
持续性环境热应激对公羊血液生化指标、生殖激素及精液品质的影响
2018-04-12 10:41:40施力光彭维祺胡海超侯冠彧周汉林
家畜生态学报 2018年3期
施力光,彭维祺,胡海超,侯冠彧,周汉林
(中国热带农业科学院 热带作物品种资源研究所,海南 儋州 571737)
热应激是当机体或细胞处于高温环境中表现出的一系列非特异性生理反应的总和[1],表现为畜禽呼吸频率加快、脱水、采食量下降、精神委靡、神经机能紊乱等,而且机体热应激的严重程度随温度和湿度的增高而加重,重度热应激甚至导致畜禽死亡,给养殖生产造成损失[2]。尽管热应激的作用机理至今尚未完全探明,但已有研究证实热应激能通过交感-肾上腺髓质系统和下丘脑-垂体-肾上腺轴引起代谢、免疫及生殖机能紊乱。血液生化值是反映机体代谢状况最敏感的指标之一,Cwynar等[3]测定了美利奴羊种公羊在不同温度下(16.5 ℃、30 ℃和50 ℃)的血液指标生理化指标,测得血液红细胞数、葡萄糖、肾上腺素等血液生理化指标在随着环境温度变化出现显著改变;雷虹等[4]通过研究湘西黄牛在夏季热应激状态下(THI>82)18项血液生理化指标,发现夏季高温会造成热应激,而且对牛血液生理指标影响较大,对血液生化参数影响不明显;姜润深等[5]通过研究热应激初期肉鸡血液生化指标与耐热性能的关系,发现热应激状态鸡血清葡萄糖浓度和免疫球蛋白含量出现显著变化。可见通过畜禽血液生化指标变化能较直观的反映畜禽机体热应激状况。热应激还对公畜繁殖性能的影响尤为显著,热应激发生时不仅造成种公畜呼吸和心率加快导致血液循环加快,出现呼吸性碱中毒症状;同时会导致下丘脑促性腺激素释放激素(CnRH)和垂体前叶促性腺激素的分泌减少,影响性欲和交配能力[6-7]。Hamilton[8]通过热应激环境下公羊睾丸和附睾精液品质测定及氧化还原酶系活性证实,热应激造成的精子不可逆转的损伤。本课题组前期研究显示,海南地区夏季高温湿热持续强度大、时间长,环境温度持续在30 ℃左右的时间长达4~5个月之久[9],对畜禽影响极为明显。
本试验通过测定海南高温湿热气候条件下种公羊血液生化指标、血清生殖激素和精液品质等相关指标的变化,探讨种公羊对热应激的生理响应,旨在为我国南方热区养羊生产缓解热应激提供参考。
1 材料与方法
1.1 试验动物与饲养管理
本试验用种公羊由中国热带农业科学院热带作物品种资源研究所海南黑山羊种羊场提供,共15只,体重32~48 kg,年龄2~4岁,体质健康。试验羊舍为半开放高床羊舍,试验分2个阶段进行,即春季(非应激期)(2017年3月10日~2017年3月30日)和夏季(持续性热应激期)(2017年7月10日~2017年7月30日)。试验期间日常管理相同,专人负责定期投喂,自由饮水,定时定点测定羊舍温、湿度,参照奶牛环境温湿度指数(temperature humidity index,THI)计算方法[10],用以评价炎热程度。试验前统一驱虫并进行常规免疫。
1.2 样品采集
试验期间,分别对非热应激期和持续性热应激期的种公羊采用颈静脉采血方法进行空腹采血10 mL,每5天采集一次,其中5 mL EDTA抗凝处理,4 ℃保存待测生理指标;另5 mL室温静置0.5~1 h,4 ℃ 3 000~3 500 rpm离心20 min分离血清,待测激素和生化指标。每次采血结束后,采用电刺激法采集种公羊精液,现场常规方法分别进行精液品质检测。
1.3 血清生化指标分析
血清谷丙转氨酶(ALT)、谷草转氨酶(AST)、总胆固醇(CHO)、葡萄糖(Glu)等生化指标委托海南西部中心医院通过美国贝克曼全自动生化分析仪进行检测。促卵泡素(FSH)、促黄体素(LH)、睾酮(T)等血清生殖激素采用ELISA法测定(ELISA检测试剂盒购自上海哈灵生物有限公司),具体操作步骤参照试剂盒提供使用说明书进行。
1.4 精液品质分析
射精量采集到的精液直接用集精杯测量射精体积。精子活力用预温生理盐水稀释精液100倍,混匀后滴在载玻片上,盖上洁净的盖玻片。置于显微镜下观察(放大倍数250至400倍左右)。精子活力=呈直线前进运动精子数/总精子数×100%。精子密度测定用预温生理盐水按1∶100比例稀释精液,利用血细胞计数板计数测定。每毫升精子数量=5个中方格数的精子数×5×10×1000×100(稀释倍数)。畸形率的测定用姬姆萨染色法检测,取稀释后精液样品1滴制作抹片,4%多聚甲醛固定10 min,姬姆萨染色1 h,风干后镜检计数。
1.5 数据处理与统计分析
试验数据采用SPSS 17.0软件进行分析,应激期和非应激期2阶段组间平均数比较采用独立样本t检验,结果以平均值±标准差(Mean±SD)表示,P<0.01为差异极显著,P<0.05为差异显著。
2 结果与分析
2.1 不同试验期羊舍环境参数监测
试验羊舍温湿度指标监测结果如表1所示,在非应激期羊舍THI为70~72,试验动物无热应激或短暂性热应激;在热应激期羊舍平均THI达84以上,试验公羊遭受持续性强热应激。

表1 试验羊舍温湿度指标Table 1 Temperature and humidity index of goat house
2.2 持续性热应激对公羊血液生理指标的影响
由表2可知,持续性热应激造成海南黑山羊种公羊血液中的红细胞数显著增加,白细胞数量显著降低(P<0.05),而对血小板数和淋巴细胞数影响不大。在应激环境中种公羊血液中血红蛋白浓度、红细胞比容和平均血红蛋白含量的影响差异显著(P<0.05)。
2.3 持续性热应激对公羊血液生化指标的影响
本试验测定了包括不同环境下种公羊血液中葡萄糖(GLU)、谷丙转氨酶(ALT)、碱性磷酸酶(ALP和免疫球蛋白等12项血液生化指标,结果如表3所示。由表3可知,持续性的环境热应激对血清免疫球蛋白浓度影响显著,极显著降低IgM和IgA浓度(P<0.01),IgG浓度也显著低于非应激期水平(P<0.05);持续性的环境热应激显著增加血清中碱性磷酸酶(ALP)和乳酸脱氢酶(LDH)酶活性(P<0.05),对血清中甘油三酯浓度也有极显著影响(P<0.01);对其他生化指标影响差异不显著(P>0.05)。
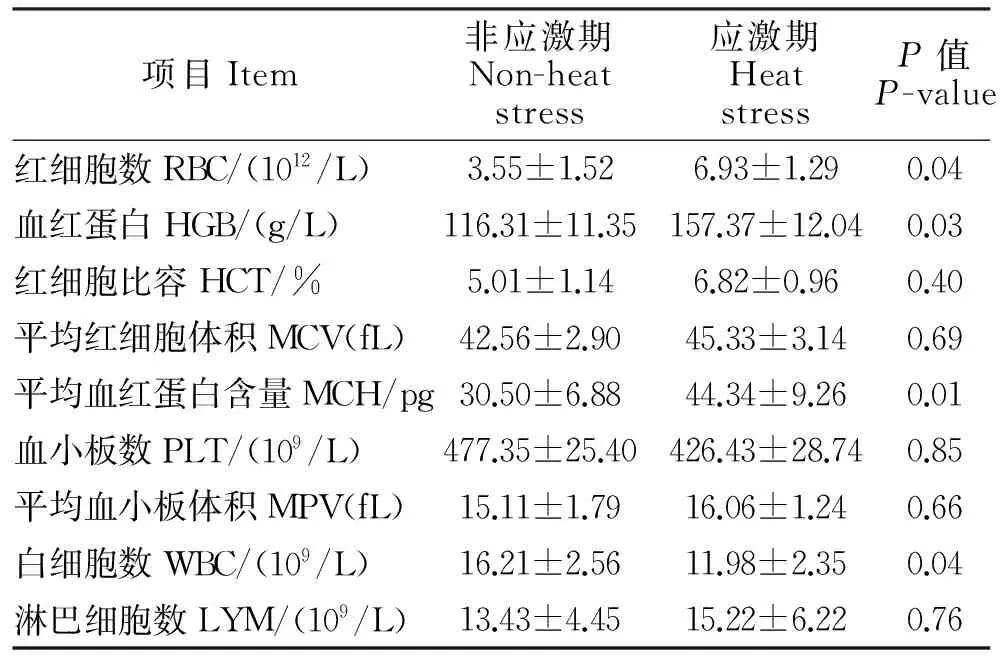
表2 血液生理指标测定结果Table 2 Results of blood physiology index
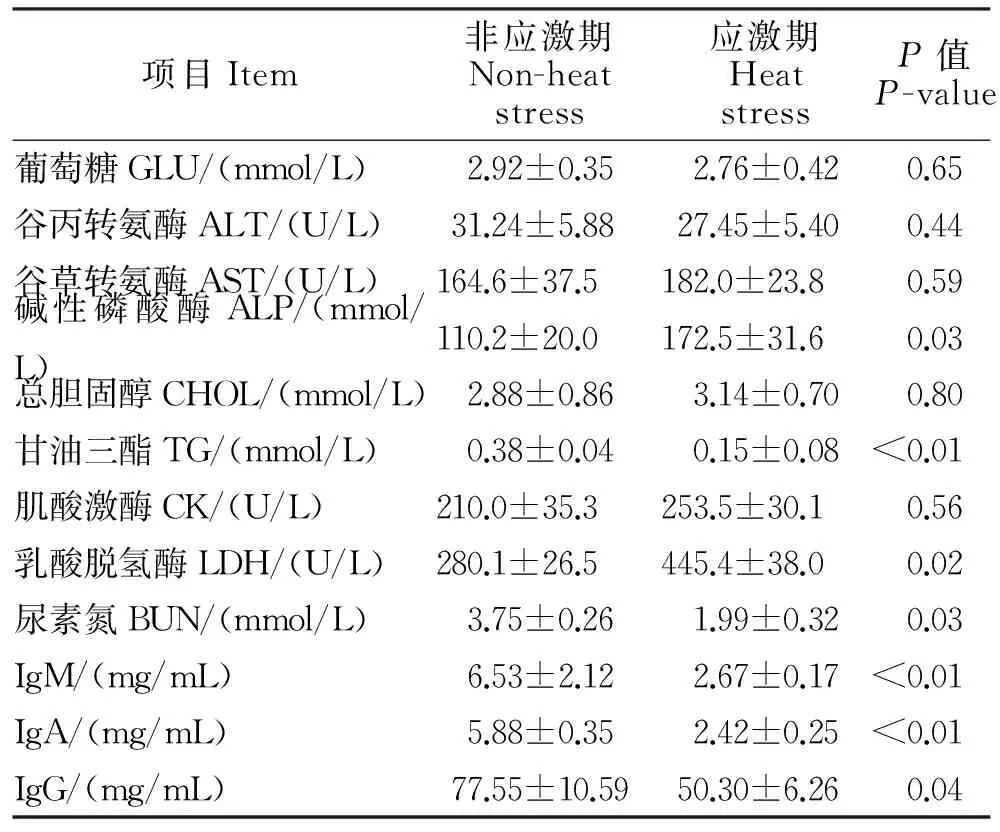
表3 血液生化指标测定结果Table 3 Results of blood biochemical indexes
2.4 持续性热应激对公羊血清生殖激素的影响
公羊血清生殖激素ELISA测定结果见表4。从表4可见,持续性的环境热应激对公羊血清生殖激素水平影响差异显著。持续性的热应激极显著降低了血清FSH和LH含量,血清睾酮T浓度也显著低于非应激期(P<0.05)。
2.5 持续性热应激对公羊精液品质的影响
由表5可知,在应激期和非应激期种公羊采精量的无显著差异(P>0.05),但持续性的环境热应激能显著降低精子密度(P<0.05),而且精子活力也受到环境热应激的影响,降低22%(P<0.01);同时持续性的热应激还极显著的使种公羊精子畸形率增加了50%以上(P<0.01)。
3 讨 论
3.1 持续性热应激对海南黑山羊种公羊血液生理指标的影响
畜禽血液生理生化指标作为反应机体代谢情况和应激状态的一组参数,对评价畜禽机体生理状况具有重要意义。因为机体对外界环境的应激,会直接引起内环境相应的调节,造成血液成分的改变和代谢的调节。从本试验检测的21项血液生理生化指标可以看出,持续性的高温环境对公羊血液细胞成分中RBC、HCT、HGB和MCH有显著影响。因为山羊汗腺不发达,环境高温时只能依靠加快呼吸频率进行辅助散热的代偿反应进行自身恒温的调节,呼吸频率的加快迫使红细胞数和血红蛋白含量增加以供给机体耗氧量的需求,这与Cwynar等[3]、雷虹等[4]、赵宗胜等[11]、黄光远等[12]测定的热应激环境下绵羊、肉牛、奶牛、成年兔血液生理指标变化结果基本一致。陶林等[13]报道,高温应激对公鸡的生理指标没有显著影响,推测可能是由于地方品种对慢性的热应激有较强的适应性和耐受力。关于热应激对畜禽血液白细胞数WBC影响的相关研究结果不尽一致,有研究显示环境热应激能够显著提高畜禽血液中白细胞数(WBC)[14];另有相关研究报道热应激对WBC影响差异不显著,人长期桑拿浴(高温应激)对外周白细胞数没有显著影响,也没有白细胞产生堆积效应[15],这可能是机体适应了类似代谢热应激产生的自我调控机制。本试验中种公羊在持续性环境热应激后WBC显著减少,与雷虹等[4]报道结果相符。结合血清免疫球蛋白测定结果,推测可能是由于在热应激条件下种公羊的免疫力有所下降所致。
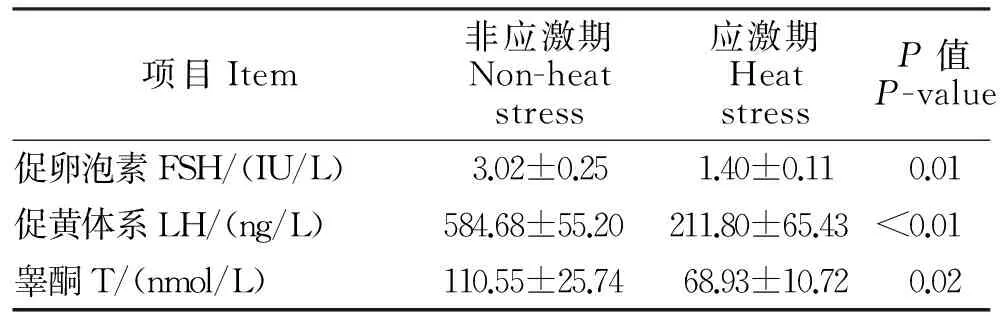
表4 血清生殖激素测定结果Table 4 Results of serum reproductive hormone
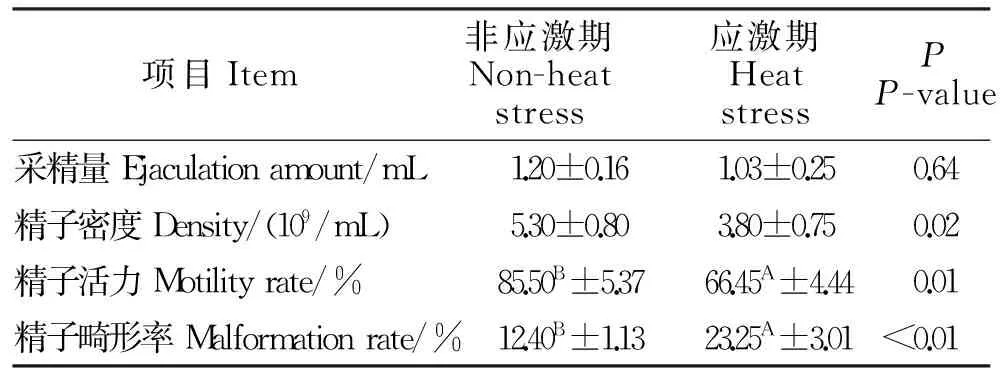
表5 精液品质测定结果Table 5 Results of semen quality
3.2 持续性热应激对海南黑山羊种公羊血液生化指标的影响
血糖是反映机体营养代谢状况最直观的指标之一,也是能反映动物应激的指标之一,因为热应激时能引起肾上腺交感神经兴奋,使肾上腺素分泌增强,导致GLU含量升高[16]。白丹丹等[17]研究显示,慢性热应激导致肉牛血清GLU含量显著降低,而Scharf等[18]报道急性热应激时肉牛血清中GLU升高,也有相关高温应激对公鸡血清GLU含量无显著影响的研究报道[19]。本试验中,持续性的热应激对种公羊血清GLU含量影响不显著,而BUN和TG在热应激阶段显著降低,可能是因为持续性的热应激引起肾上腺交感神经持续性的作用,使代谢向降解蛋白质、提高糖原合成和脂肪沉积方向进行,引起BUN和TG含量的变化。可见,持续性热应激对血液中糖、蛋白及脂肪的代谢有影响。
热应激导致机体代谢系统受到干扰,势必会造成血清中酶的活性和含量发生变化。本试验发现高温应激会引起ALP和LDH显著升高,因为热应激导致细胞膜的通透性发生改变,能量代谢的无氧酵解过程明显增强,引起LDH活性升高,与白丹丹等[17]、Hocking等[19]报道结果一致。本试验检测到ALT、AST和CK活性未受到持续性热应激显著影响,鉴于目前关于热应激反应中血清酶的变化报道不一,推测可能是品种差异导致的。
热应激状态下机体的免疫机能都会减弱[20-21]。本研究测定的3种免疫球蛋白在持续性热应激条件下含量显著降低,其中IgM和IgA分别作为体液免疫应答中最先产生的免疫球蛋白和介导调理吞噬作用的免疫球蛋白,其含量极显著减少,杀菌、溶菌和促吞噬能力下降,导致种羊的免疫系统效率降低。
3.3 持续性热应激对海南黑山羊种公羊血清生殖激素和精液品质的影响
生殖激素对种公畜繁殖性能有非常重要的调控作用,机体通过下丘脑-垂体-性腺轴调节生殖激素的分泌,因此血清生殖激素含量是反映种公畜生殖机能的重要指标之一。高温应激能引起肾上腺交感神经兴奋,调节机体代谢机能;Maurya等[22]研究证实,高温通过下丘脑-垂体-性腺轴影响畜禽的生殖激素的分泌。本试验结果表明,在持续性热应激阶段,种公羊血清FSH和LH水平极显著的降低,因为环境热应激导致的内分泌紊乱会抑制下丘脑的活动,会减少GnRH的分泌,进而影响垂体FSH和LH的分泌减少。在雄性动物中LH分泌的减少又会导致睾酮T合成降低,与杨明等[23]、Pei等[24]结果一致。本试验中除种羊射精量未受影响外,热应激致使公羊精子活力和密度显著下降,精子畸形率极显著升高。也是由于LH分泌减少,睾酮合成降低,抑制精子细胞的进一步正常发育,使精子细胞的增殖分裂和发育受阻,导致精子密度降低所致。与此同时,热应激除了造成激素分泌的影响,还有代谢紊乱积累的有毒有害物质增多,破坏精子的形态结构和功能,降低精子的活率并增加畸形精子数。重度热应激会损害精子的DNA,影响受精过程。
4 结 论
持续性的热应激可导致海南黑山羊种公羊血液RBC、HGB、MBC、TG和LDH等生理生化指标的显著改变,造成了血清生殖激素水平的显著降低和精液品质的下降,血清免疫球蛋白水平的显著降低反映热应激对还可能造成公羊免疫机能的下降。因此,在生产实践中要防范持续性环境热应激对种公羊健康及繁殖性能造成的不利影响。
参考文献:
[1]李琳,徐辰义,杨淑华,等.动物热应激反应机制研究进展[J]. 动物医学进展,2011,32(4):95-98.
[2]DAS R,SAILO L,VERMA N,et al.Impact of heat stress on health and performance of dairy animals: A review [J]. Vet World,2016,9(3):260-268.
[3]CWYNAR P,KOLACZ R,CZERSKI A.Effect of heat stress on physiological parameters and blood composition in Polish Merino rams [J].Berl Munch Tierarztl Wochenschr,2014,127(5/6):177-182.
[4]雷虹,龙云,蔡文杰,等.热应激对湘西黄牛常规血液生化指标的影响[J].湖南畜牧兽医,2016(1):38-40.
[5]姜润深,夏文涛,陈兴勇,等.利用热应激初期血清生化指标的变化幅度评价鸡的耐热性能[J].中国农业大学学报,2011,16(2):114-117.
[6]KIM B,PARK K,RHEE K.Heat stress response of male germ cells [J].Cell Mol Life Sci,2013,70(15):2 623-2 636.
[7]田允波.热应激对种公畜繁殖的影响[J].家畜生态,1994,15(1):40-42.
[8]HAMILTON T R,MENDES C M,DE CASTRO L S,et al.Evaluation of lasting effects of heat stress on sperm profile and oxidative status of ram semen and epididymal sperm [J].Oxid Med Cell Longev,2016:1687657.
[9]施力光,周雄,曹婷,等.海南夏季羊舍小气候环境分析[J].家畜生态学报,2016,37(3):73-77.
[10]RAVAGNOLO O,MISZTAL I,HOOGENBOOM G.Genetic component of heat stress in dairy cattle, development of heat index function [J].J Dairy Sci,2000,83(9):2 120-2 125.
[11]赵宗胜,米拉古丽,江华,等.冷、热应激对奶牛血液生理生化指标影响[J].中国奶牛,2011,22:18-22.
[12]黄光远,赵川,杨晓炼,等.急性持续热应激对成年兔血液部分生理生化指标的影响[J].安徽农业科学,2015,43(32):69-72.
[13]陶林,陈洁波,杜炳旺.高温热应激对麒麟鸡生理生化指标的影响[J].广东农业科学,2013,19:127-130.
[14]RANA M S,HASHEM M A,SAKIB M N,et al.Effect of heat stress on blood parameters in indigenous sheep[J].J Bangladesh Agril Univ,2014,12(1):91-94.
[15]陈小琼,肖国强.桑拿浴对人体力竭运动后外周血白细胞计数与分类的影响[J].体育研究与教育,2013,8(4):123-125.
[16]唐丽.热应激对肉种母鸡繁殖性能、相关生理生化和分子指标的影响[D].北京:中国农业科学学院,2013.
[17]白丹丹,敖日格乐,王纯洁,等.慢性冷热应激对三河牛血液生化指标及相关基因表达的影响[J].中国农业大学学报,2017,22(8):50-56.
[18]SCHARF B,CARROLL J A,RILEY D G,et al.Evaluation of physiological and blood serum differences in heat-tolerant (Romosinuano) and heat-susceptible (Angus) Bostaurus cattle during controlled heat challenge [J].J Anim Sci,2010,88(7):2 321-2 336.
[19]HOCKING P M,MAXWELL M H,MITCHELL M A.Haematology and blood composition at two ambient temperatures in genetically fat and lean adult broiler breeder females fed ad libitum or restricted throughout life [J].Br PoultSci,1994,35(5):799-807.
[20]SAKER K E,FIKE J H,VEIT H,et al.Brown seaweed-(TascoTM) treated conserved forage enhances antioxidant status and immune function in heat-stressed Wether lambs [J].J Anim Physiol Anim Nutr,2004,88(3-4):122-130.
[21]蔡明成.热应激对肉牛生理生化指标及外周血microRNA表达水平的影响[D].重庆:西南大学,2014.
[22]MAURYA V P,SEJIAN V,KUMAR D,et al.Impact of heat stress, nutritional restriction and combined stresses (heat and nutritional) on growth and reproductive performance of Malpura rams under semi-arid tropical environment[J].J AnimPhysiolAnimNutr (Berl),2016,100(5):938-946.
[23]杨明,秦应和.热应激小鼠体内睾酮浓度的变化及其与支持细胞HSPs表达的关系[J].中国畜牧杂志,2013,49(21):22-25.
[24]PEI Y,WU Y,CAO J,et al.Effects of chronic heat stress on thereproductive capacity of male Rex rabbits [J].Livest Sci,2012,146(1):13-21.
猜你喜欢
中国动物保健(2022年10期)2022-11-04 02:39:17
经济与管理(2020年4期)2020-12-28 00:44:12
世界科学(2020年1期)2020-02-11 07:39:34
中国生物医学工程学报(2019年5期)2019-07-16 07:56:56
新闻传播(2018年13期)2018-08-29 01:06:32
Coco薇(2017年5期)2017-06-05 13:03:24
初中生之友·中旬刊(2015年9期)2015-09-15 16:59:46
现代电生理学杂志(2015年4期)2015-07-18 11:00:44
爆笑show(2015年1期)2015-03-26 17:52:25
微生物与感染(2015年1期)2015-02-28 17:42:35
