
miR-26调控BID蛋白对结肠癌细胞增殖和侵袭的影响
2018-03-20 07:14汪旭伟王建伟陈玉星
胃肠病学和肝病学杂志 2018年3期
汪旭伟,马 沛,王建伟,陈玉星
南阳市第一人民医院普外科,河南 南阳 473010
结肠癌是消化系统最常见的恶性肿瘤,每年全球的发病率仅次于胃癌[1]。然而,由于其异质性和多变性,结肠癌的具体分子发病机制尚未明确[2]。家族遗传性和基因突变也是结肠癌发病机制中的重要部分[3-4]。因此,目前需要更好地了解结肠癌发病的分子基础和发现对抗结肠癌的有效靶向药物。
微小RNA(miRNA)是一种由20~25个核苷酸组成的非编码RNA,可以通过与靶向mRNA的3′UTR段结合配对达到促进mRNA降解和翻译的抑制过程[5-7]。目前已经确定的相关miRNA在肿瘤中发挥致癌和抑癌作用的就有数百种,而且在不同类型的肿瘤miRNA扮演的角色也不尽相同[8]。最近各种报道[9-10]提出,多种miRNA在结肠癌中表达异常,且存在表达失控的现象(例如let-7、miR-21、miR-26、miR-221和miR-222等),对结肠癌细胞的增殖、迁移、侵袭和凋亡行为都有一定的调节能力。
肿瘤细胞的增殖、侵袭和凋亡在肿瘤发生、发展过程中具有重要作用[11]。BID作为Bax家族成员之一,对细胞损伤和线粒体修复有一定的调控作用,可以决定一群细胞的分化方向和存活比例[12]。Bax家庭成员拥有同源性结构域,称为BH1、BH2、BH3和BH4,其中BID占据最为关键的BH2[13]。
在我们的研究中,通过调控miR-26的表达,检测BID的表达水平,发现miR-26和BID有相互调节的关系,miR-26表达的下调可以通过BID的表达间接影响结肠癌的行为。
1 材料与方法
1.1细胞培养及标本HT29、SW480、CW-2和SW620细胞购自武汉典型培养物保藏中心,并在含有质量浓度为100 g/L的FBS、100 mg/ml青霉素和100 mg/ml链霉素的DMEM培养基中,37 ℃,在体积分数为5%的CO2培养箱中培养。2016年1月至2016年12月在本院接受结肠癌组织切除术的80例患者得到新鲜的结肠癌组织和相邻结肠组织,手术切除后立即将组织放置液氮冷冻,储存于-80 ℃直至使用。来自病理科的60例患者的结肠癌组织均获得了医院保护人类受试者委员会的批准,并获得了所有患者的知情同意。
1.2miR-inhibitor转染实验miRNA和小干扰RNA(siRNA)由GenePharma(中国上海)合成。miRNA模拟物是代表成熟miRNA的合成双链体。使用lip2000(QIAGEN)进行siRNA和miRNA转染。20 nmol/L siRNA或miRNA用于无血清培养基中的转染。转染后48~72 h制备总RNA和蛋白质,并进一步用于定量聚合酶链反应(PCR)或蛋白质印迹分析。
1.3mRNA的定量分析根据制造商的方案,使用miRVana miRNA分离试剂盒(Ambion)从培养的细胞或结肠癌的临床样品中提取总RNA和miRNA。使用TaqManq PCR方法来检测组织和细胞中miR-26的表达。对于mRNA表达的定量分析,使用Primescript RT试剂盒(TaKaRa)检测。实验重复3次。
1.4蛋白质提取和免疫印迹使用细胞裂解缓冲液(Cell Signaling)裂解细胞,用BCA蛋白质测定试剂盒(Pierce)测定蛋白质浓度。将等量的总蛋白质在质量浓度为100 g/L的SDS聚丙烯酰胺凝胶中分离并转移到聚偏二氟乙烯膜(Bio-Rad)。用含有质量浓度为0.5 g/L的Tween20的Tris缓冲盐水冲洗3次,然后使用质量浓度为50 g/L的牛血清白蛋白封闭PVDF膜2 h,然后孵育BID一抗(浓度1∶1 000),4 ℃冰箱,孵育过夜,次日,使用含有质量浓度为0.5 g/L的Tween20的Tris缓冲盐水冲洗3次,然后孵育二抗(浓度1∶5 000),之后通过ECL显影。
1.5荧光素酶报告基因测定验证miR-26和BID的相关关系,使用双荧光素酶测定,将24孔板中的miR-26和NC细胞与0.4 mg萤火虫荧光素酶报告载体和0.08 mg含有海肾萤光素酶(Promega)的pRL-TK对照载体共转染,使用siPORTneoFX(Ambion,Austin,TX),根据使用方法操作,在转染后48 h制备裂解物。使用双光发光报告基因测定(Applied Biosystems)测量荧光素酶活性。将萤火虫荧光素酶活性标准化后测定海肾萤光素酶活性。实验重复3次。
1.6MTT测定将对数期结肠癌细胞使用胰蛋白酶消化后,制成单细胞悬浮液并以1×103细胞/孔的密度接种到96孔板中。在细胞培养7 d后,加入20 μl MTT测定液,每孔充分混合均匀,并在37 ℃下孵育4~6 h。然后使用无菌吸管吸出上清液,每孔加入150 μl二甲基亚砜 (DMSO),在室温下搅拌10 min保证晶体充分溶解。然后在24 h、48 h、72 h和96 h进行MTT测定波长为490 nm时的吸光度,计算结肠癌细胞的增殖情况。实验重复3次。
1.7统计学分析采用SPSS 16.0软件包(SPSS,Chicago,IL)进行统计学分析,P<0.05为差异有统计学意义。使用Wilcoxon秩和检验分析miR-26在结肠癌和配对临近组织中的差异表达。采用单因素方差分析方法评估细胞增殖和侵袭实验三项之间差异,采用最小差别t检验分析。
2 结果
2.1miR-26的表达在结肠癌组织和细胞株中的表达使用qPCR检测60例患者的结肠癌和相邻非肿瘤结肠组织(癌旁组织)中miR-26的表达情况。结果表明,与癌旁组织相比,miR-26的表达在结肠癌组织中明显升高(P<0.05)(见图1)。在4种结肠癌细胞株(HT29、SW480、CW-2和SW620)中观察到miR-26的表达均较高,其中SW480与其他细胞株相比,miR-26表达最高,且SW480的侵袭性相对较强(见图2)。因此,选择了SW480细胞株进行后续研究。
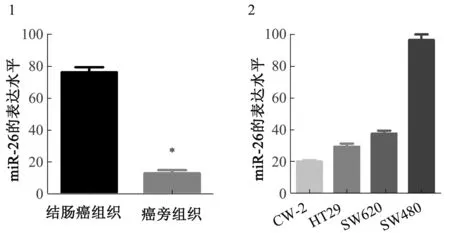
注:与结肠癌组织比较,*P<0.05。
Fig1ExpressionofmiR-26incoloncancertissues;Fig2ExpressionofmiR-26incoloncancercelllines
2.2miR-26在不同临床病理分期结肠癌中的表达情况通过qPCR检测miR-26的表达与结肠癌的病理分期的关系。实验结果显示,结肠癌的病理分期越高,miR-26在结肠癌组织中表达水平越高(见图3)。
2.3miR-26-inhibitor转染结肠癌细胞的转染效率miR-26-inhibitor靶向干扰抑制物由Gnenpharma有限公司合成。miR-26-inhibitor(5′-GGAAGAGCGAUCAAT-3′)。使用对照NC目标序列(5′-UUCUUCGAACCGUTT-3′)作为对照。实验结果显示,重组质粒转染结肠癌细胞株的比例在80%以上(见图4~5)。miR-26-inhibitor转染结肠癌细胞效果良好。

图3 miR-26在不同临床病理分期结肠癌中的表达情况Fig 3 Expression of miR-26 in different clinical pathological stages of colon cancer
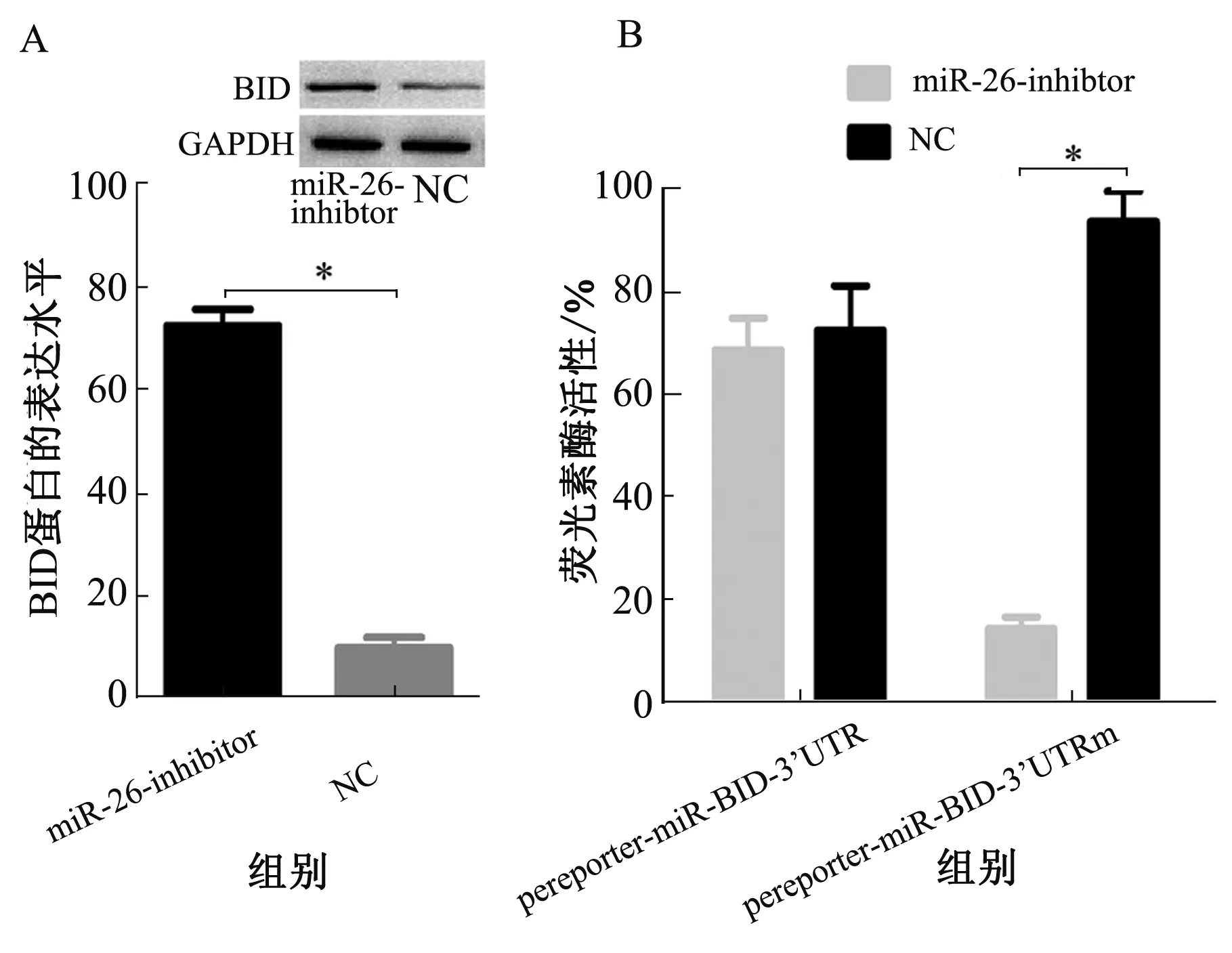
2.4双荧光素酶验证miR-26与BID的关联为了寻找miR-26的下游靶点,利用生物信息学分析发现,miR-26的序列与下游BID基因的部分序列有相同的结合位点。为了验证二者之间是否有调节关系,进行双荧光素酶标记实验。首先,构建了携带3′-UTR的荧光素酶报告因子,其中每个基因的结合位点与miR-26结合位点相对应。荧光素酶测定显示,miR-26下游结合3′-UTR结合位点与BID相符合,而不是其他靶点基因,沉默miR-26过后可以导致荧光素酶活性显著降低(见图6B)。沉默miR-26后可明显上调SW480细胞中BID的表达(见图6A)。
2.5miR-26对结肠癌细胞增殖能力的影响为了研究miR-26对结肠癌细胞增殖能力的影响,使用SW480细胞株进行慢病毒转染以沉默miR-26的表达。通过MTT实验检测miR-26对SW480细胞增殖情况的影响。miR-26-inhibitor组的细胞的生长速度比对照组明显减慢[24 h:(17.2±2.5)%vs(8.1±1.2)%,P<0.05;48 h:(36.2±4.2)%vs(18.4±2.6)%,P<0.05;72 h:(67.5±8.1)%vs(21.4±2.8)%,P<0.01,96 h:(89.2±12.3)%vs(24.4±3.2)%,P<0.05],培养96 h后抑制效果差异有统计学意义(P<0.05,见图7)。抑制miR-26后可以明显抑制SW480结肠癌细胞株的增殖能力。
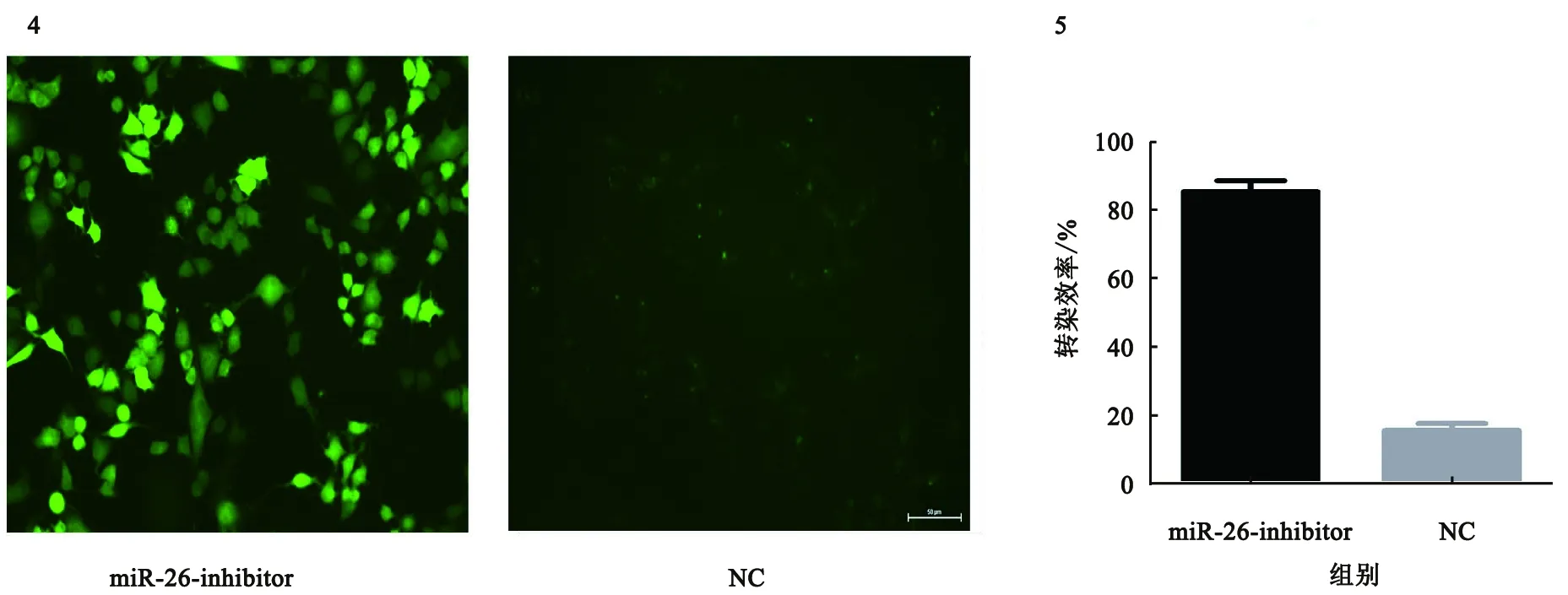
图4 miR-26-inhibitor在结肠癌细胞中的转染效率(400×);图5 miR-26转染效率对比Fig 4 Transfected efficiency of miR-26-inhibitor in colon cancer cells (400×); Fig 5 miR-26 transfected efficiency
注:*P<0.05。
2.6miR-26对结肠癌细胞侵袭能力的影响为了研究miR-26对肝细胞癌侵袭能力的影响,使用SW480细胞株进行慢病毒转染以沉默miR-26的表达。Transwell侵袭实验检测miR-26对SW480细胞侵袭情况的影响。miR-26-inhibitor组细胞的侵袭细胞数目比对照组明显减少[(51.25±8.54)%vs(195.24±11.65)%,P<0.05],差异有统计学意义(见图8)。Transwell侵袭实验结果表明,miR-26-inhibitor组侵袭细胞数目明显少于对照组,细胞侵袭能力明显降低。抑制miR-26后可以明显抑制SW480结肠癌细胞株的侵袭能力。

注:96 h时两组比较,*P<0.05。

注:*P<0.05。
3 讨论
结肠癌是全世界消化系统恶性肿瘤发病率最高的疾病之一,发病率及死亡率仅次于胃癌[14]。有研究[15-16]显示,多种基因蛋白在结肠癌中起重要作用,可以调控结肠癌的发生、发展。然而,对于结肠癌的治疗,目前临床上仍然局限于手术切除及术后化疗,生物靶向治疗尚未在临床上开展实施,所以,探讨结肠癌发生、发展过程中的分子病理机制尤为重要。
结肠癌发生主要与遗传基因和环境因素有很大关联。而部分基因位点的改变可能会导致部分miRNA的表达异常[17]。CLIMENT等[18]通过研究结肠癌组织中miR-26的表达发现,miR-26在结肠癌组织中表达明显上调,且miR-26可通过激活Smad2和STAT3发挥促癌作用。目前有相关研究已经证明miRNA在结肠癌发生、发展中起重要作用,例如miR-21、miR-143和miR-145等[19]。据报道[20],miR-26在结肠癌中上调,因此被认为是一种有促癌作用的miRNA,而且miR-26a通过结合下游相关靶点发挥促进肿瘤细胞的侵袭和扩散。然而,miR-26的下游具体靶点及它们之间是如何作用的,目前尚未见相关研究报道。
miR-26位于19q14.12染色体,有研究[21]表明,miR-26与肿瘤关系密切,一般在肿瘤细胞中表现为促进肿瘤生长的作用。成熟的miR-26一般在乳腺癌、肝癌及结肠癌等组织中呈现高表达。LIU等[22]检测miR-26在乳腺癌中的表达异常,乳腺癌中miR-26表达水平与正常组织相比也出现表达异常,且与乳腺癌的迁移和侵袭能力相关。本研究通过qPCR检测骨肉瘤组织和正常肺组织miR-26的表达发现,miR-26在结肠中表达明显上调,表明miR-26在结肠癌中起促癌作用。结合前面实验结果,通过抑制miR-26的表达后,进一步研究miR-26在结肠癌细胞增殖和侵袭过程中的作用。结果表明,抑制miR-26后,可以抑制结肠癌细胞的增殖和侵袭能力。而miR-26是否是通过调控下游蛋白的表达来调控结肠癌的行为,后续实验我们继续探讨。
Bax家族是凋亡蛋白家族中一种典型蛋白,在许多神经细胞肿瘤和生殖系统中进行过研究[23],Bax可以通过诱导细胞色素c从线粒体中释放,引起细胞膜损伤和细胞凋亡[24]。之前有相关报道,Bax可刺激细胞色素c流出,但其中具体机制尚不明确,在以前的报道中,简单地统计了少数Bax蛋白诱导细胞凋亡和影响增殖的事件,其中包括线粒体构象的改变,但具体的分子机制仍不明确,只是初步认为在线粒体内环境中发生了某种突变[25]。BID是Bax家族中的一种,BID的表达水平可以影响Bax在线粒体外膜上的表达情况[26]。我们之前已经研究过BID可以引起的Bax构象的变化导致其下游N-末端结构域的变化,从而影响其对肿瘤细胞干扰作用[27]。Bax家庭成员BID在调节中细胞色素c释放的过程中发挥重要作用。但是BID能否对miR-26抑制肿瘤细胞的部分生物学功能尚无相关研究。我们实验拟研究BID对结肠癌细胞的影响及其上游可能起总调控作用的miRNA定位,探讨miR-26对BID的调控影响结肠癌的恶性生物学行为。
已知BID是可以受到多种miRNA调控,介导在不同肿瘤细胞中的调控作用。针对BID在细胞中的表达不同,有研究[28]提出,其可能和细胞膜表面受体数量相关,当其表达上调或激活时,可以影响细胞本身的增殖行为。在本研究中,我们检查了miR-26介导BID调控的网络作用的可能性。实验结果表明,miR-26可以调控下游BID基因表达情况,可能是通过调控BID的表达影响结肠癌细胞株的生物学行为。我们的实验观察到BID是miR-26表达调控过程中所必需的。结果表明,在结肠癌中常见的miR-26异常表达可以影响BID的表达,进而调控结肠癌的生物学行为。
本研究系统地研究了miR-26和BID在结肠癌细胞增殖侵袭中的作用和可能机制。BID被鉴定为miR-26的可能新靶标,并被证明是结肠癌细胞增殖所必需的。这些发现将有助于更好地了解结肠癌的分子发病机制,并提示miR-26和BID的类似物可能被认为是结肠癌诊断和治疗过程中的新策略。
[1] O’BRIEN C A, POLLETT A, GALLINGER S, et al. A human colon cancer cell capable of initiating tumour growth in immunodeficient mice [J]. Nature, 2007, 445(7123): 106-110. DOI: 10.1038/nature05372.
[2] ANDERSEN C L, JENSEN J L, ØRNTOFT T F. Normalization of real-time quantitative reverse transcription-PCR data: a model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets [J]. Cancer Res, 2004, 64(15): 5245-5250. DOI: 10.1158/0008-5472.CAN-04-0496.
[3] ZHANG J X, SONG W, CHEN Z H, et al. Prognostic and predictive value of a microRNA signature in stage Ⅱ colon cancer: a microRNA expression analysis [J]. Lancet Oncol, 2013, 14(13): 1295-1306. DOI: 10.1016/S1470-2045(13)70491-1.
[4] TSUJII M, KAWANO S, TSUJI S, et al. Cyclooxygenase regulates angiogenesis induced by colon cancer cells [J]. Cell, 1998, 93(5): 705-716.
[5] AKAO Y, NAKAGAWA Y, NAOE T. let-7 microRNA functions as a potential growth suppressor in human colon cancer cells [J]. Biol Pharm Bull, 2006, 29(5): 903-906.
[6] SARVER A L, FRENCH A J, BORRALHO P M, et al. Human colon cancer profiles show differential microRNA expression depending on mismatch repair status and are characteristic of undifferentiated proliferative states [J]. BMC Cancer, 2009, 9: 401. DOI: 10.1186/1471-2407-9-401.
[7] BU P, CHEN K Y, CHEN J H, et al. A microRNA miR-34a-regulated bimodal switch targets Notch in colon cancer stem cells [J]. Cell Stem Cell, 2013, 12(5): 602-615. DOI: 10.1016/j.stem.2013.03.002.
[8] AKAO Y, NAKAGAWA Y, NAOE T. MicroRNA-143 and -145 in colon cancer [J]. DNA Cell Biol, 2007, 26(5): 311-320. DOI: 10.1089/dna.2006.0550.
[9] TAZAWA H, TSUCHIYA N, IZUMIYA M, et al. Tumor-suppressive miR-34a induces senescence-like growth arrest through modulation of the E2F pathway in human colon cancer cells [J]. Proc Natl Acad Sci U S A, 2007, 104(39): 15472-15477. DOI: 10.1073/pnas.0707351104.
[10] GLOSTEN L R, MILGROM P R. Bid, ask and transaction prices in a specialist market with heterogeneously informed traders [J]. J Financ Econom, 1985, 14: 71-100. DOI: 10.1016/0304-405X(85)90044-3.
[11] PAYNE R. Informed trade in spot foreign exchange markets: an empirical investigation [J]. J Intern Econom, 2003, 61: 307-329. DOI: 10.1016/S0022-1996(03)00003-5.
[12] LUO X, BUDIHARDJO I, ZOU H, et al. Bid, a Bcl2 interacting protein, mediates cytochrome c release from mitochondria in response to activation of cell surface death receptors [J]. Cell, 1998, 94(4): 481-490.
[13] PEREZ E A, ROMOND E H, SUMAN V J, et al. Trastuzumab plus adjuvant chemotherapy for human epidermal growth factor receptor 2-positive breast cancer: planned joint analysis of overall survival from NSABP B-31 and NCCTG N9831 [J]. J Clin Oncol, 2014, 32(33): 3744-3752. DOI: 10.1200/JCO.2014.55.5730.
[14] WANG Z X, CAO J X, LIU Z P, et al. Combination of chemotherapy and immunotherapy for colon cancer in China: a meta-analysis [J]. World J Gastroenterol, 2014, 20(4): 1095-1106. DOI: 10.3748/wjg.v20.i4.1095.
[15] SORIA-VALLES C, GUTIéRREZ-FERNáNDEZ A, GUIU M, et al. The anti-metastatic activity of collagenase-2 in breast cancer cells is mediated by a signaling pathway involving decorin and miR-21 [J]. Oncogene, 2014, 33(23): 3054-3063. DOI: 10.1038/onc.2013.267.
[16] TOIYAMA Y, OKUGAWA Y, TANAKA K, et al. A panel of methylated microRNA biomarkers for identifying high-risk patients with ulcerative colitis-associated colorectal cancer [J]. Gastroenterology, 2017, 153(6): 1634-1646. e8. DOI: 10.1053/j.gastro.2017.08.037.
[17] ZEITELS L R, ACHARYA A, SHI G, et al. Tumor suppression by miR-26 overrides potential oncogenic activity in intestinal tumorigenesis [J]. Genes Dev, 2014, 28(23): 2585-2590. DOI: 10.1101/gad.250951.114.
[18] CLIMENT M, QUINTAVALLE M, MIRAGOLI M, et al. TGFβ triggers miR-143/145 transfer from smooth muscle cells to endothelial cells, thereby modulating vessel stabilization [J]. Circ Res, 2015, 116(11): 1753-1764. DOI: 10.1161/CIRCRESAHA.116.305178.
[19] YU Y, NANGIA-MAKKER P, FARHANA L, et al. miR-21 and miR-145 cooperation in regulation of colon cancer stem cells [J]. Mol Cancer, 2015, 14: 98. DOI: 10.1186/s12943-015-0372-7.
[20] CHEN C Y, CHANG J T, HO Y F, et al. MiR-26 down-regulates TNF-α/NF-κB signalling and IL-6 expression by silencing HMGA1 and MALT1 [J]. Nucleic Acids Res, 2016, 44(8): 3772-3787. DOI: 10.1093/nar/gkw205.
[21] BOREL F, KONSTANTINOVA P, JANSEN P L. Diagnostic and therapeutic potential of miRNA signatures in patients with hepatocellular carcinoma [J]. J Hepatol, 2012, 56(6): 1371-1383. DOI: 10.1016/j.jhep.2011.11.026.
[22] LIU P, TANG H, CHEN B, et al. miR-26a suppresses tumour proliferation and metastasis by targeting metadherin in triple negative breast cancer [J]. Cancer Lett, 2015, 357(1): 384-392. DOI: 10.1016/j.canlet.2014.11.050.
[23] MU Q, LIU P, HU X, et al. Neuroprotective effects of Buyang Huanwu decoction on cerebral ischemia-induced neuronal damage [J]. Neural Regen Res, 2014, 9(17): 1621-1627. DOI: 10.4103/1673-5374.141791.
[24] FICKOVA M, MACHO L, BRTKO J. A comparison of the effects of tributyltin chloride and triphenyltin chloride on cell proliferation, proapoptotic p53, Bax, and antiapoptotic Bcl-2 protein levels in human breast cancer MCF-7 cell line [J]. Toxicol In Vitro, 2015, 29(4): 727-731. DOI: 10.1016/j.tiv.2015.02.007.
[25] ZHU W, WANG H, ZHANG W, et al. Protective effects and plausible mechanisms of antler-velvet polypeptide against hydrogen peroxide induced injury in human umbilical vein endothelial cells [J]. Can J Physiol Pharmacol, 2017, 95(5): 610-619. DOI: 10.1139/cjpp-2016-0196.
[26] ORZECHOWSKA E J, KOZLOWSKA E, CZUBATY A, et al. Controlled delivery of BID protein fused with TAT peptide sensitizes cancer cells to apoptosis [J]. BMC Cancer, 2014, 14: 771. DOI: 10.1186/1471-2407-14-771.
[27] HOCKINGS C, ANWARI K, NINNIS R L, et al. Bid chimeras indicate that most BH3-only proteins can directly activate Bak and Bax, and show no preference for Bak versus Bax [J]. Cell Death Dis, 2015, 6: e1735. DOI: 10.1038/cddis.2015.105.
[28] ROUSALOVA I, BANERJEE S, SANGWAN V, et al. Minnelide: a novel therapeutic that promotes apoptosis in non-small cell lung carcinoma in vivo [J]. PLoS One, 2013, 8(10): e77411. DOI: 10.1371/journal.pone.0077411.
猜你喜欢
天津医科大学学报(2021年4期)2021-08-21
中日友好医院学报(2021年1期)2021-04-14
山东医药(2020年9期)2020-05-20
中成药(2018年3期)2018-05-07
西南国防医药(2016年7期)2016-12-01
腹腔镜外科杂志(2016年10期)2016-06-01
中国卫生标准管理(2015年1期)2016-01-14
医学研究杂志(2015年11期)2015-06-10
中国当代医药(2015年26期)2015-03-01
中国当代医药(2015年16期)2015-03-01
