
农业景观非农生境植物多样性及其影响因素
2018-04-19 01:21卢训令梁国付丁圣彦
生态学报 2018年5期
卢训令,梁国付,汤 茜,丁圣彦,*
1 河南大学黄河中下游数字地理技术教育部重点实验室,开封 475004 2 河南大学环境与规划学院,开封 475004 3 河南大学生态科学与技术研究所,开封 475004
在全球人口激增的背景下,农业用地持续扩张,集约化日益增强,统计数据显示目前农业用地已经占据了地球陆地表层非冰雪覆盖面积的40%[1-2],农业景观已成为地球陆地表层最重要的景观类型,农业用地进一步扩展的空间已很有限[3]。而随着农业用地持续扩张,土地利用强度和集约化程度的日益增强,与之相伴的是农业景观中生物多样性的持续丧失和生态系统服务持续性的下降和波动[4- 7]。如何能够在维持粮食供给的同时降低农业活动对环境的负效应,从而实现农业景观的持续健康发展是本世纪人类面临的最大挑战[1,8- 9]。研究表明,农业景观中非农生境的存在能有效的提高植物、鸟类、地表动物、微生物等的多样性[10-15]。而同时还发现,景观背景的差异、尺度效应和跨营养级的生物级联效应会也会直接或间接的影响到生物多样性的变化和响应[16-19],但缺乏明确的结论。那么,景观异质性特征和群落结构会对植物多样性的维持产生什么样的影响?在不同尺度下,各影响因子的相对重要性如何?这些均是未来探讨农业景观生物多样性维持措施的重要前提和基础。
从景观水平上探讨生物多样性保护和生态系统服务是景观生态学研究中的重要论题[20],同时在景观尺度下推出的管理措施才能更有效和易于施行,且研究发现在多尺度下关注生物多样性保护策略会更有效[21]。研究表明,景观异质性、景观连接度、景观复杂度、景观基质等不同景观结构参数均可能对区域生物多样性产生不同程度的影响,而且具有一定的尺度效应[22- 23]。目前景观结构的时空异质性在不同尺度上对生物多样性有何影响仍是生态学研究中的重要论题之一,且依然存在着较多的争论与争议[24],通过在不同空间尺度上的研究有望能得出更新颖和更稳健的结论[25]。研究者希望能找出景观异质性与区域生物多样性特征间关系的尺度性。
本研究以地处黄河中游的巩义市为研究区,探讨区域内不同非农生境(山地次生灌丛、次生林地、人工林地、农田边缘)中植物群落物种多样性特征的差异,分析在不同空间尺度下,景观异质性和群落结构特征等对物种多样性特征和组成的影响效应,并探讨景观异质性特征与植物群落物种多样性特征间关系的尺度特征,从而为未来区域内农业景观中植物多样性保护措施的提出提供理论依据。
1 研究区概况
研究区位于巩义市,地处黄河中游,地理坐标为34°31′—34°52′N,112°49′—113°17′E之间,总面积约1042.8km2,地貌上横跨我国第二、三阶梯,地势自南向北呈阶梯状迅速降低,东南部为中低山地、向北经丘陵到河谷平原,景观结构复杂,土壤以山地棕壤、丘陵黄土和平原潮土为主,属暖温带大陆性季风气候,多年平均降水量约583mm,年均温14.6℃,地带性植被为落叶阔叶林,目前仅在南部中山区有小片原始林零星分布,其他山地上多为次生林和次生灌丛,丘陵和平原地带多为农田和少量的人工植被(图1)。
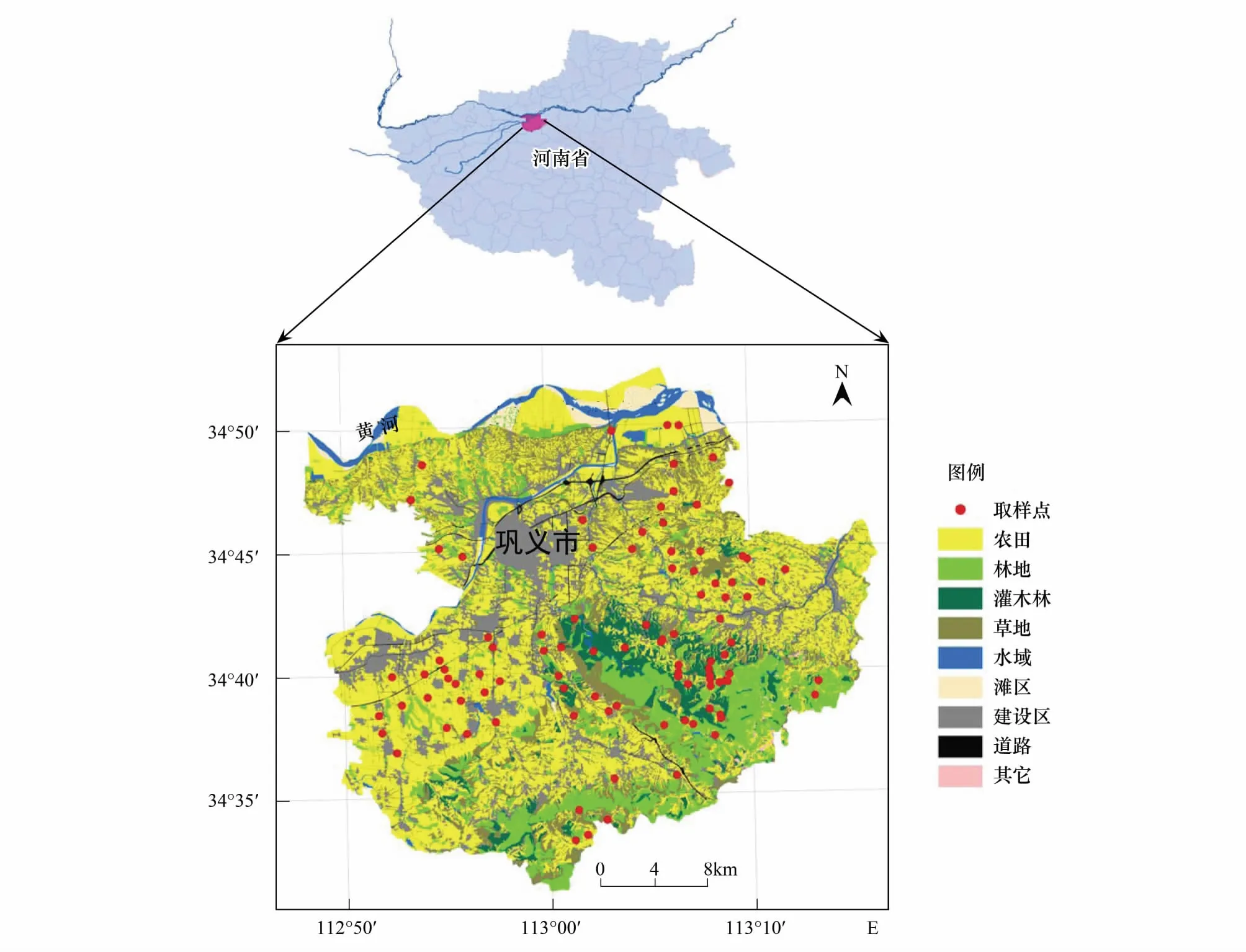
图1 研究区景观概况及野外调查样点分布图Fig.1 Landscape classification map and the field investigation plots distribution
2 研究方法
2.1 样地设置和数据收集
在研究区卫星影像、地貌分区和土地利用现状图的基础上,对区内的山地、丘陵和平原上各研究样点中主要非农生境(次生灌丛、次生林地、人工林地和农田边缘等)里的植物群落采用法瑞学派的典型样地法进行调查,记录所有调查物种的名称、多优度、群集度、物候期和各样方的地理坐标等。野外调查在2012—2015年的5—8月间进行,在98个调查样点上,共计调查有效样方910个(其中山地次生灌丛210个,次生林地60个,人工林地295个,农田边缘345个)(图1)。因山地次生灌丛和农田边缘生境基本不含乔木层,人工林地绝大多数是杨树或泡桐纯林,因此本文在数据分析过程中仅对灌草层物种多样性进行探讨。
2.2 数据分析方法
(1)物种丰富度:以样地中实际出现的物种数目S表示。为区别常见种和偶见种对群落贡献的差异性,采用稀疏化法对物种丰富度进行随机重抽样计算和外推,并绘制物种——样点数量累积曲线图,估算各生境中的物种丰富度和多样性特征[26-27],从而更有效的显示物种丰富度特征。
(2)物种丰富度的拆分:对植物多样性数据进行加性分配分析时,将多样性数据定义为两个水平:样方尺度和样点尺度;随机化重抽样次数为100;结果中α为样方水平的物种丰富度,显示的是样方尺度上物种丰富度的平均值,β1为样点尺度上样方间的多样性差异,显示的是样点尺度和样方尺度上物种丰富度间的差值,α+β1=α样点,β2为样点间多样性的差异,显示的是总物种丰富度与样点尺度上物种丰富度间的差值。具体计算过程在PARTITION 3.0中进行[28]。
(3)景观异质性和群落结构特征对生物多样性影响效应分析:首先,依据研究区土地利用现状图(2011年,1∶1万)绘制研究区景观分类图,共分为:水浇地、旱地、果园、林地、灌木林、草地、河流、坑塘、沟渠、滩地景观、建设用地、道路和裸地共13种景观类型(图1)。以各取样点为圆心,计算不同缓冲区半径内的景观异质性特征。缓冲区半径设置为:200、300、400、500、600、800、1000、1250、1500、1750、2000、2500、3000m;使用Fragstats 4.2对不同半径缓冲区域内的景观异质性特征进行计算,并通过对指数间的共线性分析,最终从景观水平上选取面积-边缘类指数(Area-Edge index)、形状类指数(Shape index)、聚合度类指数(Aggregation)和景观多样性指数(Diversity)等4大类共19个指数来表征研究区不同取样点上的总体异质性特征。其次,选取灌木层高度、盖度,草本层高度、盖度等指标来表征群落结构特征。并选取样方尺度的物种丰富度(α样方)、样点尺度的物种多样性(α样点)、β多样性(β2)、均匀度指数(E)、Shannon多样性指数(H′)和优势度指数(D′)等表征群落物种多样性特征(表1)。最后借助多元回归模型分析(逐步回归,P<0.05)探讨在不同空间尺度下景观异质性和群落结构特征与植物多样性特征间的关系。数据处理和分析在Microsoft Office Excel 2010和SPSS 19.0中进行。
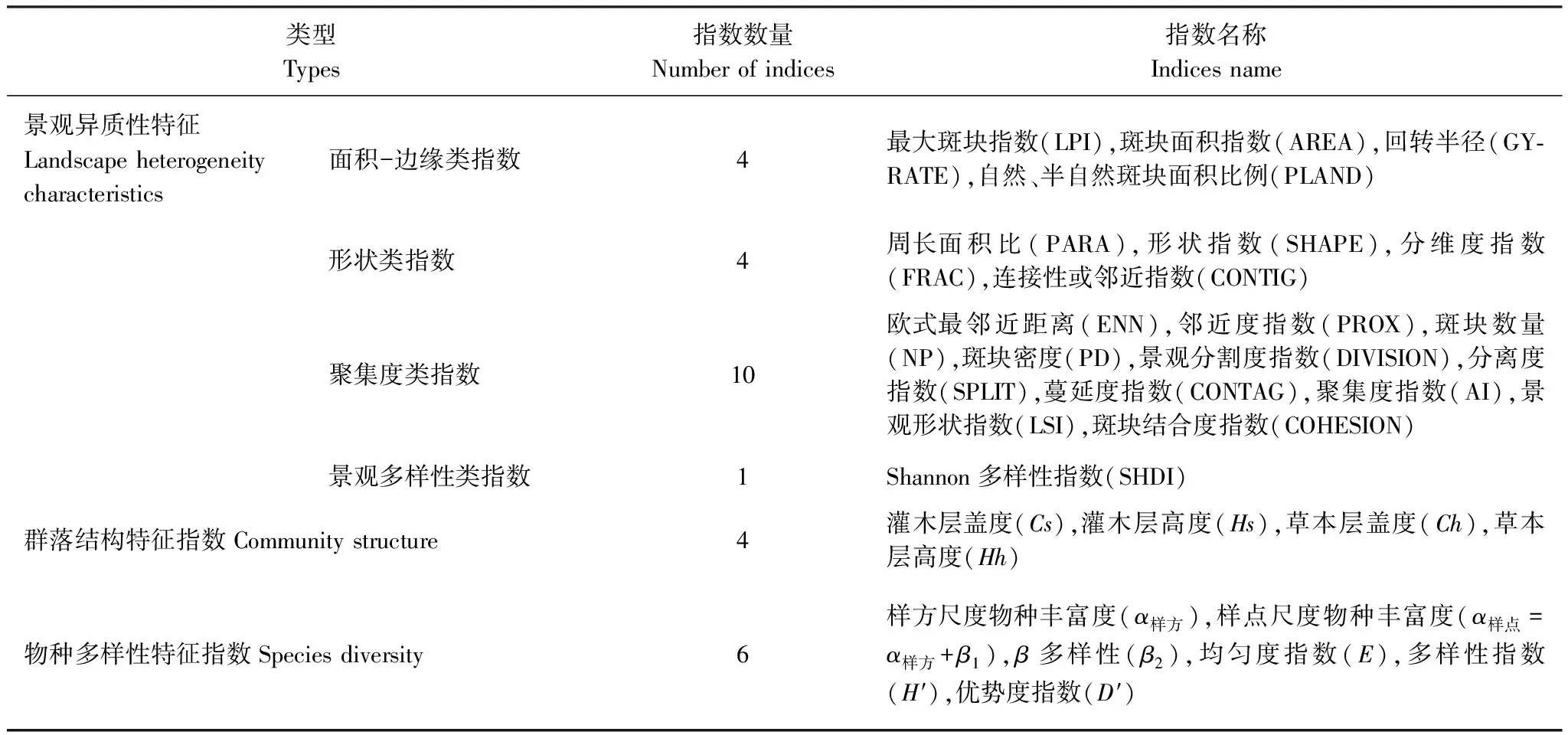
表1 景观异质性指数的选取
LPI:Largest patch index;AREA:Patch area;GYRATE:Radius of gyration;PLAND:Percentage of landscape;PARA:Perimeter-area ratio;SHAPE:Shape index;FRAC:Fractal dimension index;CONTIG:Contiguity index;ENN:Euclidean nearest neighbor distance;PROX:Proximity index;NP:Number of patches;PD:Patch density;DIVISION:Landscape division index;SPLIT:Splitting index;CONTAG:Contagion;AI:Aggregation index;LSI:Landscape shape index;COHESION:Patch cohesion index;SHDI:Shannon′s diversity index
3 研究结果
3.1 各生境中物种多样性特征
采用EstimateS 9.1.0对物种丰富度数据进行稀疏化处理,绘制稀疏化曲线,并基于抽样统计的方法将各生境物种丰富度数据统一外推至70个样点,得到物种丰富度-样点数量累积曲线图。并对Shannon-wiener多样性指数稀疏化结果绘图(图2)。
结果显示,物种丰富度以次生灌丛和农田边缘生境中最高(实际调查数据和估测数据分别为299种、329.28种和254种、254.86种),人工林生境中的物种丰富度稍低于农田边缘,共调查到235种,(估测值为243.04种),次生林生境中的物种丰富度最低(实调与估测数据分别为143种和204.34种);方差分析显示,次生灌丛生境中物种丰富度显著高于其他生境,次生林生境则显著低于其他3类生境(P<0.05)。Shannon-wiener多样性指数显示,次生灌丛中物种多样性最高(4.4),且方差分析表明该生境显著高于其他3类生境(P<0.05),人工林和农田边缘生境中物种多样性次之,均为4.12,次生林生境中最低(3.68),显著低于其他生境(P<0.05)(图2)。
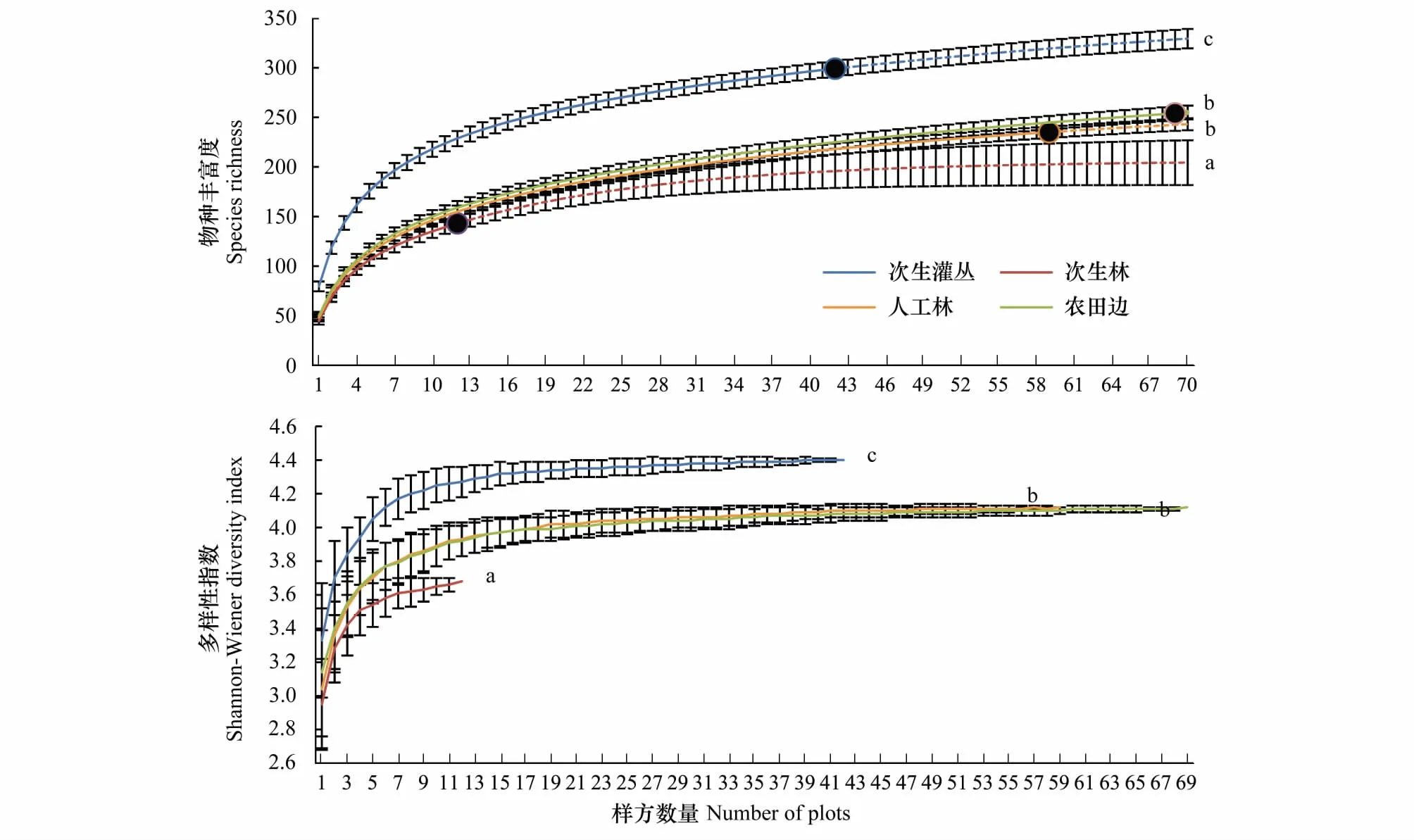
图2 不同生境中物种丰富度和多样性指数稀疏化与外推曲线图Fig.2 The rarefaction and extrapolation curves of plant species richness and diversity index in different habitats图中实线段为实际调查物种丰富度稀疏化结果,实心圆为实际调查物种丰富度值,虚线段为外推结果曲线,误差线为标准差;图上字母a、b、c表示方差分析的结果,P<0.05
3.2 物种多样性拆分特征
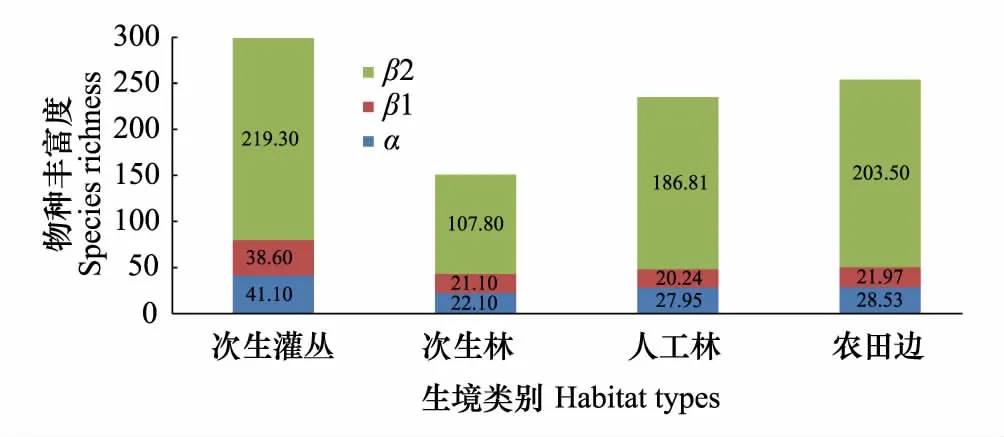
图3 不同生境中物种丰富度的加性分配拆分图 Fig.3 Additive partitioning of plant species richness in different habitats 图中数字为不同层次物种丰富度数值,误差线为标准差,α为样方尺度物种多样性,β1为样方间物种多样性差异,β2为样点间的物种多样性差异
物种丰富度拆分(加性分配)结果显示(图3):总体上各生境中物种丰富度组成均以样点间的物种多样性差异(β2)占绝对优势(农田边:203.5种,80.1%;人工林:186.8种,79.5%;次生灌丛:219.3种,73.3%;次生林:107.8种,71.4%);样方尺度上物种丰富度(α)对各生境中总物种丰富度的贡献性均较低,约在11%—15%之间;样方间物种多样性差异(β1)在各生境总物种丰富度中的比例最低,人工林和农田边缘生境中仅为8.6%和8.7%,次生灌丛和次生林生境中稍高,但也仅为12.9%和14.0%。
从不同生境中各层次物种丰富度组成来看,样方尺度上的物种丰富度(α)和样方间物种多样性差异(β1)在次生林和次生灌丛两类自然生境中,其对总物种丰富度的贡献要明显高于人工林和农田边缘等两类生境,尤其是β1的差异更为明显。
3.3 景观异质性和群落结构特征与生物多样性的关系
通过对景观异质性和群落结构特征指数与表征生物多样性特征的各指数间的多元回归模型分析(表2)发现:总体上,群落结构特征对植物多样性特征的影响更为直接和显著,但各指标间又有一定的差异:样方尺度上的物种多样性在小尺度上(缓冲区半径≤600m)与草本层盖度的关系最为密切,而在600m半径之外表征景观异质性特征的自然、半自然斑块(包括林地、灌木、草地等)面积比例(PLAND)与之关系更为密切;草本层高度(Hh)在各尺度上均对样点尺度的物种多样性(α样点)、β多样性(β2)和物种均匀度指数(E)的影响最为显著;Shannon多样性指数(H′)则更主要的受群落中草本层盖度(Ch)的影响;而灌木层盖度(Cs)与群落物种优势度指数(D′)关系最为密切。
从各回归模型的解释力(调整R2)来看(图4),景观异质性和群落结构特征对群落β多样性(β2)和样点尺度的物种丰富度(α样点)解释力最强,其次是样方尺度的物种丰富度(α样方),而对均匀度指数、Shannon多样性指数和优势度指数的解释力相对较低。从空间尺度来看,除β多样性和样点尺度的物种丰富度之外,其他各指数的回归方程基本都呈现倒“~”形状(图4),即在较小尺度上,景观异质性和群落结构特征能较好的拟合物种多样性特征,随着空间尺度(300—800m)的增加,这种关系有一定的减弱,但在超过800m之后会转而升高,且在大约1750—2500m左右达到高值,并转而随着空间尺度的进一步增大而降低。
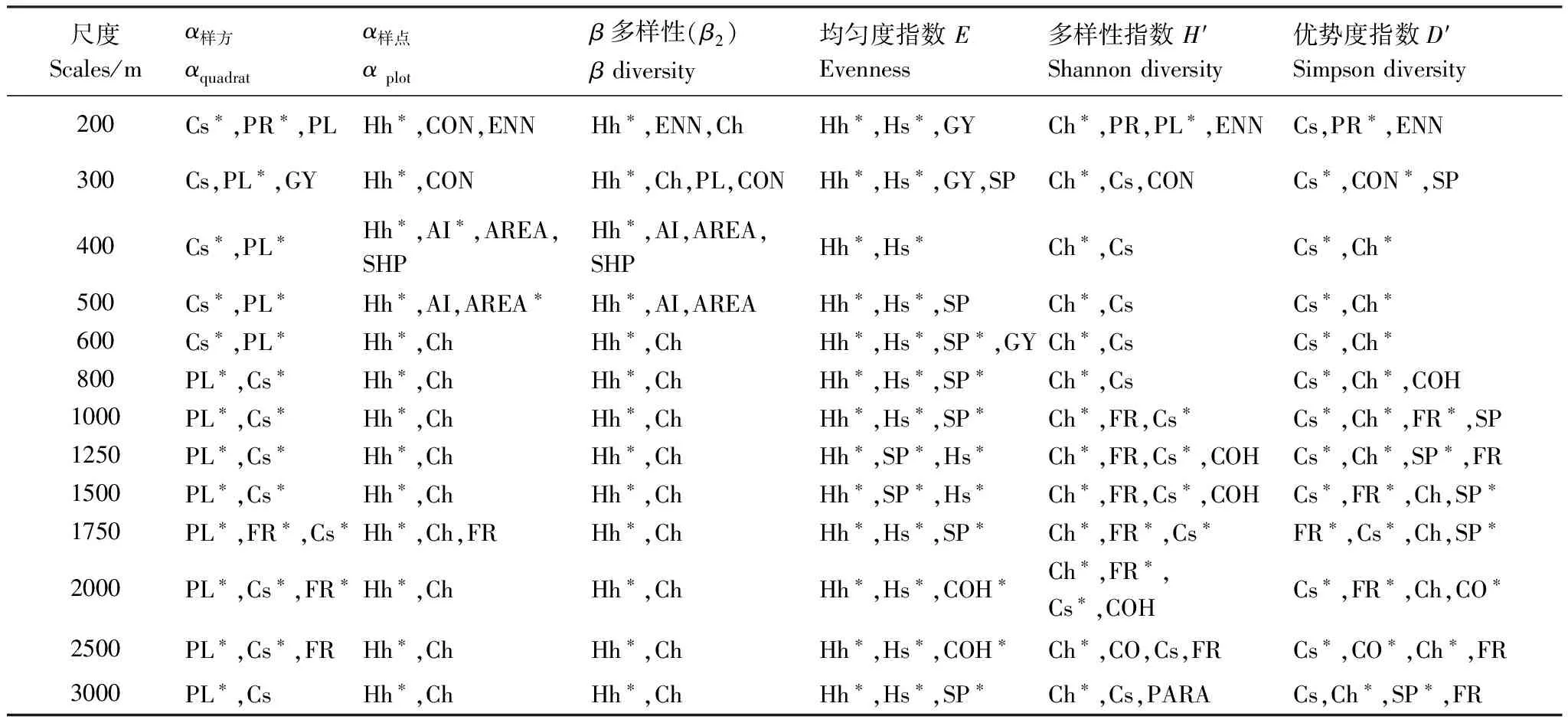
表2 景观异质性指数和群落结构指标与生物多样性特征指标间关系的多元回归结果
*为极显著(P<0.01);各缩写的意义如下:Cs:灌木层盖度,Coverage of the shrub layer;Ch:草本层盖度,Coverage of the herb layer;Hs:灌木层高度,Height of the shrub layer;Hh:草本层高度,Height of the herb layer;PR:邻近度指数,Proximity index (PROX);CON:蔓延度指数,Contagion index (CONTAG);PL:自然、半自然斑块面积比例,Percentage of landscape (PLAND);COH:斑块结合度指数,Patch cohesion index (COHESION);ENN:欧式最邻近距离,Euclidean nearest neighbor distance;AI:聚集度指数,Aggregation index;SP:分离度指数,Splitting index (SPLIT);FR:分维数,Fractal dimensions index (FRAC);SHP:形状指数,Shape index (SHAPE);GY:回转半径,Radius of gyration (GYRATE);AREA:平均斑块面积,Patch area;PARA:周长面积比,Perimeter-area ratio
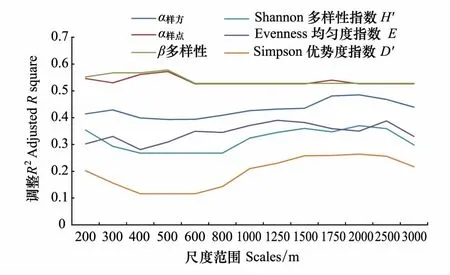
图4 景观异质性指数和群落结构指标对生物多样性特征指标的解释力Fig.4 The explanatory power of landscape heterogeneity and community structure on biodiversity
4 讨论
4.1 农业景观中非农生境在生物多样性保护和生态系统服务中的作用
尽管人类采取了多样的生物多样性保护计划,但全球生物多样性下降的趋势仍在持续[29-30],而农业景观在生物多样性保护中具有重要作用[31-32],农业景观中残存的自然、半自然斑块及其上存在的植物群落对农药、化肥、降水等具有过滤、缓冲和沉降作用,成为众多物种的食源地、栖息地、庇护所等,对区域生物多样性保护和生态系统服务维持发挥着举足轻重的作用[33-35]。本研究的结果表明:次生灌丛等自然生境中的总物种丰富度显著高于其他生境。次生林地作为另一种自然生境,其内的灌草层物种丰富度要低于其他几类生境,受多方面原因的影响[36],一方面自然林地生境中在局地尺度上物种丰富度的比例要明显高于其他生境(图3,表3,α样方在γ中的比例达14.6%),同时由于生境异质性的下降而使区域间物种多样性的差异(β2)在总物种丰富度(γ)中的比重迅速下降;另一方面由于研究区内的次生林地多处于坡度大、难以调查的区域内,本次取样点的数量相较于其他生境数量明显不足也在一定程度上影响了其物种丰富度的大小。农田边缘生境是农业景观中的一种特有生境,多处于田块间或梯田边坡位置,呈条带状分布,宽度一般不超过1m,但该生境中保有的物种丰富度却仅次于次生灌丛生境,成为区域内植物多样性保护的重要场所,其他研究也发现该类生境在区域生物多样性保护和生态系统服务中也具有非常重要的作用[15,37-39]等。而人工林这种半自然生境在区域植物多样性保护方面并未能发挥出更高的效应[40]。
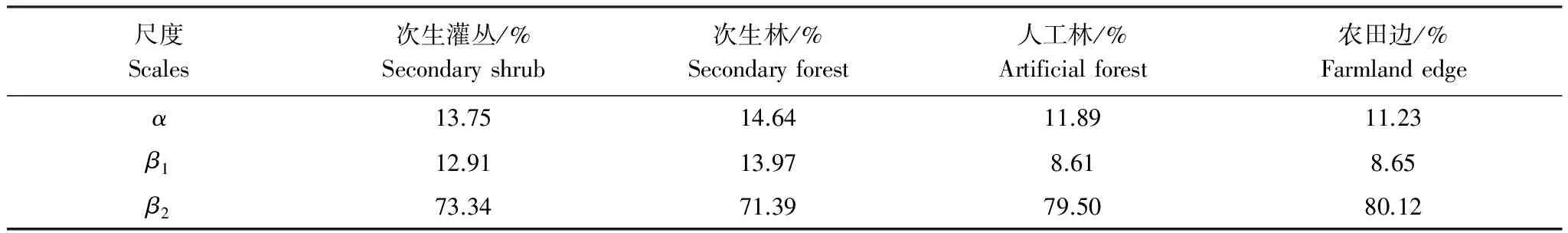
表3 不同生境中各尺度物种丰富度对总物种丰富度的贡献度
4.2 景观异质性和群落结构特征对植物物种多样性影响的相对重要性
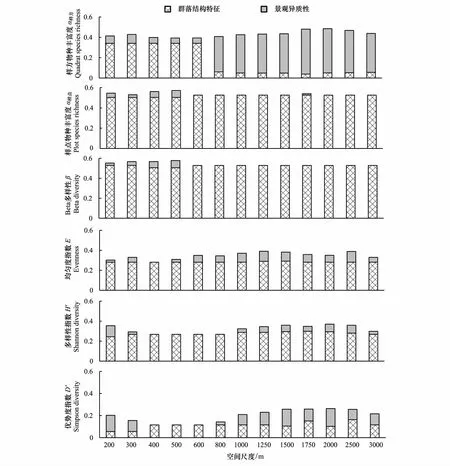
图5 多元线性回归模型总体解释力(调整R2)中群落结构特征和景观异质性指标的分解Fig.5 The variance decomposition of landscape heterogeneity and community structure in the multiple linear regression models
研究发现,农业景观中,景观异质性变化会对区域内的生物多样性产生重要影响[33,41-42],但不同生物类群的响应和不同景观异质性特征的影响也存在着差异,并表现出强烈的尺度效应,波动强烈[18- 19,43-44]。从本研究结果来看(图4),景观格局异质性和植物群落结构对植物多样性特征不同指数的综合解释力存在一定的差异,并呈现出一定的尺度效应。随着空间尺度的增大,模型对多个多样性指数的解释力呈先下降后上升的态势。研究者认为原因如下:从回归模型中各指标对模型总体解释力的贡献来看(图5),总体上群落结构特征指标对各植物多样性指数的解释力随空间尺度变化而较稳定,但在小尺度上(300m以下),表征景观异质性特征的自然、半自然斑块面积比例(PLAND)对于样方尺度的物种丰富度具有极高的解释力,二者叠加导致模型总体解释力较高;随着空间尺度增大(300—800m),景观异质性指标的解释力较弱,因而在该尺度范围内,模型总体解释力最低;随空间尺度的进一步增大(800m以上),景观异质性指数的解释力在迅速上升,因而模型解释力在上升,而空间尺度超过2500m之后,景观异质性指数的解释力开始下降,模型总体解释力进而呈下降态势本研究还发现,群落结构特征对生物多样性的影响更为直接和强烈,区域内其他研究也证实在农业景观中林地蜘蛛种群物种多度主要受林下草本层高度和盖度的影响,而对各景观异质性指数的响应较弱[45]。而关于景观异质性与植物物种多样性间的关系研究发现二者关系均较弱,且表现出强烈的尺度波动性,缺乏一致性[46-47],这种强烈的尺度依赖性和似然性,很可能是由于所用的这些景观异质性指数很多没有明确的生态学意义[48],而传统的生物多样性指数也都是基于统计学得出,因而这种相关关系的得出很可能仅是统计上的相关而非生态学上的。
从表征植物多样性特征的不同指数来看,以往常用的物种多样性指数,如Shannon多样性指数(H′)、均匀度指数(E)和优势度指数(D′)等,并不能很好的被解释,而对拆分后的物种丰富度指数有更好的解释力。因而通过对物种丰富度拆分之后进行探讨景观格局变化、人类活动等对生物多样性的影响研究会更有有利,另一方面物种丰富度的加性分配因其度量单位一致,其生态学意义会更容易理解和解释,能有效的用于比较不同时空尺度、人类活动强度和景观背景中的物种多样性差异分析[49- 50]。
4.3 景观异质性对区域植物多样性影响的重要性
前述结果中显示景观异质性对植物多样性的影响较弱,但并不能说景观异质性对植物多性的影响不大、不重要,相反的,景观异质性变化对区域生物多样性保护具有举足轻重的作用。
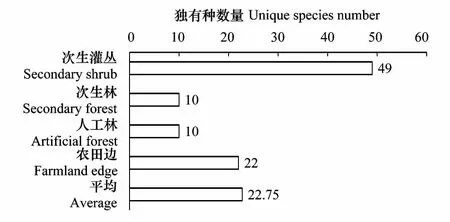
图6 不同生境中独有种数量Fig.6 The unique species number in different habitats
景观异质性的变化会通过景观组成要素变化(景观斑块类型丰富度的增减)而影响到区域植物物种丰富度和多样性的变化,本研究的数据显示(图6),各类生境中均有一定量的独有种存在,而每增加一个新的生境类别(景观要素),平均会有22.75个新的物种出现,因而景观组成的变化会迅即影响到区域总物种丰富度的变化。前人研究发现景观构型与景观组成对生物多样性的影响也存在着差异[43,51-53],但本研究目前已有的数据对该问题尚无法给出令人信服的结论,但推测在景观组成一定的情况下,随着景观构型的复杂化,一些偶见种将会有更大的概率能够占据一定的生态位,从而促进区域物种丰富度的提高,但该效应同时会因一些大生境种(对最小生境斑块要求高的物种)的消失而降低。
还有研究发现,景观异质性特征对植物群落和物种多样性的影响表现出一定的“时滞”效应[54-57],并因不同生物类群“灭绝债务”的不同而存在差异[58-59],因而在区域生物多样性保护过程中,景观异质性的重要性仍不容忽视。
5 结论和建议
在农业景观中,植物群落结构特征等局地环境因子与植物群落物种多样性的关系更为直接和密切,但随着空间尺度的增大,景观异质性作用将会逐渐增强,成为影响植物群落物种多样性及其组成结构的潜在和间接因素,并通过局地生态因子的变化进而影响到生物多样性的变化,而生物多样性的变化常常会通过beta多样性的变化展现出来。因而在未来持续农业景观构建过程中,生物多样性保护和生态系统服务的维持在小尺度上需要重视非农生境中植物群落结构的作用,而在大尺度上,景观组成和构型异质性的提高(同时还要考虑景观破碎化的负效应)具有不可忽视的重要作用,从而从区域上能够进一步提高生物多样性保护和生态系统服务的持续供给。
致谢:感谢董翠芳、张晓阳、王润、卞子亓、王美娜、任嘉衍等同学对野外实验调查的帮助。
参考文献(References):
[1] Foley J A,Ramankutty N,Brauman K A,Cassidy E S,Gerber J S,Johnston M,Mueller N D,O′Connell C,Ray D K,West P C,Balzer C,Bennett E M,Carpenter S R,Hill J,Monfreda C,Polasky S,Rockström J,Sheehan J,Siebert S,Tilman D,Zaks D P M. Solutions for a cultivated planet. Nature,2011,478(7369): 337- 342.
[2] Barnosky A D,Brown J H,Daily G C,Dirzo R,Ehrlich A H,Ehrlich P R,Eronen J T,Fortelius M,Hadly E A,Leopold E B,Mooney H A,Myers J P,Naylor R L,Palumbi S,Stenseth N C,Wake M H. Maintaining humanity′s life support systems in the 21st century. 2013. http://consensusforaction.stanford.edu/see-scientific-consensus/consensus_english.pdf.
[3] Food and Agriculture Organization of the United Nations. FAO statistical yearbook 2013 world food and agriculture. United Nations. Rome: FAO,2013.
[4] Foley J A,DeFries R,Asner G P,Barford C,Bonan G,Carpenter S R,Chapin F S,Coe M T,Daily G C,Gibbs H K,Helkowski J H,Holloway T,Howard E A,Kucharik C J,Monfreda C,Patz J A,Prentice I C,Ramankutty N,Snyder P K. Global consequences of land use. Science,2005,309(5734): 570- 574.
[5] Godfray H C J,Beddington J R,Crute I R,Haddad L,Lawrence D,Muir J F,Pretty J,Robinson S,Thomas S M,Toulmin C. Food security: the challenge of feeding 9 billion people. Science,2010,327(5967): 812- 818.
[6] Meehan T D,Werling B P,Landis D A,Gratton C. Agricultural landscape simplification and insecticide use in the Midwestern United States. Proceedings of the National Academy of Sciences of the United States of America,2011,108(28): 11500- 11505.
[7] Cumming G S,Buerkert A,Hoffmann E M,Schlecht E,von Cramon-Taubadel S,Tscharntke T. Implications of agricultural transitions and urbanization for ecosystem services. Nature,2014,515(7525): 50- 57.
[8] Power A G. Ecosystem services and agriculture: tradeoffs and synergies. Philosophical Transactions of The Royal Society B: Biological Sciences,2010,365(1554): 2959- 2971.
[9] Tilman D,Balzer C,Hill J,Befort B L. Global food demand and the sustainable intensification of agriculture. Proceedings of the National Academy of Sciences of the United States of America,2011,108(50): 20260- 20264.
[10] Bright J A,Morris A J,Field R H,Cooke A I,Grice P V,Walker L K,Fern J,Peach W J.Higher-tier agri-environment scheme enhances breeding densities of some priority farmland birds in England. Agriculture,Ecosystems & Environment,2015,203: 69- 79.
[11] Wretenberg J,Pärt T,Berg Å. Changes in local species richness of farmland birds in relation to land-use changes and landscape structure. Biological Conservation,2010,143(2): 375- 381.
[12] Rundlöf M,Edlund M,Smith H G. Organic farming at local and landscape scales benefits plant diversity. Ecography,2010,33(3): 514- 522.
[13] Crowder D W,Northfield T D,Strand M R,Snyder W E. Organic agriculture promotes evenness and natural pest control. Nature,2010,466(7302): 109- 112.
[14] Holland J M,Oaten H,Moreby S,Birkett T,Simper J,Southway S,Smith B M. Agri-environment scheme enhancing ecosystem services: A demonstration of improved biological control in cereal crops. Agriculture,Ecosystems & Environment,2012,155: 147- 152.
[15] D′Acunto L,Semmartin M,Ghersa C M. Uncultivated margins are source of soil microbial diversity in an agricultural landscape. Agriculture,Ecosystems & Environment,2016,220: 1- 7.
[16] Flohre A,Rudnick M,Traser G,Tscharntke T,Eggers T. Does soil biota benefit from organic farming in complex vs. simple landscapes? Agriculture,Ecosystems & Environment,2011,141(1/2): 210- 214.
[17] Scherber C,Eisenhauer N,Weisser W W,Schmid B,Voigt W,Fischer M,Schulze E D,Roscher C,Weigelt A,Allan E,Beßler H,Bonkowski M,Buchmann N,Buscot F,Clement L W,Ebeling A,Engels C,Halle S,Kertscher I,Klein A M,Koller R,König S,Kowalski E,Kummer V,Kuu A,Lange M,Lauterbach D,Middelhoff C,Migunova V D,Milcu A,Müller R,Partsch S,Petermann J S,Renker C,Rottstock T,Sabais A,Scheu S,Schumacher J,Temperton V M,Tscharntke T. Bottom-up effects of plant diversity on multitrophic interactions in a biodiversity experiment. Nature,2010,468(7323): 553- 556.
[18] Gabriel D,Sait S M,Hodgson J A,Schmutz U,Kunin W E,Benton T G. Scale matters: the impact of organic farming on biodiversity at different spatial scales. Ecology Letters,2010,13(7): 858- 869.
[19] Sutcliffe L M E,Batáry P,Becker T,Orci K M,Leuschner C. Both local and landscape factors determine plant and Orthoptera diversity in the semi-natural grasslands of Transylvania,Romania. Biodiversity and Conservation,2015,24(2): 229- 245.
[20] Wu J G. Key concepts and research topics in landscape ecology revisited: 30 years after the Allerton Park workshop. Landscape Ecology,2013,28(1): 1- 11.
[21] Bennett A B,Gratton C. Local and landscape scale variables impact parasitoid assemblages across an urbanization gradient. Landscape and Urban Planning,2012,104(1): 26- 33
[22] Wu J G,Buyantuyev A,Jenerette G D,Litteral J,Neil K,Shen W J. Quantifying spatiotemporal patterns and ecological effects of urbanization: a multiscale landscape approach//Richter M,Weiland U,eds. Applied Urban Ecology: A Global Framework. Chichester,UK: John Wiley & Sons,Ltd,2011: 33- 53.
[23] Garden J G,Mcalpine C A,Possingham H P. Multi-scaled habitat considerations for conserving urban biodiversity: native reptiles and small mammals in Brisbane,Australia. Landscape Ecology,2010,25(7): 1013- 1028.
[24] Sutherland W J,Freckleton R P,Godfray H C J,Beissinger S R,Benton T,Cameron D D,Carmel Y,Coomes D A,Coulson T,Emmerson M C,Hails R S,Hays G C,Hodgson D J,Hutchings M J,Johnson D,Jones J P G,Keeling M J,Kokko H,Kunin W E,Lanbin X,Lewis O T,Malhi Y,Mieszkowska N,Milner-Gulland E J,Norris K,Phillimore A B,Purves D W,Reid J M,Reuman D C,Thompson K,Travis J M J,Turnbull L A,Wardle D A,Wiegand T. Identification of 100 fundamental ecological questions. Journal of Ecology,2013,101(1): 58- 67.
[25] Massol F,Gravel D,Mouquet N,Cadotte M W,Fukami T,Leibold M A. Linking community and ecosystem dynamics through spatial ecology. Ecology letters,2011,14(3): 313- 323.
[26] Colwell R K. EstimateS: Statistical estimation of species richness and shared species from samples. Version 9. 2013. http://viceroy.eeb.uconn.edu/EstimateS/.
[27] Colwell R K,Chao A,Gotelli N J,Lin S Y,Mao C X,Chazdon R L,Longino J T. Models and estimators linking individual-based and sample-based rarefaction,extrapolation and comparison of assemblages. Journal of Plant Ecology,2012,5(1): 3- 21.
[28] Veech J A,Crist T O. PARTITION: software for hierarchical partitioning of species diversity,version 3.0. 2009. http://www.users.muohio.edu/cristto/partition.htm.
[29] Perrings C,Naeem S,Ahrestani F,Bunker D E,Burkill P,Canziani G,Elmqvist T,Ferrati R,Fuhrman J,Jaksic F,Kawabata Z,Kinzig A,Mace G M,Milano F,Mooney H,Prieur-Richard A H,Tschirhart J,Weisser W. Ecosystem services for 2020. Science,2010,330(6002): 323- 324.
[30] Perrings C,Naeem S,Ahrestani F S,Bunker D E,Burkill P,Canziani G,Elmqvist T,Fuhrman J A,Jaksic F M,Kawabata Z,Kinzig A,Mace G M,Mooney H,Prieur-Richard A H,Tschirhart J,Weisser W. Ecosystem services,targets,and indicators for the conservation and sustainable use of biodiversity. Frontiers in Ecology and the Environment,2011,9(9): 512- 520.
[31] Rodrigues A S L,Andelman S J,Bakarr M I,Boitani L,Brooks T M,Cowling R M,Fishpool L D C,da Fonseca G A B,Gaston K J,Hoffmann M,Long J S,Marquet P A,Pilgrim J D,Pressey R L,Schipper J,Sechrest W,Stuart S N,Underhill L G,Waller R W,Watts M E J,Yan X. Effectiveness of the global protected area network in representing species diversity. Nature,2004,428(6983): 640- 643.
[32] Fischer J,Lindenmayer D B,Manning A D. Biodiversity,ecosystem function,and resilience: ten guiding principles for commodity production landscapes. Frontiers in Ecology and the Environment,2006,4(2): 80- 86.
[33] Tscharntke T,Klein A M,Kruess A,Steffan-Dewenter I,Thies C. Landscape perspectives on agricultural intensification and biodiversity-ecosystem service management. Ecology Letters,2005,8(8): 857- 874.
[34] Van Buskirk J,Willi Y. Enhancement of farmland biodiversity within set-aside land. Conservation Biology,2004,18(4): 987- 994.
[35] Broughton R K,Shore R F,Heard M S,Amy S R,Meek W R,Redhead J W,Turk A,Pywell R F. Agri-environment scheme enhances small mammal diversity and abundance at the farm-scale. Agriculture,Ecosystems & Environment,2014,192: 122- 129.
[36] 卢训令,胡楠,丁圣彦,范玉龙,廖秉华,翟元杰,柳静. 伏牛山自然保护区物种多样性分布格局. 生态学报,2010,30(21): 5790- 5798.
[37] Morandin L A,Long R F,Kremen C. Hedgerows enhance beneficial insects on adjacent tomato fields in an intensive agricultural landscape. Agriculture,Ecosystems & Environment,2014,189: 164- 170.
[38] Dainese M,Montecchiari S,Sitzia T,Sigura M,Marini L. High cover of hedgerows in the landscape supports multiple ecosystem services in Mediterranean cereal fields. Journal of Applied Ecology,2017,54(2): 380- 388.
[39] Lecq S,Loisel A,Brischoux F,Mullin S J,Bonnet X. Importance of ground refuges for the biodiversity in agricultural hedgerows. Ecological Indicators,2017,72: 615- 626.
[40] 卢训令,汤茜,梁国付,丁圣彦. 黄河下游平原不同非农生境中植物多样性. 生态学报,2015,35(5): 1527- 1536.
[41] Tscharntke T,Tylianakis J M,Rand T A,Didham R K,Fahrig L,Batáry P,Bengtsson J,Clough Y,Crist T O,Dormann C F,Ewers R M,Fründ J,Holt R D,Holzschuh A,Klein A M,Kleijn D,Kremen C,Landis D A,Laurance W,Lindenmayer D,Scherber C,Sodhi N,Steffan-Dewenter I,Thies C,van der Putten W H,Westphal C. Landscape moderation of biodiversity patterns and processes-eight hypotheses. Biological Reviews,2012,87(3):661- 685.
[42] Puech C,Poggi S,Baudry J,Aviron S. Do farming practices affect natural enemies at the landscape scale? Landscape ecology,2015,30(1): 125- 140.
[43] Steckel J,Westphal C,Peters M K,Bellach M,Rothenwoehrer C,Erasmi S,Scherber C,Tscharntke T,Steffan-Dewenter I. Landscape composition and configuration differently affect trap-nesting bees,wasps and their antagonists. Biological Conservation,2014,172: 56- 64.
[45] 赵爽,宋博,丁圣彦,侯笑云,刘晓博,汤茜,王润. 黄河下游农业景观中景观结构和生境特征对林表生蜘蛛多样性的影响. 生态学报,2017,37(6): 1816-1825.
[46] 董翠芳,梁国付,丁圣彦,卢训令,汤茜,李栋科. 不同干扰背景下景观指数与物种多样性的多尺度效应——以巩义市为例. 生态学报,2014,34(12): 3444- 3451.
[47] 王玉婷,丁圣彦,梁国付. 农田背景下景观结构对半自然生境生物多样性的多尺度影响. 地理科学进展,2014,33(12): 1704- 1716.
[48] 陈利顶,丘君,张淑荣,傅伯杰. 复杂景观中营养型非点源污染物时空变异特征分析. 环境科学,2003,24(3):85- 90.
[49] Crist T O,Veech J A. Aditive partitioning of rarefaction curves and species-area relationships: unifying α-,β- and γ-diversity with sample size and habitat area. Ecology Letters,2006,9(8): 923- 932.
[50] 陈圣宾,欧阳志云,徐卫华,肖燚. Beta多样性研究进展. 生物多样性,2010,18(4): 323- 335.
[51] Haenke S,Kovács‐Hostyánszki A,Fründ J,Batáry P,Jauker B,Tscharntke T,Holzschuh A. Landscape configuration of crops and hedgerows drives local syrphid fly abundance. Journal of Applied Ecology,2014,51(2): 505- 513.
[52] Arroyo-Rodríguez V,Rojas C,Saldaa-Vázquez R A,Stoner K E. Landscape composition is more important than landscape configuration for phyllostomid bat assemblages in a fragmented biodiversity hotspot. Biological Conservation,2016,198: 84- 92.
[53] Matos F A R,Magnago L F S,Gastauer M,Carreiras J M B,Simonelli M,Meira-Neto J A A,Edwards D P. Effects of landscape configuration and composition on phylogenetic diversity of trees in a highly fragmented tropical forest. Journal of Ecology,2017,105(1): 265- 276.
[54] Lindborg R,Eriksson O. Historical landscape connectivity affects present plant species diversity. Ecology,2004,85(7): 1840- 1845.
[55] Helm A,Hanski I,Pärtel M. Slow response of plant species richness to habitat loss and fragmentation. Ecology Letters,2006,9(1): 72- 77.
[56] Krauss J,Bommarco R,Guardiola M,Heikkinen R K,Helm A,Kuussaari M,Lindborg R,Öckinger E,Pärtel M,Pino J,Pöyry J,Raatikainen K M,Sang A,Stefanescu C,Teder T,Zobel M,Steffan-Dewenter I. Habitat fragmentation causes immediate and time-delayed biodiversity loss at different trophic levels. Ecology Letters,2010,13(5): 597- 605.
[57] Murphy S J,Audino L D,Whitacre J,Eck J L,Wenzel J W,Queenborough S A,Comita L S. Species associations structured by environment and land-use history promote beta-diversity in a temperate forest. Ecology,2015,96(3): 705- 715.
[58] Tilman D,May R M,Lehman C L,Nowak M A. Habitat destruction and the extinction debt. Nature,1994,371(6492): 65- 66
[59] Halley J M,Monokrousos N,Mazaris A D,Newmark W D,Vokou D. Dynamics of extinction debt across five taxonomic groups. Nature Communications,2016,7: 12283.
猜你喜欢
湖北植保(2022年4期)2022-08-23
林业勘查设计(2022年1期)2022-02-15
乡村科技(2021年17期)2021-10-20
安顺学院学报(2021年4期)2021-09-16
土壤(2021年1期)2021-03-23
落叶果树(2021年6期)2021-02-12
林业调查规划(2020年3期)2020-06-03
土壤学报(2017年5期)2017-11-01
福建农业学报(2016年6期)2016-11-01
体育科技(2016年2期)2016-02-28
