
不同种源五角枫幼苗叶片N、P化学计量学特征对氮添加的响应
2018-03-08 07:38宋沼鹏何念鹏侯继华
生态学报 2018年1期
宋沼鹏, 何念鹏, 侯继华,*
1 北京林业大学森林资源与生态系统过程北京市重点实验室, 北京 100083 2 中国科学院地理科学与资源研究所生态系统网络观测与模拟重点实验室, 北京 100101
由于化肥和化石燃料的大量使用,越来越多的氮素输入到自然生态系统[1-2]。氮素对自然生态系统的影响以及生态系统的响应受到越来越多的关注。氮素作为植物生长必需的基本营养元素,氮沉降可以改变土壤化学性质、植物的生长、种间竞争、凋落物分解和其他的生态系统过程[3-4]。在氮素缺乏的森林生态系统中,适量的氮沉降能够促进该系统中植物的N吸收效率和光合能力和固碳能力[5],进一步提高林地生产力[6-7]。但是,当过多的氮素被植物吸收,植物体内出现氮饱和的时候,使得植物体内碳分配格局发生变化,抑制光合速率[8]。同时,外界环境持续的高氮沉降,会使树木因缺乏盐基阳离子而降低生产力,最终死亡[9-10],从而导致生物多样性降低,减弱生态系统的功能[11]。因此,植物在不同氮沉降浓度条件下是如何进行响应的,是全球氮沉降对森林生态系统影响的重要课题,仍然是现阶段森林生态学科研究的重点问题。
对于分布广泛的植物物种而言,不同的地方种群长期适应不同的生活环境,形成不同的生态型,这是植物对环境适应的结果[12]。氮沉降过程中,氮素通过多种途径沉降到土壤中,而土壤环境的改变将会直接导致植物的适应策略发生改变[13]。从理论来讲,不同的生态型植物对氮沉降的适应特点也会呈现区域差异。但是到目前为止,不同地区种源个体对氮沉降生态响应策略的变化受到的关注较少。大气环境氮素沉降增加导致土壤氮供给量的改变已经成为影响森林生长和稳定的重要生态因素[14-17]。因此,研究同一物种的不同种源个体对氮沉降的响应是揭示植物对氮沉降响应的有效途径,能够更加深入的了解植物对氮沉降的适应机制,也能够对该物种在不同生长地区的适应策略有更加直观的认识和区分。
五角枫(Acermono)是我国暖温带落叶阔叶林分布广泛的常见物种,具有较高的表型多样性以及遗传多样性,对环境适应能力比较强[18-22]。姚婧[20]通过研究不同林型五角枫叶性状异速生长关系发现不同发育阶段五角枫资源利用方式不同。叶片作为植物制造有机物的重要器官,其功能性状必然对环境的变化极为敏感,土壤环境条件的变化可以使叶片的形态以及化学计量发生变化,从而植物则会调整其自身的资源配置,以适应环境的变化[23-24]。本实验选择北京、山西、内蒙古地区具有明显遗传多样性[22]的五角枫幼苗进行氮添加处理,研究其主要叶片功能性状以及N、P化学计量学特征对不同浓度氮添加的响应机制,试图回答以下科学问题:(1)在氮添加条件下,不同种源的五角枫幼苗的各叶片功能性状以及N、P化学计量学特征对不同氮添加水平如何响应;(2)这种响应趋势是否具有种源差异;(3)响应的差异是否会导致各种源的五角枫幼苗资源利用策略的改变。
1 研究材料与方法
1.1 研究地区自然概况
研究地点位于山西省长治市沁源县灵空山自然保护区国家林业局太岳山森林生态系统定位站(112°07′E,36°38′N)。该地区属暖温带大陆性季风气候,四季分明春季干旱多风,夏季降水集中,年均温在9.3—12.3℃,年均降雨量662mm(7、8、9月雨量集中,超过全年降雨量的60%),年均相对湿度60%—65%,年均日照2600h,年平均无霜期110—125d。土壤多为褐土和棕壤。森林类型主要以暖温带落叶阔叶林为主[25-26]。
1.2 试验材料
选取北京(BW)、山西(SW)、内蒙古(NW)3个种源的3年生五角枫移栽幼苗为试验对象。幼苗分别选自北京风沙源育苗中心、山西省太岳山灵空山林场、内蒙古赤峰市松山区安林苗圃,从当地天然林区收集种子经人工培育而成[27]。
1.3 氮添加水平的设置
中国自然的氮沉降呈现从东南到西北递减的梯度。对五角枫幼苗进行模拟氮沉降的控制实验也必须遵循自然条件下的氮沉降水平。监测数据表明,近年来内蒙古地区的氮沉降约为20kg hm-2a-1,北京地区约为30kg hm-2a-1,山西地区约为40—45kg hm-2a-1[28]。根据不同地区的氮沉降量,同时参照北美Harvard Forest的高浓度氮添加实验[29]以及Bobbink等的低浓度氮添加实验[30],本研究设置对照(N0,0kg hm-2a-1)、低氮(N1,15kg hm-2a-1)、中氮(N2, 25kg hm-2a-1)、高氮(N3, 50kg hm-2a-1)、超高氮(N4, 150kg hm-2a-1) 5个氮添加梯度。
1.4 实验方法
1.4.1 实验设计
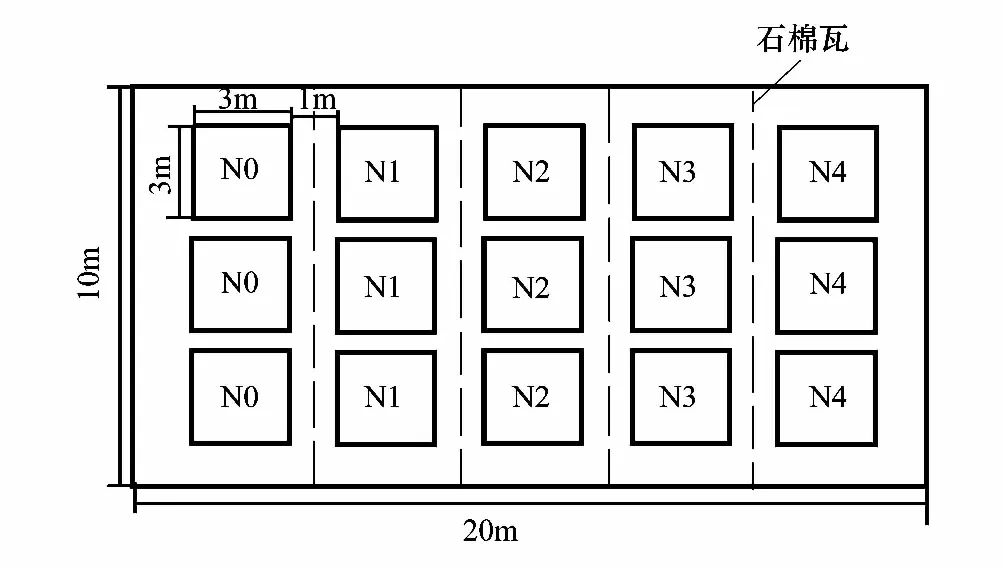
图1 不同种源五角枫氮添加实验样地设计图(引自蒋思思 [27])Fig.1 The design plot of different provenances Acer mono under nitrogen deposition experimental[27]N0:对照,0kg hm-2 a-1; N1:低氮,15kg hm-2 a-1;N2:中氮,25kg hm-2 a-1;N3:高氮,50kg hm-2 a-1;N4:超高氮,150kg hm-2 a-1N0:Control,0kg hm-2 a-1; N1: low nitrogen,15kg hm-2 a-1;N2: middle nitrogen,25kg hm-2 a-1;N3:high nitrogen,50kg hm-2 a-1;N4:the supersaturated nitrogen,150kg hm-2 a-1
在太岳山森林生态系统定位站以南200m,选取合适大小为(10m×20m)的平坦试验样地。由于实验设计为5—6年的氮添加实验,而最初种植时五角枫幼苗根系长度小于20cm,故50cm的原土深度满足实验要求。首先移除样地内深50cm的土壤,将取自林区林下的土壤过筛、充分混匀后,回填到样地内。林下土壤的有机质含量为(20.91±2.30)g/kg,全氮含量为(1.15±0.03)g/kg,全磷含量为(0.61±0.02)g/kg。幼苗的栽种密度参照刘史力油松人工林幼苗抚育实验[31],人工林种植密度满足10株/10m2,幼苗株间距定为50cm。因此在满足实验重复的前提下,设置每个小样方面积为3m×3m。样地进一步划分为15个3m×3m的小样方,小样方间隔1m。为了避免相邻样方的植物根系受不同的氮添加水平影响,小样方之间用石棉瓦隔开,石棉瓦深度为50cm,具体样地设计图见图1。2011年10月将3个不同种源的五角枫幼苗做好标记后,移栽至样地内随机混植,每个小样方中每个种源的幼苗各6株,株间距为50cm。
自2012年6月起,每月将相应质量的NH4NO3溶于3L水中再分别向每个小样方中的地面均匀喷施(N0处理喷施等量的水,样地中其他的水均来源于自然降雨)。
1.4.2 样品采集
2015年9月底,在每个小样方内随机选取3个种源的五角枫幼苗各3株进行样品采集,即每个氮添加水平下每个种源的五角枫幼苗共9株作为重复。采集每株五角枫幼苗接近顶端的倒数第3枝条,枝、叶进行分离,叶片装入自封袋中称量鲜重并于当天测量叶面积,擦净叶片,剪掉叶柄,将每株所采叶片用HP Scanjet G3110扫描仪进行扫描,用ImageJ软件计算叶片的叶面积,之后在80℃的温度下烘干24h至恒重,称其干重,并测定氮磷化学计量学指标。
1.4.3 主要功能性状指标及氮磷化学计量特征的测定
叶片干物质含量LDMC/(g/kg)= 叶片干重/叶片鲜重,
LDMC变化量/(g/kg)= 各氮水平处理(N1、N2、N3、N4)的LDMC-N0的LDMC;
比叶面积SLA /(m2/kg)= 叶面积/叶干重,
SLA变化量/(g/kg)= 各氮水平处理(N1、N2、N3、N4)的SLA-N0的SLA。
将烘干的叶片进行研磨,过5mm筛,用分析天平称取样品5g,用硫酸纸送至消煮管底部,植物样品的消煮采用H2SO4-H2O2消煮法,将消煮液过滤转移入100mL容量瓶中,用蒸馏水定容保存,N、P的测定用Seal Analytical AA3(德国)连续流动分析仪。
氮、磷面积浓度(Narea, Parea) =氮、磷质量浓度(Nmass, Pmass)×单位面积的干物质含量。
1.4.4 数据处理
叶性状数值由算术平均值和标准误差来表示。采用单因素方差分析(one-way ANOVA)来检验模拟氮沉降对植物功能性状产生的影响,采用Fisher LSD法分析各性状在不同处理间是否差异显著(α=0.05)。用 Pearson 相关性分析计算叶片性状间的相关性,双因素(Univariate two-way)方差分析检验种源和模拟氮沉降处理对植物性状的交互作用,统计分析用(SPSS Inc.,USA)及R完成。
2 结果与分析
2.1 氮添加对不同种源五角枫幼苗的比叶面积与干物质含量的影响
不同氮添加处理下,内蒙古种源五角枫幼苗(NW)和山西种源五角枫幼苗(SW)的LDMC表现出较为一致的响应趋势,即氮添加条件下幼苗的LDMC均显著低于对照,分别平均降低11.15%,13.99%,同时在超高氮(N4)的条件下幼苗的LDMC显著高于低氮、中氮和高氮处理(P<0.01)。同时,NW幼苗的LDMC含量普遍比SW低。北京种源幼苗(BW)的LDMC在各氮添加处理下变化不显著,在低、超高浓度氮添加下表现为增加,但差异不显著(P>0.05)(图2a,表1)。
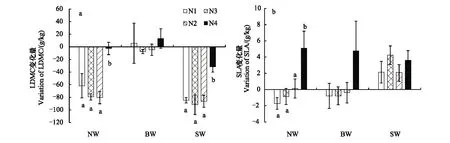
图2 氮添加处理下不同种源五角枫的叶片干物质含量(LDMC)及比叶面积(SLA)的变化规律Fig.2 Variations between LDMC and N0 and SLA and N0 of three provenance Acer mono seedlings under different nitrogen additionLDMC:叶片干物质含量, Leaf dry matter content;SLA:比叶面积,Specific leaf area;NW:内蒙古种源五角枫幼苗,Inner Mongolia provenances A. mono seedlings;BW:北京种源五角枫幼苗,Beijng provenances A. mono seedlings; SW:山西种源五角枫幼苗,Shanxi provenances A.mono seedlings
3个种源五角枫幼苗的比叶面积(SLA)与对照的差值在减小,说明对氮添加的响应总体上呈增加趋势,NW,SW和BW分别增加4.16%、21.51%、4.67%。随着氮添加浓度的增加,NW与BW的SLA表现为先降低后增加,并在N4条件下显著增加,并达到最大(P<0.05);SW的SLA的在氮添加条件下增加,在N2的水平下达到最大后降低,处理间差异显著(P<0.05),SW的平均SLA大于其他两个种源幼苗(图2b,表1)。
2.2 氮添加对五角枫叶片N、P化学计量学特征的影响
不同种源的五角枫幼苗的叶片氮质量浓度Nmass、磷质量浓度Pmass、氮面积浓度Narea、磷面积浓度Parea以及N∶P,呈现不同的变化趋势(表2)。氮添加处理下SW种源幼苗叶片的Pmass显著增加,并在低氮(N1)水平最大(P<0.05),Nmass在N1和N3水平显著高于对照(P<0.05)。N∶P在N1水平表现为降低,N2、N3水平表现为增加,但是变化均不显著。NW和BW幼苗叶片的N、P化学计量学特征变化均不显著,但是Nmass和Narea在中氮和中高氮水平下表现为增加趋势,而Pmass和Parea在中氮和中高氮水平下表现为降低趋势,同时N∶P在低氮水平表现为降低,高氮水平表现为增加,这与N、P浓度的变化趋势相近。
表1氮添加处理对不同种源五角枫的叶片干物质含量和比叶面积影响的单因素方差分析及多重比较(均值±标准误,n=9)
Table1SinglefactorvarianceanalysisandLSDoftheeffectofnitrogenadditionontheLDMCandSLAofthreeprovenanceAcermonoseedlings(mean ± SE,n=9)
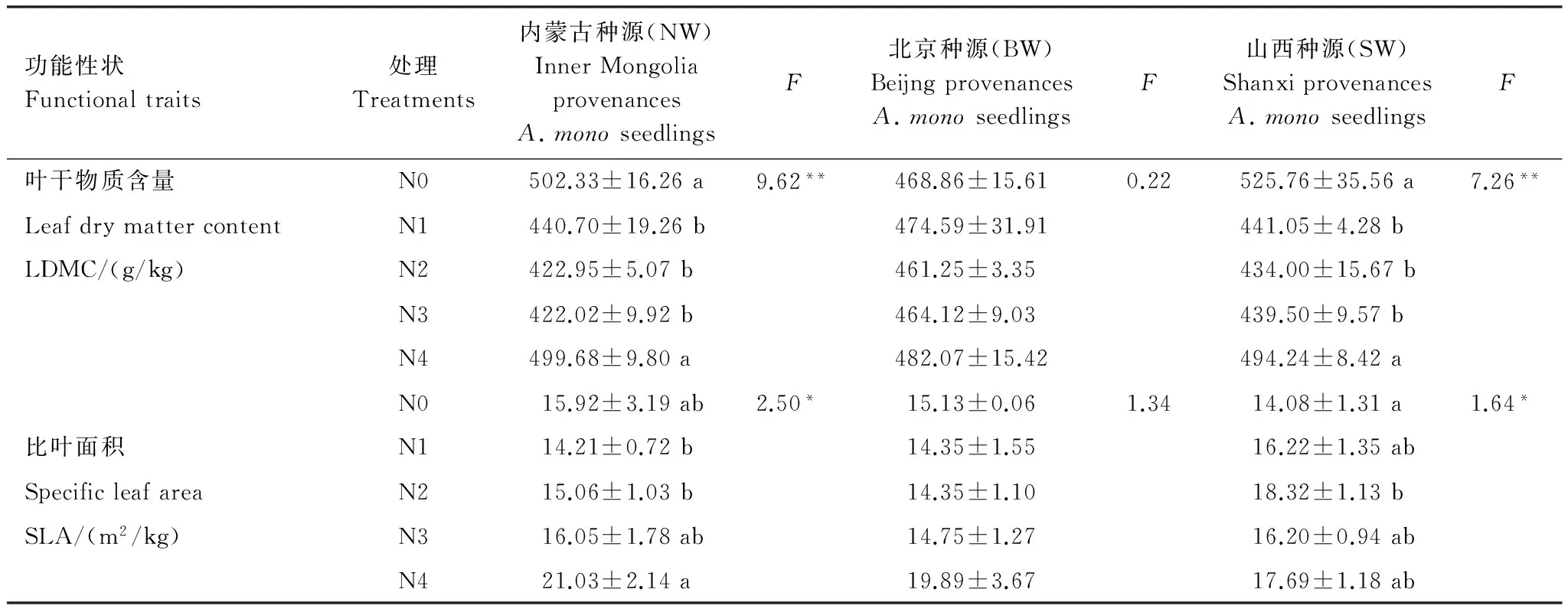
功能性状Functionaltraits处理Treatments内蒙古种源(NW)InnerMongoliaprovenancesA.monoseedlingsF北京种源(BW)BeijngprovenancesA.monoseedlingsF山西种源(SW)ShanxiprovenancesA.monoseedlingsF叶干物质含量N0502.33±16.26a9.62**468.86±15.610.22525.76±35.56a7.26**LeafdrymattercontentN1440.70±19.26b474.59±31.91441.05±4.28bLDMC/(g/kg)N2422.95±5.07b461.25±3.35434.00±15.67bN3422.02±9.92b464.12±9.03439.50±9.57bN4499.68±9.80a482.07±15.42494.24±8.42aN015.92±3.19ab2.50*15.13±0.061.3414.08±1.31a1.64*比叶面积N114.21±0.72b14.35±1.5516.22±1.35abSpecificleafareaN215.06±1.03b14.35±1.1018.32±1.13bSLA/(m2/kg)N316.05±1.78ab14.75±1.2716.20±0.94abN421.03±2.14a19.89±3.6717.69±1.18ab
同列不同小写字母表示不同处理间差异显著(P<0.05);F检验值表示该组处理差异显著,*:P<0.05;**:P<0.01
表2氮添加对不同种源五角枫的化学计量学指标的单因素方差分析及变异系数(均值±标准误,n=9)
Table2SinglefactorvarianceanalysisandcoefficientofvariationoftheeffectofnitrogenadditiononthestoichiometriccharacteristicsofthreeprovenanceAcermonoseedlings(mean±SE,n=9)
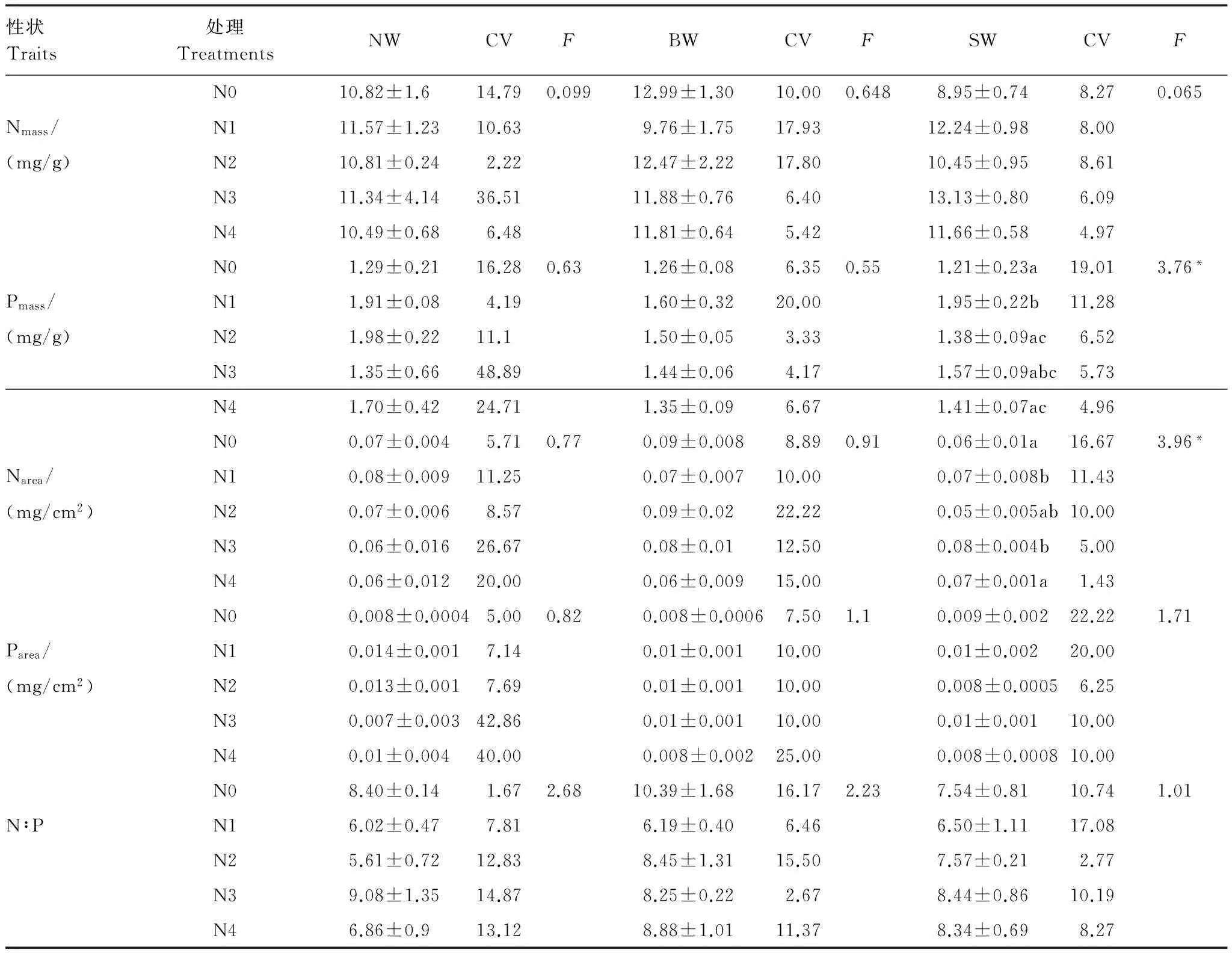
性状Traits处理TreatmentsNWCVFBWCVFSWCVFN010.82±1.614.790.09912.99±1.3010.000.6488.95±0.748.270.065Nmass/N111.57±1.2310.639.76±1.7517.9312.24±0.988.00(mg/g)N210.81±0.242.2212.47±2.2217.8010.45±0.958.61N311.34±4.1436.5111.88±0.766.4013.13±0.806.09N410.49±0.686.4811.81±0.645.4211.66±0.584.97N01.29±0.2116.280.631.26±0.086.350.551.21±0.23a19.013.76*Pmass/N11.91±0.084.191.60±0.3220.001.95±0.22b11.28(mg/g)N21.98±0.2211.11.50±0.053.331.38±0.09ac6.52N31.35±0.6648.891.44±0.064.171.57±0.09abc5.73N41.70±0.4224.711.35±0.096.671.41±0.07ac4.96N00.07±0.0045.710.770.09±0.0088.890.910.06±0.01a16.673.96*Narea/N10.08±0.00911.250.07±0.00710.000.07±0.008b11.43(mg/cm2)N20.07±0.0068.570.09±0.0222.220.05±0.005ab10.00N30.06±0.01626.670.08±0.0112.500.08±0.004b5.00N40.06±0.01220.000.06±0.00915.000.07±0.001a1.43N00.008±0.00045.000.820.008±0.00067.501.10.009±0.00222.221.71Parea/N10.014±0.0017.140.01±0.00110.000.01±0.00220.00(mg/cm2)N20.013±0.0017.690.01±0.00110.000.008±0.00056.25N30.007±0.00342.860.01±0.00110.000.01±0.00110.00N40.01±0.00440.000.008±0.00225.000.008±0.000810.00N08.40±0.141.672.6810.39±1.6816.172.237.54±0.8110.741.01N∶PN16.02±0.477.816.19±0.406.466.50±1.1117.08N25.61±0.7212.838.45±1.3115.507.57±0.212.77N39.08±1.3514.878.25±0.222.678.44±0.8610.19N46.86±0.913.128.88±1.0111.378.34±0.698.27
不同小写字母表示不同处理间存在显著差异(P<0.05);Nmass:氮质量浓度,Nitrogen mass concentration; Pmass:磷质量浓度,Phosphorus mass concentration; Narea: 氮面积浓度,Nitrogen area concentration; Parea:磷面积浓度,Phosphorus area concentration; N∶P:氮磷比,N-P ratio; NW:内蒙古种源五角枫幼苗,Inner Mongolia provenancesA.monoseedlings; BW:北京种源五角枫幼苗,Beijng provenancesA.monoseedlings; SW:山西种源五角枫幼苗,Shanxi provenancesA.monoseedlings; CV:变异系数,Coefficient of Variance
2.3 五角枫叶片功能性状变异来源的相关性分析
为探究五角枫幼苗LDMC与SLA性状及氮磷化学计量学特征发生变异的来源,对实验数据进行氮添加处理和种源的双因素方差分析及相关性检验(表3)。结果表明:SLA、LDMC、Parea及N∶P与氮添加处理呈显著的相关关系(P< 0.05),与种源无显著的相关关系。同时五角枫幼苗的SLA、LDMC、Nmass、Pmass、Narea、Parea和N∶P对氮添加处理和种源的交互作用均未表现出相关关系。说明五角枫幼苗的这些性状变化主要受氮添加处理影响。
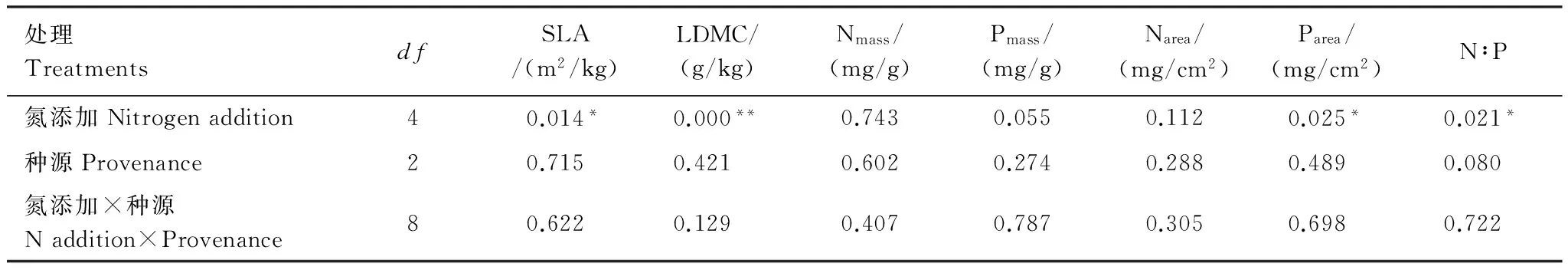
表3 氮添加和种源对幼苗生长指标影响效应的双方差分析Table 3 Variance analysis of the growth index effect in Processing and provenance of Acer mono seedling
*,P< 0.05; **,P< 0.01; LDMC:叶干物质含量,Leaf dry matter content; SLA比叶面积,Specific leaf area; Nmass:氮质量浓度,Nitrogen mass concentration; Pmass:磷质量浓度,Phosphorus mass concentration; Narea: 氮面积浓度,Nitrogen area concentration; Parea:磷面积浓度,Phosphorus area concentration; N∶P:氮磷比,N-P ratio
3 讨论
叶片作为木本植物制造有机物的主要器官,与植物的有机物生产过程密切相关,并且在不同生境条件下和不同物种之间存在很大的差异性[32-33]。在本研究中,NW和SW叶片的LDMC含量在氮添加条件下比BW低,低中浓度的氮添加条件下表现为抑制作用,在N4条件下表现为显著的促进作用。同时,BW的LDMC含量在N1、N2、N3条件下均高于同水平下的SW和NW。氮添加条件下,SW的SLA增加程度最大。有研究指出资源相对丰富的环境中植物的SLA更大, 植物个体对资源的利用、光合和生长能力更强,而在贫瘠的环境中SLA小的植物适应能力更强[34-35]。山西种源五角枫幼苗(SW)原生境的环境资源相对丰富,对高氮环境有更好的资源利用能力,更能适应氮沉降量相对较多的环境。该结论与SW原生境地区氮沉降背景相一致[28]。将SLA和LDMC综合考虑,一般认为SLA减小的同时LDMC增加表示植物所处环境条件的恶化,此时植物倾向于适应保守的生长策略[36,34]。就资源获取方面,Rose等认为具有低SLA和高LDMC的植物,其资源获取能力以及水分的利用效率往往更高,更容易适应恶劣的环境条件[37]。土壤N浓度的改变,使山西种源五角枫幼苗资源利用方式发生变化,使其倾向保守的生长策略,即对环境变化较为灵活的生长策略。因此可以看出,在氮添加条件下,SW比BW和NW有更强的环境资源利用能力,并且其在生长过程中逐步采取较为保守的生长方式,相比而言NW在高氮沉降的环境中的资源利用能力最差。因此氮沉降改变了五角枫幼苗的适应策略。在植物根、茎、叶各生理活动的运作过程中,碳、氮、磷各元素之间达到相互平衡,尤其是植物叶片氮磷相互作用,能对外界环境的变化做出积极的响应,因此根据植物叶片氮磷含量判断生境营养状况是生态化学计量学的重要部分[38]。相比对照而言,氮添加条件下3个种源五角枫的Nmass、Narea、Pmass和Parea均表现为增加,这与众多研究的结果一致,均认为适量的氮添加能够增加植物叶片的TN、TP[7,39]。在不同浓度的氮添加下,只有SW的Pmass在N1水平下显著增加,Nmass在N1和N3水平下表现为显著增加。说明在低氮水平下, SW对P的吸收加强,从而增加植物体内对P的积累。同时在低氮和中高氮条件下, SW对氮的吸收加强,说明SW能较好的适应低氮和高氮环境,使其生存能力更强,能够适应不同的环境条件。同时比较N、P变化情况可以发现,随着氮添加浓度的升高,N、P值在中氮或者高氮水平达到最大值后开始下降,说明一定范围之内外源氮输入对于植物生长的起到促进作用,而超过了这个阈值,植物的生长就会出现抑制现象[40,9]。
植物叶片的N∶P从某种程度上能够直观的反映植物生长受N或P的限制情况。Güsewell认为N∶P<10和>20可作为植被水平氮、磷限制的评价指标[41]。通过表2中N∶P的变异系数可以看出,N∶P的变异系数小于N、P,也说明N∶P对植物体生长存在一定的关联。早期通过多种施肥实验总结得出,当植物叶片N∶P > 16表示植物生长主要受P限制,而叶片N∶P < 14时,主要受N限制, 当 N∶P位于14到16之间时,则植物生长受到二者的共同限制[42]。在本研究中,五角枫幼苗的N∶P<14,说明研究的五角枫幼苗的生长均受到N的限制,而对于不同种源的五角枫幼苗,在氮添加条件下受限制程度不同。可以看出,SW的N∶P在氮添加条件下增加,施氮有利于SW的N积累。同样在石贤萌等对哀牢山两种湿性常绿阔叶树种幼苗的研究中得出[43],氮添加可以增加植物N含量。而NW和BW比对照有所降低,可能原因是由于氮添加促进其对P素的吸收。有氮添加控制实验[44]发现,氮沉降有利于降低植物N∶P,而主要原因是由于植物体内P的增加,可能原因是多余的氮素能够促进陆地生态系统中P的循环速率,从而促进植物对P的吸收[45]。而目前植物P含量对氮添加的响应存在不同的规律,李德军等[40]及Macklon等[46]地研究说明,氮沉降使土壤有效N增加,引起土壤酸化,在增加植物体内氮的同时,抑制幼苗对P的吸收,使N∶P增加。在本研究中,氮添加促进实验样地中五角枫对P的吸收,使得不同种源五角枫叶P比叶N增加更多,可能是导致N∶P降低的原因。而对于本实验来说,氮添加如何促进土壤P的循环以及寻找五角枫受N限制的其他原因,还需要进一步探究。
根据叶经济谱理论,LDMC、SLA、叶N含量、叶P含量以及N∶P是考量植物对环境资源权衡策略的部分重要指标[47]。在一些研究中指出,有较低LDMC、较高SLA、叶N含量和叶P含量的树种通常具有较高的光合速率和生长速率,具有这些功能性状变化的植物往往属于“快速投资-收益”型,反之则倾向于“缓慢投资-收益”型[48]。本研究中,在氮添加的条件下,NW的LDMC显著降低,SLA显著增大,同时叶N含量、叶P含量以及N∶P比较高;SW的LDMC显著降低,但均值比NW大,SLA增大,同时叶N含量、叶P含量表现为显著增大;BW的LDMC和SLA较其他两个种源的五角枫也有所增加,但变化不显著,同时叶N含量、叶P含量以及N∶P含量比较低。由此可见, SW和NW属于“快速投资-收益”型,而BW倾向于“缓慢投资-收益”型。生存环境发生变化,植物会对自身资源进行调整,从而优化配置,使自身适应新的环境[49]。因此,在氮添加下,五角枫在适应周围环境的过程中,不同种源的五角枫的适应策略发生了改变。从变异来源分析得出,各种源五角枫适应策略的改变主要来源于氮添加处理,也说明氮沉降确实改变了五角枫的适应性。而与种源没有显著的关系,可能原因是由于物种的不同,不同的物种在种源的适应性方面存在一定的差异。所以对于五角枫而言,种源不是五角枫发生适应性差异的主要原因。另外,本实验选取的部分功能指标变化未显示出与种源的显著关系,其他的指标研究有待于在下一步进行的工作中进行。
综上,不论是从SLA和LDMC的结果,还是N、P化学计量学指标的结果均说明,在氮添加条件下,不同种源的五角枫表现出较为显著的差异,证明不同种源的五角枫对氮沉降具有差异性的适应策略。所以,五角枫资源的保护利用以及引种栽培,均要充分考虑五角枫的种源差异,才能做出更加合理的方案和决策。
[1] Galloway J N. The global nitrogen cycle: past, present and future. Science in China Series C: Life Sciences, 2005, 48(S2): 669-678.
[2] Niu S L, Wu M Y, Han Y, Xia J Y, Zhang Z, Yang H J, Wan S Q. Nitrogen effects on net ecosystem carbon exchange in a temperate steppe. Global Change Biology, 2010, 16(1): 144-155.
[3] Niu S L, Classen A T, Dukes J S, Kardol P, Liu L L, Luo Y Q, Rustad L, Sun J, Tang J W, Templer P H, Thomas R Q, Tian D S, Vicca S, Wang Y P, Xia J Y, Zaehle S. Global patterns and substrate-based mechanisms of the terrestrial nitrogen cycle. Ecology Letters, 2016, 19(6): 697-709.
[4] 肖迪, 王晓洁, 张凯, 何念鹏, 侯继华. 氮添加对山西太岳山天然油松林主要植物叶片性状的影响. 植物生态学报, 2016, 40(7): 686-701.
[5] 李林森, 程淑兰, 方华军, 于贵瑞, 徐敏杰, 王永生, 党旭升, 李英年. 氮素富集对青藏高原高寒草甸土壤有机碳迁移和累积过程的影响. 土壤学报, 2015, 52(1): 183-193.
[6] Liu P, Huang J H, Sun O J, Han X G. Litter decomposition and nutrient release as affected by soil nitrogen availability and litter quality in a semiarid grassland ecosystem. Oecologia, 2010, 162(3): 771-780.
[7] 黄菊莹, 余海龙. 四种荒漠草原植物的生长对不同氮添加水平的响应. 植物生态学报, 2016, 40(2): 165-176.
[8] Berger T W, Glatzel G. Response ofQuercuspetraeaseedlings to nitrogen fertilization. Forest Ecology and Management, 2001, 149(1/3): 1-14.
[9] Bauer G A, Bazzaz F A, Minocha R, Long S, Magill A, Aber J, Berntson G M. Effects of chronic N additions on tissue chemistry, photosynthetic capacity, and carbon sequestration potential of a red pine (PinusresinosaAit.) stand in the NE United States. Forest Ecology and Management, 2004, 196(1): 173-186.
[10] 袁颖红, 樊后保, 李燕燕, 刘文飞, 廖迎春, 沈芳芳, 黄荣珍. 模拟氮沉降对土壤酸化和土壤盐基离子含量的影响. 应用与环境生物学报, 2011, 17(4): 461-466.
[11] Stark S, Männistö M K, Eskelinen A. Nutrient availability and pH jointly constrain microbial extracellular enzyme activities in nutrient-poor tundra soils. Plant and Soil, 2014, 383(1/2): 373-385.
[12] 李庆康, 马克平. 植物群落演替过程中植物生理生态学特性及其主要环境因子的变化. 植物生态学报, 2002, 26(增刊): 9-19.
[13] Yang J Y, Fan J. Review of study on mineralization, saturation and cycle of nitrogen in forest ecosystems. Journal of Forestry Research, 2003, 14(3): 239-243.
[14] Galloway J N, Dentener F J, Capone D G, Boyer E W, Howarth R W, Seitzinger S P, Asner G P, Cleveland C C, Green P A, Holland E A, Karl D M, Michaels A F, Porter J H, Townsend A R, Vöosmarty C J. Nitrogen cycles: past, present, and future. Biogeochemistry, 2004, 70(2): 153-226.
[15] Verhoeven J T A, Koerselman W, Meuleman A F M. Nitrogen-or phosphorus-limited growth in herbaceous, wet vegetation: relations with atmospheric inputs and management regimes. Trends in Ecology & Evolution, 1996, 11(12): 494-497.
[16] Clark C M, Tilman D. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature, 2008, 451(7179): 712-715.
[17] Liu X J, Zhang Y, Han W X, Tang A H, Shen J L, Cui Z L, Vitousek P, Erisman J W, Goulding K, Christie P, Fangmeier A, Zhang F S. Enhanced nitrogen deposition over China. Nature, 2013, 494(7438): 459-462.
[18] 李岩, 董吉成, 张惠君, 徐辉. 色木槭播种育苗技术探讨. 绿色科技, 2011, (12): 107-107.
[19] 郑万钧. 中国树木志(第4卷). 北京: 中国林业出版社, 2004: 4258-4259.
[20] 姚婧, 李颖, 魏丽萍, 蒋思思, 杨松, 侯继华. 东灵山不同林型五角枫叶性状异速生长关系随发育阶段的变化. 生态学报, 2013, 33(13): 3907-3915.
[21] 张翠琴, 姬志峰, 林丽丽, 赵瑞华, 王祎玲. 五角枫种群表型多样性. 生态学报, 2015, 35(16): 5343-5352.
[22] 张翠琴. 五角枫种群遗传多样性研究[D]. 临汾: 山西师范大学, 2015.
[23] Scoffoni C, Rawls M, McKown A, Cochard H, Sack L. Decline of leaf hydraulic conductance with dehydration: relationship to leaf size and venation architecture. Plant Physiology, 2011, 156(2): 832-843.
[24] Cornelissen J H C, Cerabolini B, Castro-Díez P, Villar-Salvador P, Montserrat-Martí G, Puyravaud J P, Maestro M, Werger M J A, Aerts R. Functional traits of woody plants: correspondence of species rankings between field adults and laboratory-grown seedlings? Journal of Vegetation Science, 2003, 14(3): 311-322.
[25] 朱建奎. 山西太岳山地区森林土壤理化性状研究[D]. 北京: 北京林业大学, 2009.
[26] 姚雪芹, 毕润成, 张钦弟, 蒋鹏飞. 山西太岳山辽东栎群落木本植物功能群分类. 西北植物学报, 2015, 35(6): 1246-1253.
[27] 蒋思思, 魏丽萍, 杨松, 肖迪, 王晓洁, 侯继华. 不同种源油松幼苗的光合色素和非结构性碳水化合物对模拟氮沉降的短期响应. 生态学报, 2015, 35(21): 7061-7070.
[28] 顾峰雪, 黄玫, 张远东, 闫慧敏, 李洁, 郭瑞, 钟秀丽. 1961—2010年中国区域氮沉降时空格局模拟研究. 生态学报, 2016, 36(12): 3591-3600.
[29] Magill A H, Aber J D, Currie W S, Nadelhoffer K J, Martin M E, McDowell W H, Melillo J M, Steudler P. Ecosystem response to 15 years of chronic nitrogen additions at the Harvard Forest LTER, Massachusetts, USA. Forest Ecology and Management, 2004, 196(1): 7-28.
[30] Bobbink R, Hicks K, Galloway J, Spranger T, Alkemade R, Ashmore M, Bustamante M, Cinderby S, Davidson E, Dentener F, Emmett B, Erisman J W, Fenn M, Gilliam F, Nordin A, Pardo L, De Vries W. Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecological Applications, 2010, 20(1): 30-59.
[31] 刘史力. 黄土高原人工种植油松幼苗的生长特性[D]. 西安: 陕西师范大学, 2013.
[32] Milla R, Reich P B. The scaling of leaf area and mass: the cost of light interception increases with leaf size. Proceedings of the Royal Society B: Biological Sciences, 2007, 274(1622): 2109-2115.
[33] 张慧文, 马剑英, 孙伟, 陈发虎. 不同海拔天山云杉叶功能性状及其与土壤因子的关系. 生态学报, 2010, 30(21): 5747-5758.
[34] Wilson P J, Thompson K, Hodgson J G. Specific leaf area and leaf dry matter content as alternative predictors of plant strategies. New Phytologist, 1999, 143(1): 155-162.
[35] 祁建, 马克明, 张育新. 北京东灵山不同坡位辽东栎(Quercusliaotungensis)叶属性的比较. 生态学报, 2008, 28(1): 122 -128.
[36] Grime J P, Thompson K, Hunt R, Hodgson J G, Cornelissen J H C, Rorison I H, Hendry G A F, Ashenden T W, Askew A P, Band S R, Booth R E, Bossard C C, Campbell B D, Cooper J E L, Davison A W, Gupta P L, Hall W, Hand D W, Hannah M A, Hillier S H, Hodkinson D J, Jalili A, Liu Z, Mackey J M L, Matthews N, Mowforth M A, Neal A M, Reader R J, Reiling K, Ross-Fraser W, Spencer R E, Sutton F, Tasker D E, Thorpe P C, Whitehouse J. Integrated screening validates primary axes of specialisation in plants. Oikos, 1997, 79(2): 259-281.
[37] Rose L, Rubarth M C, Hertel D, Leuschner C. Management alters interspecific leaf trait relationships and trait-based species rankings in permanent meadows. Journal of Vegetation Science, 2013, 24(2): 239-250.
[38] 曾德慧, 陈广生. 生态化学计量学: 复杂生命系统奥秘的探索. 植物生态学报, 2005, 29(6): 1007-1019.
[39] 黄菊莹, 袁志友, 李凌浩. 羊草绿叶氮、磷浓度和比叶面积沿氮、磷和水分梯度的变化. 植物生态学报, 2009, 33(3): 442-448.
[40] 李德军, 莫江明, 彭少麟, 方运霆. 南亚热带森林两种优势树种幼苗的元素含量对模拟氮沉降增加的响应. 生态学报, 2005, 25 (9): 2165-2172.
[41] Güsewell S. N∶P ratios in terrestrial plants: Variation and functional significance. New Phytologist, 2004, 164(2): 243-266.
[42] Koerselman W, Meuleman A F M. The vegetation N∶P ratio: a new tool to detect the nature of nutrient limitation. Journal of Applied Ecology, 1996, 33(6): 1441-1450.
[43] 石贤萌, 杞金华, 宋亮, 刘文耀, 黄俊彪, 李苏, 卢华正, 陈曦. 哀牢山中山湿性常绿阔叶林两种优势幼苗C、N、P化学计量特征及其对N沉降增加的响应.植物生态学报, 2015, 39(10): 962-970.
[44] Liu J X, Huang W J, Zhou G Y, Zhang D Q, Liu S Z, Li Y Y. Nitrogen to phosphorus ratios of tree species in response to elevated carbon dioxide and nitrogen addition in subtropical forests. Global Change Biology, 2013, 19(1): 208-216.
[45] Marklein A R, Houlton B Z. Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytologist, 2012,193(3): 696-704.
[46] Macklon A E S, Sim A. Modifying effects of non-toxic levels of Aluminium on the uptake and transport of phosphate in Ryegrass. Journal of Experimental Botany, 1992, 43(252): 915-923.
[47] Wright I J, Reich P B, Westoby M, Ackerly D D, Baruch Z, Bongers F, Cavender-Bares J, Chapin T, Cornelissen J H C, Diemer M, Flexas J, Garnier E, Groom P K, Gulias J, Hi-kosaka K, Lamont B B, Lee T, Lee W, Lusk C, Midgley J J, Navas M L, Niinemets Ü, Oleksyn J, Osada N, Poorter H, Poot P,Prior L,Pyankov V I, Roumet C, Thomas S C, Tjoelker M G, Veneklaas E J, Villar R. The world-wide leaf economics spectrum. Nature, 2004, 428(6985): 821-827.
[48] Kikuzawa K. A cost-benefit analysis of leaf habit and leaf longevity of trees and their geographical pattern. The American Naturalist, 1991,138(5):1250-1263.
[49] Wright J P, Sutton-Grier A. Does the leaf economic spectrum hold within local species pools across varying environmental conditions? Functional Ecology, 2012, 26(6): 1390-1398.
猜你喜欢
特产研究(2022年6期)2023-01-17
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
辽宁林业科技(2021年1期)2021-03-16
现代园艺(2020年12期)2020-12-20
新疆农业科学(2020年1期)2020-02-14
现代园艺(2018年13期)2018-07-11
大众摄影(2018年6期)2018-06-19
武夷学院学报(2017年9期)2017-11-29
广西林业科学(2016年3期)2016-03-16
中国麻业科学(2015年5期)2015-12-28
