
转基因抗草甘膦棉栽培地生存竞争能力
2018-03-26 06:56:58姜伟丽马小艳任相亮胡红岩马亚杰
生物安全学报 2018年1期
姜伟丽, 马小艳, 任相亮, 胡红岩, 马亚杰, 王 丹, 马 艳
中国农业科学院棉花研究所/棉花生物学国家重点实验室,河南 安阳 455000
据国际农业生物技术应用服务组织(2017)发布的《2016年全球生物技术/转基因作物商业化发展态势》显示,全球转基因作物的种植面积从1996年的170万hm2增加到1.851亿hm2,增加了110倍。其中,耐除草剂作物的种植面积为8650万hm2,占全球转基因作物种植面积的47%。草甘膦和草铵膦两大灭生广谱性除草剂被首选,主要应用于转基因大豆、油菜、玉米、苜蓿和棉花等作物。目前,抗草甘膦性状已在大豆、棉花和油菜作物中成功应用并在市场上占据了主导地位(郭书巧等,2012; 邱龙等,2012)。
近年来,转基因作物新品种的大面积推广种植在带来直接经济与生态效益的同时,其生物安全问题在全世界范围内引起了广泛关注。因此,研发与安全评价同步是转基因作物育种的重要原则。转基因作物环境安全性评价主要包括基因漂移、与近缘野生种的可交配性、对非靶标生物的影响及生存竞争性等方面。其中,转基因作物的生存竞争能力是转基因作物环境安全性评价的重要内容之一(刘娜等,2006; James,1996)。针对我国研发的耐除草剂转基因作物在生存竞争能力方面的评价有:在荒地的生存竞争能力且是否能演变成超级杂草的可能性(陈小文等,2012; 崔荣荣等,2012; 马小艳等,2013; 宋小玲等,2009; 孙红炜等,2014),以及在栽培地的生存竞争能力(马小艳等,2014; 张兴华等,2010)。但目前,有关转基因抗草甘膦棉花的环境适应和生存竞争能力鲜有报道。因此,本试验选用我国研发的具有自主知识产权的转EPSPS基因抗除草剂棉花GGK-2为研究对象,参照农业部953号公告-12-2007《转基因植物及其产品环境安全检测 抗虫棉花 第2部分:生存竞争能力》(中华人民共和国农业部,2007),对其栽培地生存竞争能力进行研究,以期为转基因抗草甘膦棉花的生产应用提供理论依据。
1 材料与方法
1.1 材料
试验品种为转EPSPS基因抗草甘膦棉(GGK-2),对照品种为受体材料(K312)及当地主栽品种中棉所49(简称中49),均由中国农业科学院棉花研究所育种研究室提供。
1.2 方法
1.2.1 试验设计 试验在河南安阳中国农业科学院棉花研究所试验农场中进行。于2016年4月23日播种,每个小区面积为25.6 m2(行宽0.8 m,行长8 m,4行区)。每种材料设4次重复,随机区组排列,种植密度为5.5万株·hm-2,农事操作按常规管理进行。
1.2.2 调查内容及方法 棉花生长发育指标的调查:于棉花苗期(6月3日)、现蕾期(6月29日)、花铃期(7月27日)和吐絮期(9月12日)各调查一次,每个小区随机选取10株棉花,测量其株高、茎直径、果枝数、蕾铃数等生长指标。
棉花产量及品质调查:10月初,每个小区均匀采收25铃并进行室内考种,考种项目包括铃重、衣分、籽指、棉籽数等产量参数;然后随机抽取每个小区采收的棉纤维约100 g,由中华人民共和国农业部棉花品质监督检验测试中心对棉纤维样品的5项品质进行分析,包括纤维长度、整齐度指数、断裂比强度、断裂伸长率和马克隆值。
1.3 数据分析
运用SPSS 13.0 软件对所有数据进行Duncan′s差异显著性分析,检验转基因棉花与受体材料和当地主栽品种间生长参数及棉花产量和品质指标的差异。
2 结果与分析
2.1 营养生长竞争力
由表1可以看出,转基因抗草甘膦棉花在苗期、现蕾期、花铃期和吐絮期的株高均较低,与受体材料相比分别低0.12、2.91、7.78和0.95 cm,与中49相比分别低1.42、2.99、1.45和4.40 cm,但差异均未达到显著水平(P>0.05)。转基因抗草甘膦棉花在苗期、花铃期和吐絮期的茎直径均低于受体材料,分别低0.13、0.60和1.57 mm,在现蕾期的茎直径比受体材料高0.78 mm,但这些差异均未达到显著水平(P>0.05);转基因抗草甘膦棉花在苗期和现蕾期的茎直径比中49分别低0.11和0.41 mm,到花铃期和吐絮期却比中49分别高0.20和0.98 mm,但这些差异均不显著(P>0.05)。因此,转基因抗草甘膦棉花与2个对照品种相比在营养生长方面无竞争优势。
2.2 生殖生长竞争力
由表1可以看出,在苗期、现蕾期、花铃期和吐絮期,3个品种间的果枝数均无显著差异(P>0.05)。花铃期,转基因抗草甘膦棉花的每株花蕾数比受体材料少3.58个,差异达到显著水平(P<0.05),但与对照品种中49相当;现蕾期和吐絮期,3个品种间的每株花蕾数无显著差异(P>0.05)。花铃期,转基因抗草甘膦棉花与受体材料的每株铃数无显著差异(P>0.05),但二者均显著低于中49(P<0.05);吐絮期,3个品种的每株铃数差异不显著(P>0.05)。总体来说,与受体材料和中49相比,转基因抗草甘膦棉花在生殖生长方面无竞争优势。
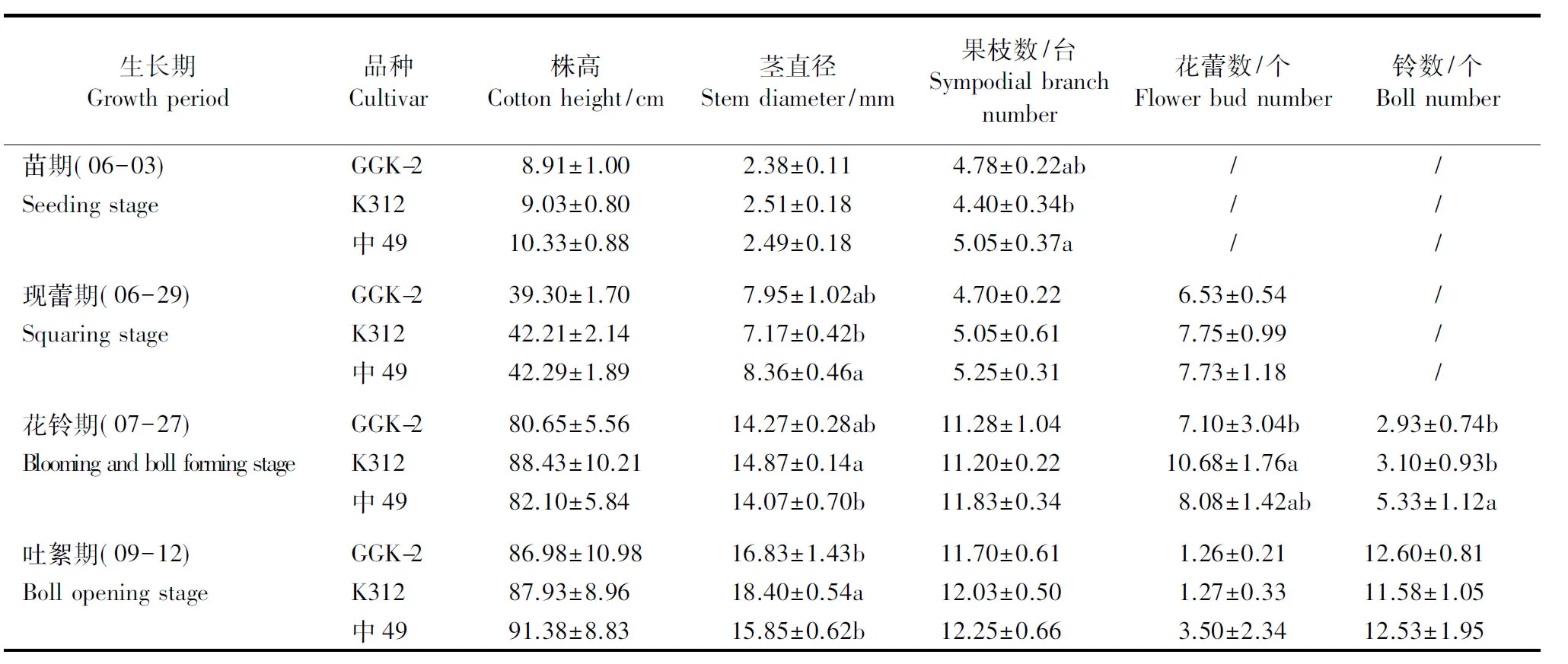
表1 转基因抗草甘膦棉花品系的生物参数Table 1 Biological parameters of the transgenic glyphosate-resistant cotton strain
数据为平均值±标准差;同一列中小写字母不同者表示同一时期不同品种间在0.05水平上差异显著。
Values are means±SD; different small letters in the same column represent significant differences among different cultivars in the same growth period at 0.05 level.
2.3 产量及品质竞争力
测产结果(表2)表明,转基因抗草甘膦棉花的单铃重比中49低0.80 g,差异达到显著水平(P<0.05),但与受体材料无明显差异(P>0.05);3个品种的衣分、籽指和衣指无显著差异(P>0.05)。转基因抗草甘膦棉花和受体材料的籽棉产量均低于中49,但转基因抗草甘膦棉花产量比受体材料高91.4 kg·hm-2,差异均不显著(P>0.05)。通过对棉纤维5项品质指标的检测(表3)可知,与受体材料相比,转基因抗草甘膦棉花的整齐度指数、断裂伸长率、马克隆值均有一定程度的提高,但差异不显著(P>0.05)。总体来说,与受体材料相比,转基因抗草甘膦棉花能在一定程度上提高棉花产量,改善棉花纤维品质,具有一定的竞争优势,但这种优势未达到显著水平。
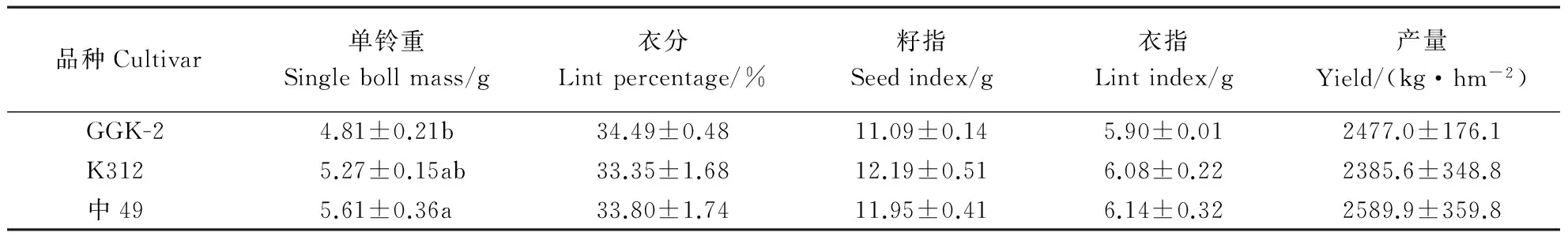
表2 转基因抗草甘膦棉花品系的产量竞争力Table 2 Yield competition of transgenic glyphosate-resistant cotton strain
数据为平均值±标准差;同一列中小写字母不同者表示在0.05水平上差异显著。
Values are means±SD; different small letters in the same column represent significant differences at 0.05 level.

表3 转基因抗草甘膦棉花品系的品质竞争力Table 3 Fiber quality competition of transgenic glyphosate-resistant cotton strain
数据为平均值±标准差;同一列中小写字母不同者表示在0.05水平上差异显著。
Values are means±SD; different small letters in the same column represent significant differences at 0.05 level.
3 讨论
Bartschetal.(1996,2001)研究表明,当外源基因转入受体植物后,植物机体为了维持转入基因的运转,需要消耗一定的能量,可能造成受体植物在能量分配上的改变,从而导致植物在生长发育和繁殖能力方面的损耗,不表现竞争优势;同时,外源基因的随机插入可能会引起植物生理生化及代谢途径的改变,从而导致转基因植物逆境应答能力的变化(李维江等,2001; 聂呈荣等,2006)。Benbrook (1999)和Elmoreetal.(2001)研究表明,抗草甘膦转基因大豆的田间产量比非转基因品系低5%~10%,认为外源基因的随机插入导致的非预期效应可能是转基因大豆减产的主要原因。康岭生等(2010)研究表明,在正常播种条件下,高油酸转基因大豆HOA80的生存竞争能力和繁育能力与受体大豆相一致。
本研究结果表明,转基因抗草甘膦棉花在苗期、现蕾期、花铃期和吐絮期的株高均低于受体材料,但差异未达到显著水平;在花铃期每株花蕾数显著低于受体材料,但在吐絮期单株铃数多于受体材料;同时,其产量比受体材料高91.4 kg·hm-2,棉纤维品质也有一定程度的改善。因此,外源基因的插入在某种程度上改变了棉花本身的生长性状,但这种改变并未影响棉花后期的产量和纤维品质,反而起到了一定的改善作用。此外,转基因抗草甘膦棉花的各项生长指标、产量及纤维品质均与对照品种中棉所49相当。由此可知,转基因抗草甘膦棉花在栽培地环境下可正常生长,不会对农业生产造成明显的负面影响,具有一定的栽培价值;但有关其在栽培地的生存竞争优势还有待进一步研究和改进。
陈小文, 李吉崇, 郭玉海, 董学会, 2012. 抗虫转基因玉米荒地生存竞争力评价. 杂草科学, 30(1): 31-34.
崔荣荣, 韦颖, 孟攀潘, 马云龙, 金媛, 李秋然, 强胜, 戴伟民, 宋小玲, 2012. 抗草铵膦转基因水稻明恢86B杂草化潜力评价. 中国水稻科学, 26(4): 467-475.
国际农业生物技术应用服务组织, 2017. 2016年全球生物技术/转基因作物商业化发展态势. 中国生物工程杂志, 37(4): 1-8.
郭书巧, 倪万潮, 王庆南, 沈新莲, 束红梅, 张香桂, 徐鹏, 巩元勇, 2012. 转基因抗除草剂棉花的创制及产业化前景. 江苏农业科学, 28(5): 943-947.
康岭生, 杨向东, 王玉民, 谭喜昌, 宋新元, 于壮, 张欣芳, 张明, 2010. 高油酸转基因大豆HOA80生存竞争能力检测. 吉林农业科学, 35(6): 1-3.
李维江, 张冬梅, 唐薇, 董合忠, 2001. 转Bt基因抗虫棉和有色棉苗期耐盐性差异研究. 棉花学报, 13(4): 234-237.
刘娜, 李葱葱, 徐文静, 董英山, 2006. 转基因作物环境安全性研究进展. 分子植物育种, 4(1): 9-14.
马小艳, 马艳, 彭军, 姜伟丽, 马亚杰, 2013. 转基因抗草甘膦抗虫棉的荒地生存竞争能力. 生物安全学报, 22(2): 103-108.
马小艳, 彭军, 姜伟丽, 马亚杰, 马艳, 2014. 转双价基因(Chi+Glu)抗病棉花栽培地生存竞争能力研究. 中国棉花, 41(9): 22-24.
聂呈荣, 骆世明, 王建武, 2006. Bt玉米光合作用和生长性状的变化. 生态学报, 26(6): 1957-1962.
邱龙, 马崇烈, 刘博林, 章旺根, 2012. 耐除草剂转基因作物研究现状及发展前景. 中国农业科学, 45(12): 2357-2363.
宋小玲, 强胜, 彭于发, 2009. 抗草甘膦转基因大豆(Glycinemac(L.) Merri)杂草性评价的试验实例. 中国农业科学, 42(1): 145-153.
孙红炜, 杨淑珂, 李凡, 徐晓辉, 高瑞, 路兴波, 2014. 抗虫耐除草剂棉花生存竞争能力研究. 山东农业科学, 46(2): 100-103.
张兴华, 乔艳艳, 李捷, 2010. 转双价(Chi+Glu)基因抗病棉栽培地生存竞争能力研究. 江西农业学报, 22(6): 1-3.
中华人民共和国农业部, 2007. 转基因检测标准: 第953号公告. 北京: 中华人民共和国农业部.
BARTSCH D, BRAND U, MORAK C, POHL-ORF M, SCHUPHAN I, ELLSTRAND N C, 2001. Biosafety of hybrids between transgenic virus-resistant sugar beet and swiss chard.EcologicalApplications, 11(1): 142-147.
BARTSCH D, SCHMIDT B, POHL-ORF M, HAAG C, SCHUPHAN I, 1996. Competitiveness of transgenic sugar beet resistant to beet necrotic yellow vein virus and potential impact on wild beet populations.MolecularEcology, 5(2): 199-205.
BENBROOK C M, 1999.EvidenceoftheMagnitudeandConsequencesoftheRoundupReadySoybeanYieldDragfromUniversity-BasedVarietalTrialsin1998. Sandpoint, Idaho: Benbrook Consulting Services.
ELMORE R W, ROETH F W, NELSON L A, SHAPIRO C A, KLEIN R N, KNEZEVIC S Z, 2001. Glyphosate-resistant soybean cultivar yields compared with sister lines.AgronomyJournal, 93(2): 408-412.
JAMES K, 1996. Could transgenic super crops one day breed super weeds.Science, 274: 180-181.
猜你喜欢
疯狂英语·读写版(2024年1期)2024-03-13 13:22:55
农药科学与管理(2019年6期)2019-11-23 08:17:10
环境科技(2015年4期)2015-11-08 11:10:48
农民致富之友·下半月(2015年6期)2015-07-09 10:46:48
农民致富之友(2015年12期)2015-06-11 19:33:04
营销界(2015年23期)2015-02-28 22:06:12
中国农资(2015年22期)2015-02-01 12:30:07
乡村科技(2014年15期)2014-10-29 07:32:46
现代农业科技(2014年13期)2014-09-18 09:33:38
新疆农垦科技(2014年5期)2014-02-28 19:19:56
