
CHK1抑制剂SCH900776抑制胶质瘤细胞U251的增殖及迁移*
2018-03-01 18:25李召君李荣耀费洪荣王凤泽
中国病理生理杂志 2018年2期
李召君, 李荣耀, 王 茹, 费洪荣, 王凤泽△
(泰山医学院 1药学院, 2生命科学学院, 山东 泰安 271016)
神经胶质瘤是神经系统最常见的原发性颅内肿瘤,其死亡率较高,手术治疗后易复发。目前临床上胶质瘤以手术治疗为主,辅以放化疗等综合治疗,但是由于其恶性程度高,且呈浸润性生长,手术难以根除。因此,寻求控制或治愈胶质瘤的有效手段仍是其临床治疗的主要任务。当前,更多的科研工作者致力于探讨胶质瘤发生发展的内在分子机制,以期寻找胶质瘤诊断和治疗的新途径[1]。SCH900776是一种新型的细胞周期检测点激酶1(checkpoint kinase 1,CHK1)抑制剂,对肺癌、肝癌和乳腺癌等多种肿瘤均表现出较好的抑制作用[2-3]。本研究以人胶质瘤细胞U251为实验对象,观察SCH900776对U251细胞增殖和迁移的影响,同时初步探索其分子机制,为SCH900776未来的临床应用提供实验依据。
材 料 和 方 法
1实验材料
SCH900776购自Selleck Chemicals;DMEM培养基和胎牛血清购自 Gibco;MTT和抗β-actin抗体购自Sigma Aldrich;ECL 试剂盒购自Merck Millipore;抗细胞周期蛋白B1(cyclin B1)、细胞分裂周期蛋白2(cell division cycle protein 2,Cdc2)、p38、细胞外信号调节激酶(extracellular signal-regulated kinase,ERK)1/2、p-ERK1/2和Akt抗体购自Santa Cruz;抗p-Cdc2和p-p38抗体购自Cell Signaling Technology;抗p-Akt抗体购自Abcam;抗p27抗体购自BD Biosciences;II 抗均购自北京中杉金桥生物技术有限公司。
2实验方法
2.1细胞株 人神经胶质瘤细胞株U251购自中国科学院上海生命科学研究院细胞资源中心,用含10%胎牛血清的DMEM培养基在37 ℃、5% CO2条件下培养。
2.2MTT法检测细胞活力 取对数生长期的细胞接种于96孔板内,37 ℃培养过夜。次日于各孔内加入含不同浓度(0、0.1、0.25、0.5、1、2.5、5、10、20 μmol/L)SCH900776的培养基各100 μL,于37 ℃继续培养24 h。终止培养前4 h每孔中加入MTT 溶液 20 μL(5 g/L),然后吸去各孔中的溶液,加入150 μL DMSO充分溶解混匀,于490 nm 处检测各孔的吸光度(A)值,分析计算实验结果。
2.3细胞集落形成实验 接种U251细胞于6孔培养板中,然后加入不同浓度(0、0.5、1和5 μmol/L)的SCH900776进行干预12 d。终止培养后用PBS洗涤2次,接着加入无水甲醇固定细胞20 min。PBS洗涤细胞后,加入结晶紫室温染色10 min。最后于倒置显微镜下对细胞集落进行计数。
2.4流式细胞术检测细胞周期 接种U251细胞于6孔培养板中,然后用不同剂量(0、0.5、1和5 μmol/L)的SCH900776处理细胞24 h。胰酶消化并收集细胞,PBS 洗涤细胞2次后,加入预冷的75%乙醇于4 ℃固定过夜。离心去除乙醇,加入RNase A,于37 ℃作用30 min,接着加入碘化丙啶,4 ℃避光处理 20 min。用FACSCalibur 流式细胞仪检测细胞周期。
2.5划痕愈合实验检测U251细胞的迁移能力 接种U251细胞于24孔板中,然后用枪头行“十”字划痕,PBS洗涤细胞2次后,加入不同浓度(0、0.5、1和5 μmol/L)的含药新鲜培养基继续培养细胞,于划痕后的0 h和48 h在相同位置观察并拍照记录划痕宽度。
2.6Western blot实验 U251细胞用不同浓度SCH900776(0.1、5和10 μmol/L)处理24 h,再用含蛋白酶抑制剂的RIPA裂解液冰上裂解 20 min后,4 ℃离心20 min,取上清,进行SDS-PAGE。电泳结束后将凝胶中蛋白质转移至硝酸纤维素膜上,并用5%脱脂奶粉室温封闭 2 h,TBST 洗膜1次,加入相应 I 抗,4 ℃孵育过夜;TBST洗膜3次,加入 II 抗室温孵育2 h,ECL显色试剂盒曝光显影。实验重复3次。
3统计学处理
采用SPSS 18.0 统计软件分析,数据用均数±标准差(mean±SD)表示,多组数据比较采用单因素方差分析,组间两两比较应用SNK-q检验,以P<0.05 为差异有统计学意义。
结 果
1SCH900776对U251细胞增殖的抑制作用
MTT 实验结果表明,U251细胞经1 μmol/L以上浓度的SCH900776处理24 h后,细胞活力显著降低,细胞活力抑制率显著升高(P<0.05),见图1;细胞经SCH900776作用12 d后,细胞集落形成能力明显受到抑制(P<0.05),表明SCH900776能够明显抑制U251细胞的增殖,见图2。
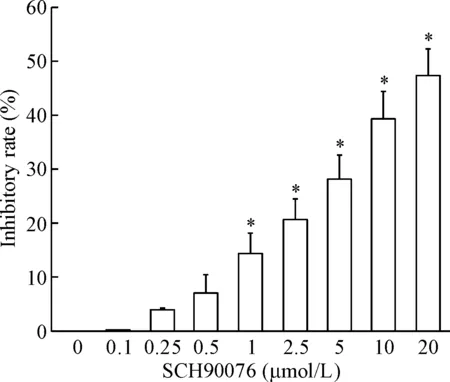
Figure 1. The inhibitory effect of SCH900776 on the viability of U251 cells. Mean±SD.n=3.*P<0.05vs0 μmol/L group.
图1SCH900776抑制U251细胞活力
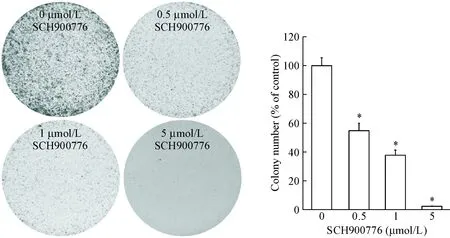
Figure 2. SCH900776 inhibited the colony formation ability of U251 cells. The histogram represents the mean colony numbers. Mean±SD.n=3.*P<0.05vs0 μmol/L group.
图2SCH900776抑制U251细胞集落形成
2SCH900776对U251细胞周期的影响
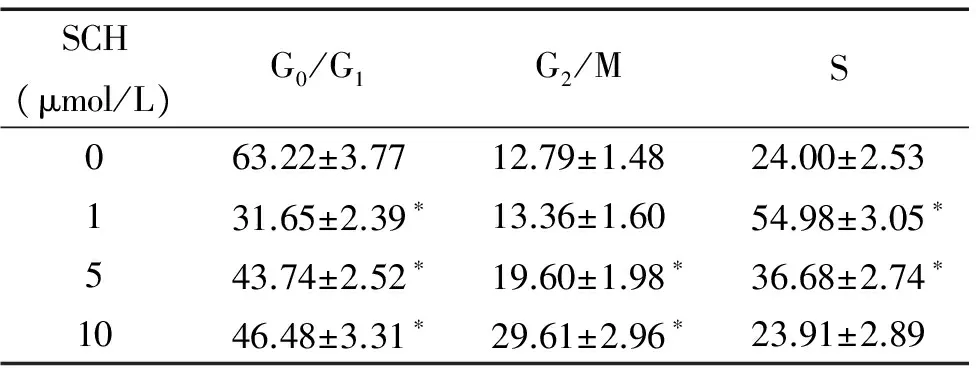
从流式细胞术分析细胞周期结果可以看出,U251细胞经SCH900776作用24 h,在1 μmol/L SCH900776作用的细胞中,S期细胞比例明显增多,而G1期细胞比例则减少(P<0.05);当分别用5 μmol/L和10 μmol/L的SCH900776处理细胞后,G2/M期细胞比例则显著升高(P<0.05)。以上结果表明,较低浓度的SCH900776诱导U251细胞发生S期阻滞,而较高浓度的SCH900776阻滞细胞于G2/M期,见图3、表1。
互动混合式教学借助互联网平台和信息通信技术,将MOOC、微课、翻转课堂等热议的网络教学模式与传统课堂面对面教学的优势相结合,弥补传统教学中存在的不足[3]。现代职教课程可概括为“线上、线下、职场化”。基本做法是将在线教学与面对面课堂教学进行一体化设计,教师提前录制教学微视频等课程资源上传到教学平台,学生课前登录教学平台观看课件、视频,在线测试、讨论或作业。教师根据学生在线学习情况,完善课堂教学设计。课堂教学以学生为主,开展分组讨论、项目实战、任务学习、展示交流、作业及评价等活动,教师主要负责解疑答惑、组织活动。
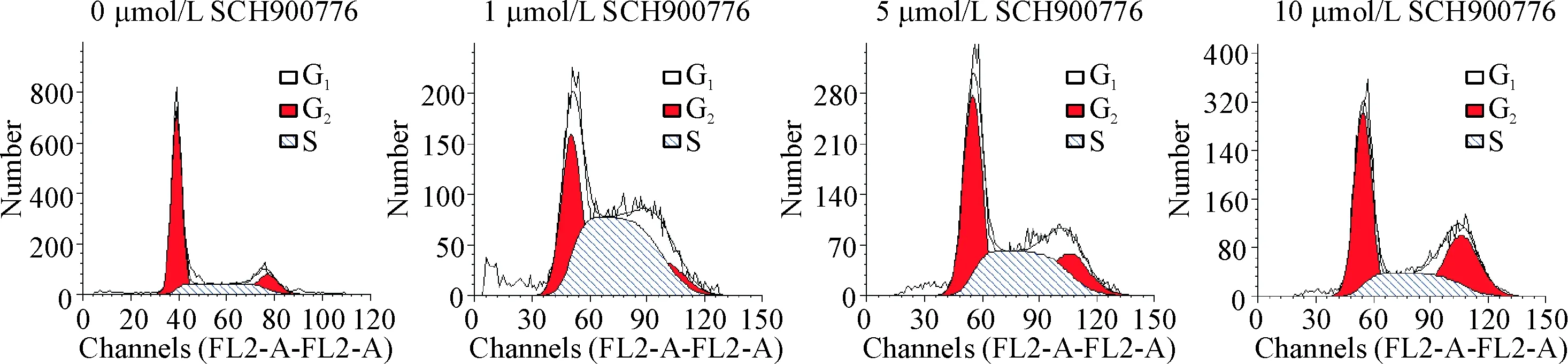
Figure 3. The effect of SCH900776 on the cell cycle distribution of U251 cells.
图3SCH900776对U251细胞周期的影响
表1SCH900776对U251细胞周期的影响
Table 1. Upon exposure to SCH900776 (SCH) for 24 h, the cell cycle distribution of U251 cell was determined by flow cytometry analysis (%. Mean±SD.n=3)
SCH(μmol/L)G0/G1G2/MS063.22±3.7712.79±1.4824.00±2.53131.65±2.39∗13.36±1.6054.98±3.05∗543.74±2.52∗19.60±1.98∗36.68±2.74∗1046.48±3.31∗29.61±2.96∗23.91±2.89
*P<0.05vs0 μmol/L group.
接着,我们应用Western blot实验检测了SCH900776对细胞周期调控蛋白水平的影响。从结果可以看出,5和10 μmol/L SCH900776处理U251细胞24 h后,Cdc2和p-Cdc2的蛋白水平均明显下凋,而cyclin B1的蛋白水平则显著上调(P<0.05),p27的表达未见明显变化,见图4。
3SCH900776抑制U251细胞的迁移
划痕愈合实验结果显示,与0 μmol/L组相比,在SCH900776作用的U251细胞中,细胞迁移能力均明显下降(P<0.05),见图5。
4SCH900776对丝裂原活化蛋白激酶(mitogen-activatedproteinkinase,MAPK)信号通路相关蛋白表达的影响
为进一步探讨SCH900776抑制U251细胞增殖与迁移的分子机制,本研究检测了SCH900776对MAPK信号通路关键因子p38和ERK活性的影响。从结果可以看出,在SCH900776作用的U251细胞中,p38的磷酸化水平明显升高,表明SCH900776能够激活p38 MAPK;ERK的磷酸化水平未见明显变化,见图6。
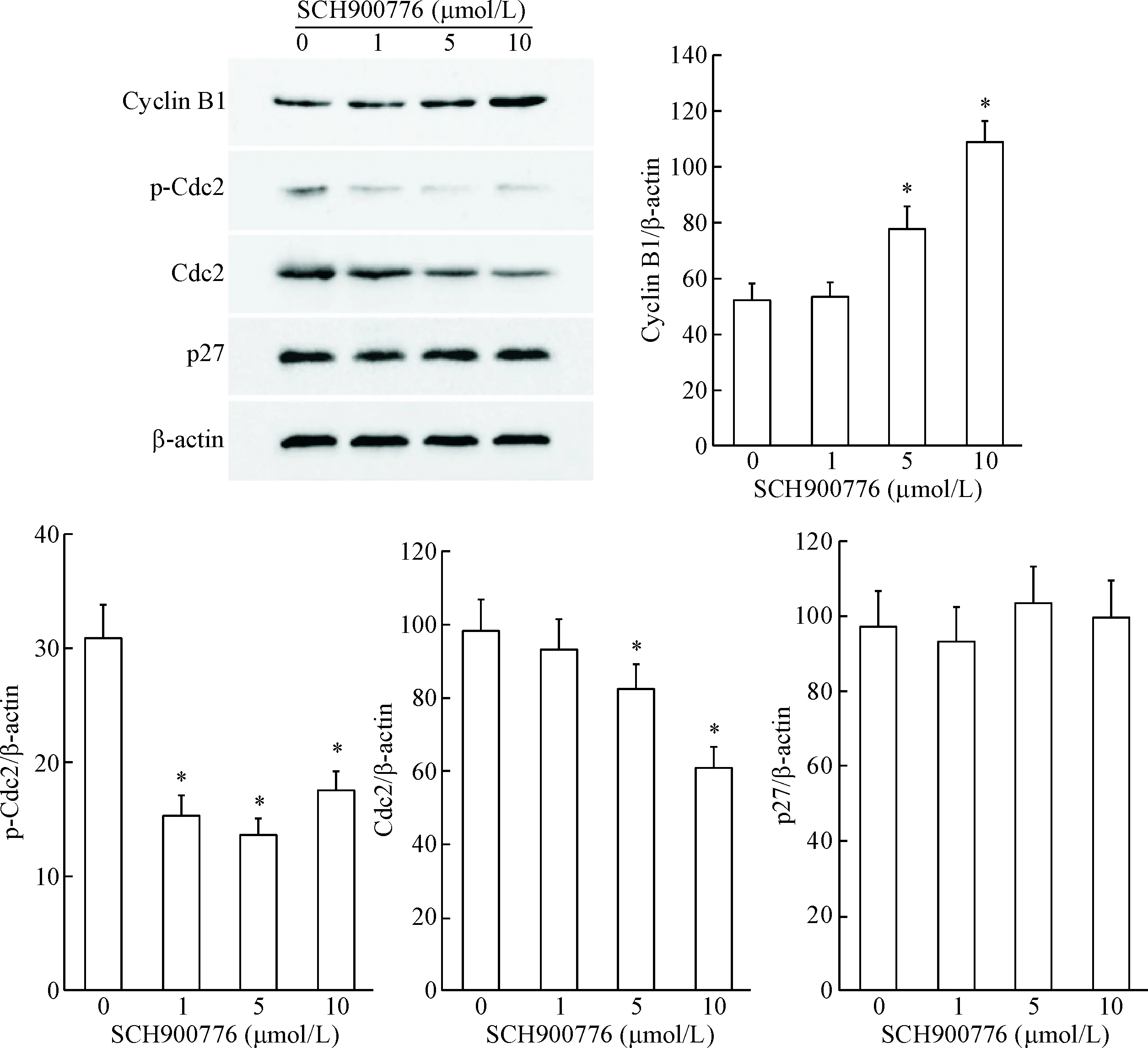
Figure 4. SCH900776 down-regulated the protein levels of p-Cdc2 and Cdc2, amd up-regulated cyclin B1 expression in U251 cells. Mean±SD.n=3.*P<0.05vs0 μmol/L group.
图4SCH900776对U251细胞周期相关蛋白表达水平的影响
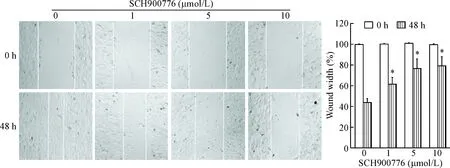
Figure 5. The effect of SCH900776 on the migration ability of U251 cells. Mean±SD.n=3.*P<0.05vs0 μmol/L group.
图5划痕愈合实验检测SCH900776对U251细胞迁移的影响
5SCH900776抑制U251细胞中Akt的磷酸化
最后,本研究检测了SCH900776对U251细胞中Akt活性的影响。Western blot结果表明,5和10 μmol/L SCH900776能够显著抑制Akt的磷酸化水平,见图7。
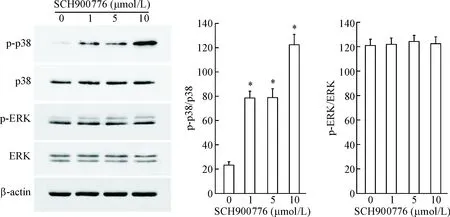
Figure 6. SCH900776 increased the phosphorylation of p38 in U251 cells. Mean±SD.n=3.*P<0.05vs0 μmol/L group.
图6SCH900776上调U251细胞中p38的磷酸化水平

Figure 7. SCH900776 suppressed the phosphorylation of Akt in U251 cells. Mean±SD.n=3.*P<0.05vs0 μmol/L group.
图7SCH900776抑制U251细胞中Akt的磷酸化水平
讨 论
SCH900776(MK-8776)是一种选择性CHK1抑制剂,能够诱导肺癌细胞和白血病细胞等发生DNA损伤,并促进肿瘤细胞对放化疗的敏感性[4-5]。本研究发现,SCH900776能够通过调控细胞周期进而抑制U251细胞的增殖和迁移。SCH900776的抗肿瘤活性可能与其激活p38 MAPK信号通路和抑制Akt磷酸化相关。
细胞周期进程决定着肿瘤细胞的增殖与生长。本实验表明,SCH900776对U251细胞增殖表现出明显的阻抑作用,较低浓度的SCH900776诱导细胞阻滞于S期;而较高浓度的SCH900776作用细胞后,U251细胞发生G2/M期阻滞。同时,细胞周期相关蛋白Cdc2和p-Cdc2的表达量降低,cyclin B1表达水平增加,表明SCH900776对U251细胞周期阻滞行为与其调控上述细胞周期相关蛋白的表达密切相关。
MAPK家族参与调控细胞增殖、迁移及凋亡等生理病理过程[6-7]。p38是细胞感受外届应激信号并引起应答的一个关键分子[8]。当细胞受紫外线或细胞因子等刺激时,非磷酸化的p38与ATP结合并发生磷酸化激活,进而激活相关下游底物,参与调节细胞的增殖和迁移等过程。有文献报道,小分子化合物对胶质瘤细胞增殖和凋亡的调控与p38 MAPK相关[9-10]。本研究发现,SCH900776促进p38的磷酸化,表明SCH900776的抗肿瘤活性可能与其激活p38 MAPK信号通路密切相关。
PI3K/Akt信号通路调控众多与肿瘤发生发展密切相关的生物学过程,如细胞凋亡、增殖及细胞周期调控等[11]。该信号通路活化能够加快细胞周期进程而促进细胞增殖,同时促进肿瘤的侵袭和转移[12]。Akt是PI3K信号途径的一个重要下游靶激酶,具有丝/苏氨酸激酶活性。拮抗Akt活性抑制了肿瘤细胞的增殖与迁移等过程[13]。本研究发现,SCH900776下调Akt的磷酸化水平,且抑制了U251细胞的增殖和迁移,预示着PI3K/Akt信号通路可能与SCH900776抑制U251细胞增殖与迁移过程相关。
总之,本研究发现CHK1抑制剂SCH900776能明显抑制神经胶质瘤U251细胞增殖与迁移,SCH900776的抑癌作用可能与其激活p38 MAPK和抑制Akt的磷酸化相关。SCH900776对p38 MAPK和Akt活性调控的详细分子机理还需进一步深入探讨。本研究对SCH900776未来的临床应用提供了理论指导和实验基础。
[1] Xiong Y, Ji W, Fei Y, et al. Cathepsin L is involved in X-ray-induced invasion and migration of human glioma U251 cells[J]. Cell Signal, 2017, 29:181-191.
[2] Zhou ZR, Yang ZZ, Wang SJ, et al. The Chk1 inhibitor MK-8776 increases the radiosensitivity of human triple-negative breast cancer by inhibiting autophagy[J]. Acta Pharmacol Sin, 2017, 38(4):513-523.
[3] Sakurikar N, Thompson R, Montano R, et al. A subset of cancer cell lines is acutely sensitive to the Chk1 inhibitor MK-8776 as monotherapy due to CDK2 activation in S phase[J]. Oncotarget, 2016, 7(2):1380-1394.
[4] Grabauskiene S, Bergeron EJ, Chen G, et al. CHK1 levels correlate with sensitization to pemetrexed by CHK1 inhibitors in non-small cell lung cancer cells[J]. Lung Cancer, 2013, 82(3):477-484.
[5] Chaudhuri L, Vincelette ND, Koh BD, et al. CHK1 and WEE1 inhibition combine synergistically to enhance therapeutic efficacy in acute myeloid leukemiaexvivo[J]. Haematologica, 2014, 99(4):688-696.
[6] 昌毓穗, 刘季春, 傅华群, 等. Ras-MAPK 通路在食管癌中的研究进展[J]. 中国病理生理杂志, 2013, 29(2): 376-380.
[7] Low HB, Zhang Y. Regulatory roles of MAPK phosphatases in cancer[J]. Immune Netw, 2016, 16(2):85-98.
[8] Cuadrado A, Nebreda AR. Mechanisms and functions of p38 MAPK signaling[J]. Biochem J, 2010, 429(3):403-417.
[9] Baik JS, Mun SW, Kim KS, et al. Apoptotic effects of cordycepin through the extrinsic pathway and p38 MAPK activation in human glioblastoma U87MG cells[J]. J Microbiol Biotechnol, 2016, 26(2):309-314.
[10] Liu QR, Liu JM, Chen Y, et al. Piperlongumine inhibits migration of glioblastoma cells via activation of ROS-dependent p38 and JNK signaling pathways[J]. Oxid Med Cell Longev, 2014, 2014:653732.
[11] Yang SX, Polley E, Lipkowitz S. New insights on PI3K/AKT pathway alterations and clinical outcomes in breast cancer[J]. Cancer Treat Rev, 2016, 45:87-96.
[12] Mayer IA, Arteaga CL. The PI3K/AKT pathway as a target for cancer treatment [J]. Annu Rev Med, 2016, 67:11-28.
[13] 李若彤, 王 丽, 曹 亭, 等. Perifosine通过抑制PI3K/Akt途径调节人胶质瘤U251细胞增殖、凋亡与自噬[J]. 中国病理生理杂志, 2016, 32(4):644-650.
猜你喜欢
中国现代医生(2022年19期)2022-08-25
皮肤性病诊疗学杂志(2021年5期)2021-11-27
皮肤病与性病(2021年3期)2021-07-30
三农资讯半月报(2021年1期)2021-01-27
中华养生保健(2020年4期)2020-11-16
东北大学学报(自然科学版)(2020年5期)2020-05-22
科学技术创新(2020年4期)2020-01-08
满族文学(2018年6期)2018-12-27
学生天地·小学低年级版(2017年12期)2018-04-16
中成药(2017年9期)2017-12-19
