华, 刘文桢. 随机分布的角尺度置信区间及其应用. 林业科学研究, 2014, 27(3): 302- 308.
[23] 周红敏, 惠刚盈, 赵中华, 胡艳波. 林分空间结构分析中样地边界木的处理方法. 林业科学, 2009, 45(2): 1- 5.
[24] 张岗岗, 王得祥, 柴宗政, 朱红燕, 张丛珊. 秦岭中段华北落叶松人工林空间结构的二元分布特征. 西北农林科技大学学报: 自然科学版, 2014, 42(9): 33- 40.
[25] 李远发. 林分空间结构参数二元分布的研究[D]. 北京: 中国林业科学研究院, 2013: 29- 32.
[26] Li Y F, Hui G Y, Zhao Z H, Hu Y B. The bivariate distribution characteristics of spatial structure in natural Korean pine broad-leaved forest. Journal of Vegetation Science, 2012, 23(6): 1180- 1190.
[27] 李远发, 王磊, 喻素芳, 叶绍明, 陆道调. 广西滨海过渡区红鳞蒲桃群落的空间结构特征. 生态学杂志, 2016, 35(3): 726- 732.
[28] 李侃竹, 高品, 王凯, 刘振鸿, 薛罡. 污水中抗生素与重金属对红霉素抗药性基因的选择性效应. 中国环境科学, 2015, 35(3): 889- 896.
[29] 胡美娟, 侯国林, 周年兴, 李在军, 亓秀云, 方叶林. 庐山森林景观空间分布格局及多尺度特征. 生态学报, 2015, 35(16): 5294- 5305.
[30] 王祎玲, 张钦弟, 郝晓杰, 闫桂琴. 山西七里峪茶条槭的种群结构与空间分布格局. 西北植物学报, 2012, 32(5): 1027- 1035.
[31] 龚直文, 顾丽, 亢新刚, 杨华, 赵浩彦. 长白山森林次生演替过程中林木空间格局研究. 北京林业大学学报, 2010, 32(2): 92- 99.
[32] 李立, 陈建华, 任海保, 米湘成, 于明坚, 杨波. 古田山常绿阔叶林优势树种甜槠和木荷的空间格局分析. 植物生态学报, 2010, 34(3): 241- 252.
[33] 叶万辉, 曹洪麟, 黄忠良, 练琚愉, 王志高, 李林, 魏识广, 王章明. 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究. 植物生态学报, 2008, 32(2): 274- 286.
[34] 巩合德, 杨国平, 鲁志云, 刘玉洪. 哀牢山常绿阔叶林树种多样性及空间分布格局. 生物多样性, 2011, 19(2): 143- 150.
[35] He F L, Legendre P, LaFrankie J V. Distribution patterns of tree species in a Malaysian tropical rain forest. Journal of Vegetation Science, 1997, 8(1): 105- 114.
[36] Graz F P. Spatial diversity of dry savanna woodlands. Biodiversity & Conservation, 2006, 15(4): 1143- 1157.
[37] 刘韶辉. 湖南会同亚热带次生阔叶林群落特征及种间关系研究[D]. 长沙: 中南林业科技大学, 2011: 80- 85.
[38] 康迪. 黄土高原丘陵沟壑区典型恢复植被群落结构及演替规律[D]. 杨凌: 西北农林科技大学, 2013: 34- 44.
[39] Pommerening A, Biber P, Pretzsch H, Stoyan D. Neue methoden zur analyse und charakterisierung von bestandesstrukturen. Forstwissenschaftliches Centralblatt vereinigt mit Tharandter forstliches Jahrbuch, 2000, 119(1/6): 62- 78.
[40] 商侃侃, 王婕, 余倩, 赵明水, 达良俊. 天目山第三纪孑遗植物缺萼枫香的群落特征及其生态学意义. 华东师范大学学报: 自然科学版, 2011, (3): 134- 144.
[41] 王世雄, 王孝安, 李国庆, 郭华, 朱志红. 陕西子午岭植物群落演替过程中物种多样性变化与环境解释. 生态学报, 2010, 30(6): 1638- 1647.
[42] 祝燕, 米湘成, 马克平. 植物群落物种共存机制: 负密度制约假说. 生物多样性, 2009, 17(6): 594- 604.
[43] 张立敏, 陈斌, 李正跃. 应用中性理论分析局域群落中的物种多样性及稳定性. 生态学报, 2010, 30(6): 1556- 1563.
[44] 王志泰, 包玉, 李毅. 东祁连山高寒草地柳灌丛群落种间关联性. 生态学杂志, 2006, 25(10): 1177- 1180.
[45] Loucks O L. Evolution of diversity, efficiency, and community stability. American Zoologist, 1970, 10(1): 17- 25.
[46] Bazzaz F A. Plant species diversity in old-field successional ecosystems in southern Illinois. Ecology, 1975, 56(2): 485- 488.
猜你喜欢
小哥白尼(野生动物)(2022年5期)2022-08-15
公民与法治(2020年17期)2020-10-27
绿色中国(2019年18期)2020-01-04
红土地(2018年12期)2018-04-29
农家科技下旬刊(2017年8期)2017-11-13
科技创新导报(2016年21期)2016-12-17
