
利用DH和IF2群体检测甘蓝型油菜株高相关性状QTL
2018-04-10 06:00:34贺亚军吴道明西南大学农学与生物科技学院重庆40076浙江省农业科学院作物与核技术利用研究所浙江杭州300
作物学报 2018年4期
贺亚军 吴道明 傅 鹰, 钱 伟,*西南大学农学与生物科技学院, 重庆 40076; 浙江省农业科学院作物与核技术利用研究所, 浙江杭州300
株高是油菜重要的农艺性状之一。油菜的株高不仅对产量有影响, 而且与植株的抗倒伏性密切相关[1]。首先, 倒伏会引起油菜干重、产量、出油率和收获指数大大降低, 油菜倒伏后一般减产 10%~30%, 严重时达到 60%, 甚至绝收, 倒伏油菜含油量也比正常油菜低10%~30%[2]。其次, 倒伏不利于油菜的机械化收获, 不仅降低了油菜的机收速度,而且会导致收获性减产[3]。调控株高以提高抗倒伏性一直是油菜育种的重要目标, 研究油菜株高及其相关性状的遗传机制对油菜抗倒伏和高产育种具有重要意义。
油菜的株高由第一分枝高、分枝区段长和主花序长度构成[4], 不同节间在发育的不同时期伸长,共同影响油菜的株高。油菜株高相关性状为多基因控制的数量性状, 表现为连续变异, 受环境的影响较大[5]。近年来, 国内外研究人员利用不同遗传背景的研究材料在不同环境条件下开展了油菜农艺性状的遗传机制研究, 越来越多的与甘蓝型油菜株高及其相关性状的QTL被相继发现, 这些QTL几乎覆盖了油菜的 19条连锁群[4-18]。其中控制株高的单个QTL可以解释2.4%~28.6%的表型变异, 其主效QTL位于A02、C02和C03连锁群, 分别可以解释20.8%、26.5%和28.6%的表型变异; 控制第一分枝高的单个QTL可以解释 3.92%~33.4%的表型变异, 其主效QTL位于 A02、C06连锁群, 分别可以解释 33.4%和 29.0%的表型变异; 控制主花序长度的单个 QTL可以解释 3.8%~51.8%的表型变异, 其主效 QTL位于C06连锁群, 可以解释51.8%的表型变异; 控制分枝区段长的单个 QTL可以解释 5.5%~13.6%的表型变异。虽然油菜株高相关性状的研究已有一定进展,但是这些 QTL定位中, 大部分使用单一的群体, 不能较全面分析性状的遗传效应。而不同的研究者所用的作图群体、标记类型、试验环境等不同, 造成连锁分析结果差异较大, 很难对不同研究者提出的同一连锁群上的 QTL进行比较或互相验证。因此,有关油菜株高及其相关性状的QTL研究仍需要进一步深入或完善。
本研究以双单倍体(doubled haploid, DH)群体和永久F2(immortalized F2, IF2)群体, 在2个年份环境条件下进行株高及其相关性状的QTL分析, 以阐明控制油菜株高相关性状的遗传基础, 并为分子标记辅助选择育种提供有用的遗传信息。利用2个遗传群体, 有助于分析QTL位点在不同环境和群体中的表达, 并且可以相互验证QTL定位结果的可靠性。而且, 由于IF2群体是由DH群体两两杂交而获得的,在QTL分析中兼备永久群体和分离群体的优点。这样, 除了可以用 DH群体检测性状的加性效应和上位效性外, 还可以用 IF2群体检测性状的显性效应,鉴定出在杂交后代中能传递和表达的显性效应QTL,对杂交育种有重要意义。
1 材料与方法
1.1 试验材料
以生态类型和地理来源上都存在明显差异的一个欧洲冬油菜品种“Express”和一个中国半冬性甘蓝型油菜自交系“SWU07”为亲本杂交, 获得杂种 F1再通过花药培养和染色体加倍得到一个包含261个株系的DH群体。根据Hua等[19]的组配方案构建IF2群体。在2010年和2011年, 利用其中234个DH株系, 随机分成2组, 每组117个家系, 从中随机各选择1个家系为杂交亲本配成 1个杂交组合, 再从两组剩余的家系中各选出 1个杂交组配, 直到所有株系用完,通过一轮杂交, 共组配117个杂交组合, 再通过一轮随机组配, 共获得234个杂交组合。2个试验年度间的杂交组合完全相同。
1.2 田间试验
DH群体的 2个亲本 Express和 SWU07, 以及DH群体和IF2群体于2010年和2011年种植于重庆市北碚区西南大学油菜工程研究中心试验基地。采用完全随机区组设计, 每个株系 3次重复, 每个重复30个单株, 3个行区, 每行10株, 行距为0.40 m,株距为0.24 m, 小区面积为3 m2。每年9月中旬人工播种, 10月下旬移栽到大田, 按照常规方式进行田间管理, 确保所有样本的外部生长环境一致, 待成熟后统计数据。
1.3 性状调查
油菜成熟后, 从每个株系随机选取10株测量株高(PH)、第1有效分枝高(BH)、主花序长(MFL)、分枝区段长(BS), 并数出有效一次分枝数, 计算平均节间长(AIL)。株高为从地面到植株最高部位的距离(cm); 第1分枝高为地面到主茎最底部第1有效分枝的距离(cm); 主花序长为从主茎最上端有效分枝到主茎顶部的距离(cm); 分枝区段长为主茎最上端有效分枝到主茎最底部第 1有效分枝的距离(cm)[4];一次分枝数为植株有效的一次分枝数; 平均节间长为分枝区段长/(一次分枝数-1)。取每个性状重复观测值的平均值作为各个株系的表型值。
1.4 遗传连锁图谱与QTL分析
遗传图谱包含293个SSR多态性标记位点, 总长1188 cM, 相邻标记间平均距离为4.05 cM[20]。采用WinQTL Cartographer 2.5软件中的复合区间作图法进行株高及其相关性状的QTL定位[21-22]。QTL扫描的参数设置是以5为背景标记数目, 10 cM作为窗口大小, 1 cM为扫描步长。在P=0.05的概率背景下进行1000次的排布测验[23], 用以确定LOD阈值并作为 QTL存在的标准。LOD≥2.5时就认为这个置信区间内存在一个QTL。对于同一性状的QTL在不同试验环境或者不同群体中置信区间发生重叠, 则认为它们为同一个QTL。
2 结果与分析
2.1 性状表型分析
2个亲本Express和SWU07在株高、第1分枝高、主花序长、分枝区段长和平均节间长上均有明显差别(图 1和表 1), 后代 DH群体和 IF2群体在 5个株高及其相关性状上均表现为连续变异, 并且存在明显的双向超亲分离现象(图2和表1), 表明这些性状均为多基因控制的数量性状。

表1 亲本及2个群体5个株高相关性状在2年的表型分析Table 1 Analysis of five plant height related traits in the two parents and two populations in 2010 and 2011 (cm)

图1 亲本 SWU07与ExpressFig. 1 Parental lines, SWU07 and Express
同一性状在年份间的相关性分析表明(表2), 在DH群体中, 同一性状在2年间的表现均呈极显著正相关, 且相关系数较大, 在IF2群体中, 5个株高相关性状中的3个, 株高、第一分枝高和主花序长各性状在2年间的表现也呈极显著正相关, 且相关系数较大。这说明这些株高相关性状遗传稳定。
性状间的相关性分析表明(表3), 在2年环境下,株高分别与主花序长、分枝区段长、平均节间长在2个群体均呈显著正相关, 株高与第1分枝高在DH群体相差不显著, 但在IF2群体中呈显著正相关, 而第 1分枝高分别与主花序长、分枝区段长、平均节间长在2年环境下2个群体中均呈显著负相关。主花序长分别与分枝区段长、平均节间长在2011年环境下2个群体均呈显著负相关, 在2010年环境下在IF2群体呈显著负相关, 在DH群体中相差不显著。分枝区段长与平均节间长在2年环境下2个群体中均呈显著正相关。综合2个群体分析结果, 除在DH群体中株高与第 1分枝高相关性不显著外, 株高分别与其构成部分第 1分枝高、主花序长、分枝区段长的相关性在2年2个群体中均显著, 而第1分枝高分别与株高的另外2个构成部分主花序长和分枝区段长在2年环境下2个群体中均呈显著负相关, 表明第 1分枝高、主花序长、分枝区段长分别对株高都发挥着极大作用。另外, 除个别性状间的相关性在不同群体间表现出差异外, 同一对性状在不同群体中的遗传相关系数差别不大。大多数性状在不同群体中的相关系数在方向上都是一致的, 只是程度上略微不同。这一结果表明, 株高相关性状间的遗传相关对群体类型的依赖程度较小, 不同的群体类型可以反应出相似的遗传相关。

图2 DH群体与IF2群体株高相关性状在2010年和2011年的频率分布Fig. 2 Frequency distribution of plant height related traits from DH and IF2 populations in 2010 and 2011
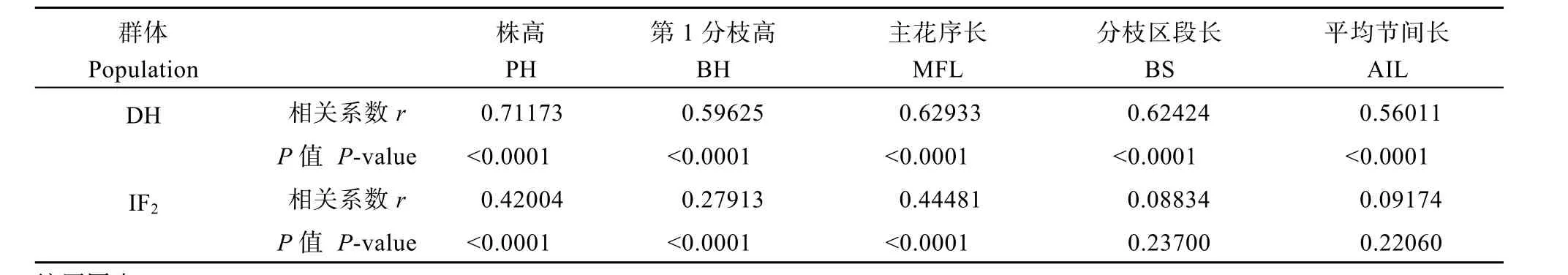
表2 各性状在2个年份间的相关性分析Table 2 Correlation analysis for plant height related traits in 2010 and 2011

表3 DH群体和IF2群体各性状间的相关系数Table 3 Correlation coefficients among plant height-related traits in DH and IF2 populations
2.2 株高及其相关性状在 2年 2个群体的 QTL分析
不同年份环境下DH群体和IF2群体中, 对5个株高相关性状共检测到46个QTL (图3和表4), 其中在DH群体中2010年检测到15个QTL, 2011年检测到 17个 QTL; 在 IF2群体中 2010年检测到 6个QTL, 2011年检测到8个QTL。这些QTL分布于甘蓝型油菜的13条连锁群上。将每个性状在不同年份或者不同群体中重复检测到的QTL整合, 发现有5个 QTL在不同年份或不同群体中被重复检测到,因此, 最终得到41个位点与这5个性状相关。
2.2.1株高在2年两群体中共检测到11个与植株高度相关的QTL, 分布于甘蓝型油菜的5条连锁群(A02、A09、C01、C02 和 C06), 分别揭示了 3.85%~13.34%的株高表型变异。其中, qPH-C01-1-2010DH和 qPH-C01-1-2011DH可以在不同年份环境中被重叠检测到, qPH-C01-2-2010DH和qPH-C01-2-2011DH可以在不同年份环境中被重叠检测到, 最终将其整合为2个可重复性的位点。这样, 通过整合这些可重复性的位点, 最终得到9个与株高相关的位点。
2.2.2主花序长在2年2个群体中共检测到18个与主花序长度相关的 QTL, 分布于甘蓝型油菜的8条连锁群(A01、A02、A05、A08、A09、C01、C03和 C05), 分别揭示了 3.82%~9.52%的主花序长度表型变异。其中, qMFL-A02-1-2011DH和qMFL-A02-2011IF2可以在不同群体中被重叠检测到, qMFLC01-2010IF2和qMFL-C01-2011IF2可以在不同年份环境中被重叠检测到, qMFL-C03-1-2011DH和qMFL-C03-2011IF2可以在不同群体中被重叠检测到, 终将其整合为3个可重复性的位点。这样, 通过整合这些可重复性的位点, 最终得到15个与主花序长度相关的位点。
2.2.3第1分枝高度在2年2个群体中共检测到11个与第1分枝高度相关的QTL, 分布于甘蓝型油菜的 5条连锁群(A01、A03、A09、C01和 C03), 这些 QTL分别揭示了 4.01%~16.54%的分枝高度表型变异。在这11个QTL中, 没有QTL可以在不同年份环境或者不同群体中被重叠检测到。
2.2.4分枝区段长在2年2个群体中共检测到4个与分枝区段长相关的 QTL, 分布于甘蓝型油菜的4条连锁群(A07、A09、C03和C04), 分别揭示了4.79%~8.10%的分枝区段长表型变异。在这4个QTL中, 没有QTL可以在不同年份环境或者不同群体中被重叠检测到。


表4 在DH群体和IF2群体中检测到的株高相关性状QTLTable 4 Putative QTLs for plant height related traits detected in DH and IF2 populations.
2.2.5平均节间长在2年2个群体中共检测到2个与平均节间长相关的 QTL, 分布于甘蓝型油菜的 2条连锁群(A07和 C05), 分别揭示了 4.29%~6.04%的平均节间长表型变异。在这2个QTL中, 没有QTL可以在不同年份环境或者不同群体中被重叠检测到。
3 讨论
株高是油菜的重要农艺性状之一, 长期以来一直受到育种家的重视。控制油菜株高无疑是防止倒伏从而增加产量和提高油菜生产效益的有效方法之一[24], 这在其他作物上已不乏先例, 如在水稻和小麦上矮杆和半矮杆的推广利用, 极大地提高了产量,被誉为第一次绿色革命的标志[25]。第1分枝高、分枝区段长和主花序长度构成了油菜的株高[4], 不同部位在发育的不同时期伸长, 共同影响油菜的株高。
本研究利用DH和IF22个群体有助于分析QTL位点在不同环境和群体中的表达, 并且可以相互验证 QTL定位结果的可靠性。在本研究中, 检测到 2个控制主花序长的QTL在2个群体中都表达, 它们分别位于 A02、C03连锁群(表 4)。由于 IF2群体是由DH群体两两杂交而获得的, 因此, 这些QTL能够在杂交后代中传递和表达, 这些QTL对分子标记辅助选择育种有更重要的利用价值, 对油菜杂交育种有更重要的作用。
本研究表明(表3), 株高相关性状间存在显著的相关性。前人研究认为, 性状间的相关在QTL分析结果上也会表现, 可能存在控制不同性状QTL之间紧密连锁或同一QTL控制不同性状的现象, 即一因多效[26]。本研究中, 在A01染色体上检测到3个与第 1分枝高和主花序长相关的 QTL, 形成交叠的QTL簇。同样在A02染色体存在控制株高和主花序长的交叠QTL, 在A07染色体存在控制分枝区段长和平均节间长的交叠QTL, 在A09染色体存在控制分枝区段长和主花序长的交叠QTL以及控制第1分枝高和株高的交叠QTL, 在C01染色体存在控制株高、第1分枝高和主花序长的交叠QTL, 在C03染色体存在控制第1分枝高和主花序长的交叠QTL。这进一步证明了株高相关性状间存在相关性。这些QTL附近的标记将有利于油菜株高的辅助选择育种。尤其值得关注的是, 在C01染色体的邻近区域,检测到不同性状、不同群体、不同年度的13个QTL,且效应相对较大。对于这些C01染色体上重点关注的QTL, 我们已经开展了 QTL的精细定位工作, 同时也开发了紧密连锁的分子标记用于育种的辅助选择。
关于油菜株高相关性状的QTL分析, 前人的定位结果[4-18]不尽一致。本研究获得了油菜株高及其相关性状的QTL 41个, 分布于油菜的13个连锁群。5个性状中, 株高、主花序长都有可重复检测到的QTL, 而第 1分枝高、分枝区段长和平均节间长 3个性状都没有, 这可能是由于这些性状受环境影响较大。与已报道的油菜株高相关QTL相比较, 前人对株高、第 1分枝高、主花序长研究较多, 而对分枝区段长, 目前, 只有Cai等[4]报道了关于油菜分枝区段长的QTL定位结果, 发现了13个控制分枝区段长的QTL分布于12个连锁群A02、A06、A07、A08、A09、A10、C03、C04、C06、C07、C08 和 C09。对平均节间长, 几乎没有报道。在油菜株高的构成中, 除了第 1分枝高和主花序长外, 分枝区段也是株高最重要的组成部分, 因此, 研究油菜分枝区段长和平均节间长, 对进一步理解油菜株高相关性状的遗传机制具有很重要作用。在本研究中, 除了对前人研究较多的株高、第1分枝高、主花序长进行QTL检测外, 还对分枝区段长和平均节间长进行了QTL检测, 研究结果可为油菜株高相关性状的遗传机制提供有用的信息。
4 结论
株高除在DH群体中与第1分枝高不相关外, 分别与其构成部分第一分枝高、主花序长、分枝区段长在2年2个群体中均呈显著正相关, 而第1分枝高、主花序长、分枝区段长两两间呈显著负相关。利用DH和IF22个群体共检测到41个油菜株高相关性状QTL, 其中在2个年份环境下或2个群体中重复检测到5个QTL, 位于A02、C01 和C03染色体上。
[1] 周清元, 李军庆, 崔翠, 卜海东, 阴涛, 颜银华, 李加纳, 张正圣. 油菜半矮杆新品系10D130株型性状的遗传分析. 作物学报, 2013, 39: 207-215 Zhou Q Y, Li J Q, Cui C, Bo H D, Yin T, Yan Y H, Li J N, Zhang Z S. Genetic analysis of plant type in semi-dwarf new line(10D130) of rapeseed.Acta Agron Sin, 2013, 39: 207-215 (in Chinese with English abstract)
[2] 刘后利. 油菜遗传育种学. 北京: 中国农业大学出版社, 2000.pp 32-45 Liu H L. Genetics and Breeding of Rapeseed. Beijing: China Agricultural University Press, 2000. pp 32-45
[3] Islam N, Evans E. InXuence of lodging and nitrogen rate on the yield and yield attributes of oilseed rape (Brassica napusL.).Theor Appl Genet, 1994, 88: 530-534
[4] Cai G, Yang Q, Chen H, Yang Q, Zhang C, Fan C, Zhou Y.Genetic dissection of plant architecture and yield-related traits inBrassica napus.Sci Rep, 2016, 6: 21625
[5] 易斌, 陈伟, 马朝芝, 傅廷栋, 涂金星. 甘蓝型油菜产量及相关性状的QTL分析. 作物学报, 2006, 32: 676-682 Yi B, Chen W, Ma C Z, Fu T D, Tu J X. Mapping of quantitative trait loci for yield and yield components inBrassica napusL.Acta Agron Sin, 2006, 32: 676-682 (in Chinese with English abstract)
[6] 王嘉, 荆凌云, 荐红举, 曲存民, 谌利, 李加纳, 刘列钊. 甘蓝型油菜株高、第一分枝高和分枝数的QTL检测及候选基因筛选. 作物学报, 2015, 41: 1027-1033 Wang J, Jing L Y, Jian H J, Qu C M, Chen L, Li J N, Liu L Z.Quantitative trait loci mapping for plant height, the first branch height, and branch number and possible candidate genes screening inBrassica napusL.Acta Agron Sin, 2015, 41: 1027-1033(in Chinese with English abstract)
[7] Cai D, Xiao ·Y, Yang W, Ye W, Wang B, Younas M, Wu J, Liu K.Association mapping of six yield‑related traits in rapeseed (Brassica napusL.).Theor Appl Genet, 2014, 127: 85-96
[8] Chen W, Zhang Y, Liu X P, Chen B Y, Tu J X, Fu T D. Detection of QTL for six yield-related traits in oilseed rape (Brassica napus)using DH and immortalized F2populations.Theor Appl Genet,2007, 115: 849-858
[9] Zhang S H, Fu T D, Zhu J C, Wang J P, Wen Y C, Ma C Z, Jiang Y Z. QTL mapping and epistasis analysis for plant height and height to the first branch in rapeseed (Brassica napusL.).BIOTECHNOLOGY: Gene Clon Funct Anal, 2007, 2: 232-235
[10] Shi J Q, Li R Y, Qiu D, Jiang C C, Long Y, Morgan C, Nancroft I,Zhao J Y, Meng J L. Unraveling the complex trait of crop yield with quantitative trait loci mapping inBrassica napus.Genetics,2009, 182: 851-861
[11] Shi T X, Li R Y, Zhao Z K, Ding G D, Long Y, Meng J L, Xu F S,Shi L. QTL for yield traits and their association with functional genes in response to phosphorus deficiency inBrassica napus.PLoS One, 2013, 8: e54559
[12] Ding G D, Zhao Z K, Liao Y, Hu Y F, Shi L, Long Y, Xu F S.Quantitative trait loci for seed yield and yield-related traits, and their responses to reduced phosphorus supply inBrassica napus.Ann Bot, 2012, 109: 747-759
[13] Udall J A, Quijada P A, Lambert B, Osbom T C. Quantitative trait analysis of seed yield and other complex traits in hybrid spring rape seed (Brassica napusL.): 2. Identification of alleles from unadapted germplasm.Theor ApplGenet, 2006, 113: 597-609
[14] Butruille D V, Guries R P, Osbom T C. Linkage analysis of molecular markers and quantitative trait loci in populations of inbred backcross lines ofBrassica napusL.Genetics, 1999, 153: 949-964
[15] Chen W, Zhang Y, Liu X P, Chen B Y, Tu J X, Fu T D. Detection of QTL for six yield-related traits in oilseed rape (Brassica napus)using DH and immortalized F2populations.Theor Appl Genet,2007, 115: 849-858
[16] 张凤启, 刘越英, 程晓晖, 童超波, 董彩华, 唐敏强, 黄军艳,刘胜毅. 利用高密度 SNP标记定位甘蓝型油菜株高 QTL. 中国油料作物学报, 2014, 36: 695-700 Zhang F Q, Liu Y Y, Cheng X H, Tong C B, Dong C H, Tang M Q, Huang J Y, Liu S Y. QTL mapping of plant height using high density SNP markers inBrassica napus.Chin J Oil Crop Sci,2014, 36: 695-700 (in Chinese with English abstract)
[17] Zhao W, Wang X, Wang H, Tian J, Li B, Chen L, Chao H, Long Y,Xiang J, Gan J, Liang W, Li M. Genome-wide identification of QTL for seed yield and yield-related traits and construction of a high-density consensus map for QTL comparison inBrassica napus. Front Plant Sci, 2016, 7: 17
[18] Wang X, Wang H, Long Y, Liu L, Zhao Y, Tian J, Zhao W, Li B,Chen Li, Chao H, Li M. Dynamic and comparative QTL analysis for plant height in different developmental stages ofBrassica napusL.Theor Appl Genet, 2015, 128: 1175-1192
[19] Hua J P, Xing Y Z, Wu W R, Xu C G, Sun X L, Yu S B, Zhang Q F.Single-locus heterotic effects and dominance by dominance interaction can adequately explain the genetic basis of heterosis in an elite hybrid.Proc Natl Acad Sci USA, 2003, 100: 2574-2579
[20] Fu Y, Lu K, Qian L, Mei J, Wei D, Peng X, Xu X, Li J, Frauen M,Dreyer F, Snowdon R J, Qian W. Development of genic cleavage markers in association with seed glucosinolate content in canola.Theor Appl Genet, 2015, 128: 1029-1037
[21] Zeng Z B. Theoretical basis for separation of multiple linked gene effects in mapping quantitative trait loci.Proc Natl Acad Sci USA, 1993, 90: 10972-10976
[22] Zeng Z B. Precision mapping of quantitative trait loci.Genetics,1994, 136: 1457-1468
[23] Churchill G A, Doerge R W. Empirical threshold values for quantitative trait mapping.Genetics, 1994, 138: 963-971
[24] 李云, 付绍红, 杨进, 王继胜, 邹琼, 陈晓华, 陶兰蓉, 康泽明,唐蓉, 张汝全. 甘蓝型油菜矮秆突变体bndf-1的遗传鉴定及利用潜力分析. 中国农学通报, 2013, 29(13): 173-177 Li Y, Fu S H, Yang J, Wang J S, Zou Q, Chen X H, Tao L R,Kang Z M, Tang R, Zhang R Q. The identification and application of dwarf mutationbndf-1inBrassica napus.Chin Agric Sci Bull, 2013, 29(13): 173-177 (in Chinese with English abstract)
[25] Khush G S. Green revolution: the way forward.Nat Rev Genet,2001, 2: 815-822
[26] Tuberosa R, Salvi S, Sanguineti M C, Landi P, Maccaferri M,Conti S. Mapping QTLs regulating morpho-physiological traits and yield: case studies, shortcomings and perspectives in droughtstressed maize.Ann Bot, 2002, 89: 941
猜你喜欢
——植物学教材质疑(六)
生命科学研究(2022年1期)2022-03-14 07:50:40
语文周报·教研版(2021年28期)2021-08-19 02:14:42
西藏农业科技(2019年3期)2019-11-04 00:35:14
西藏农业科技(2019年1期)2019-07-25 00:37:02
西藏农业科技(2018年4期)2018-04-25 06:39:28
南风(2017年34期)2018-01-16 07:29:09
数学理论与应用(2017年2期)2017-06-27 07:38:56
数学理论与应用(2017年2期)2017-06-27 07:38:54
科学种养(2017年6期)2017-06-13 08:42:04
科学种养(2017年5期)2017-05-17 10:18:50
