
棉铃红粉病鉴定及气候条件对病菌生长和孢子萌发的影响
2018-02-13 06:32焦瑞莲任毓忠李志强董海涛武刚张莉李国英
新疆农业科学 2018年10期
焦瑞莲,任毓忠,李志强,董海涛,武刚,张莉,李国英,
(1.石河子大学农学院/新疆绿洲农业病虫害治理与植保资源利用自治区高校重点实验室,新疆石河子 832003; 2.石河子开发区石大惠农科技开发有限公司,新疆石河子 832003; 3.新疆生产建设兵团第一师农科所,新疆阿拉尔 843300)
0 引 言
【研究意义】棉铃病害是棉花上的一类重要病害,棉花红粉病是棉铃病害中分布较广的铃病之一,世界各棉区都有发生,在我国各棉区也有分布。以往在新疆棉区发生很轻,但近年来发生有加重之势,并常和其它铃病混合发生。研究红粉病的病原及其气候条件对病菌生长、产孢量和分生孢子萌发的影响,对该病的防治具有重要的作用。【前人研究进展】棉铃红粉病是由粉红聚端孢菌Trichotheciumroseum引起,该菌为腐生或弱寄生菌,可引起果蔬红粉病,如:苹果、厚皮甜瓜、葡萄、枣果、穗醋栗、西红柿、菜豆等;还可引起黄麻、棉铃等至少15种植物红粉病害。Richard等[1]研究发现一定剂量的粉红聚端孢的毒性产物可致老鼠、家兔和幼猪死亡,同时,若将毒性产物注射到动物真皮时,可造成动物真皮坏死。该菌于1809年在德国柏林首次被Link发现,1902年Eustace在纽约苹果上发现红粉病,随后在世界各地不断出现该病害报道。如Shamsi等[2](2008)在孟加拉发现黄麻红粉病害;Yun等[3](2013)在韩国发现番茄红粉病;Hamid等[4](2014)在巴基斯坦发现T.roseum可导致番茄、橙子和苹果果腐等。在国内,戚佩坤(1966年)等报道了黄瓜红粉病;章战华[5](2007)对棉花、葡萄、苹果、甜瓜、香梨、油菜、茄子、草莓上的红粉病菌进行了研究;王勇等[6](2008)报道了番茄上红粉病;潘月敏等[7,8](2005)和黎鸿慧等(2011)报道了棉花红粉病,严重发生时可使棉花减产10%。冯中红等[9](2015)报道了菜豆红粉病;杨济徽等(2016)[10]报道了龙眼红粉病;均由T.roseum引起。但对气候条件对菌丝生长、产孢和孢子发芽的影响缺乏研究。另外,从近期报道的资料来看,对其学名多用Trichotheciumroseum,但有的书中用Cephalotheciumroseum,有的甚至把它们作为为害棉铃的两种菌[11]。【本研究切入点】根据前人研究的结果和新疆对其缺乏研究的实际情况,有必要对新疆棉铃红粉病进行较为系统的鉴定。研究结合气象因子对病菌生长、产孢和孢子萌发的影响。【拟解决的关键问题】研究新疆棉花红粉病的病原,明确气象因子与病菌生长、产孢和孢子萌发的关系。分析新疆棉铃红粉病菌的病原及其生物学特性,为制定有效的防治提供理论依据。
1 材料与方法
1.1 材 料
2017年分别于8和9月,对北疆石河子棉区150团4连和9连、147团19连、142团23连、133团20连,以及南疆阿克苏棉区12团、阿瓦提县、塔河种业等地一些棉田的棉铃红粉病害进行调查。共采集典型症状的病样19份,供分离鉴定,对田间症状和接种后的症状进行观察、描述和拍照。
1.2 方 法
1.2.1 病菌的分离、纯化及代表性菌株的选择
在分离前将所要分离的病铃流水冲12 h,取下,晾干表面水分。用解剖刀切取5 mm×5 mm的病斑,分别在70%酒精中15s和0.1%升汞中停留2~5s、无菌水冲洗3次、置于吸水纸上将水分吸干后,在无菌条件下移植于PDA平板上,将平板置于28℃培养箱中。1~2 d待长出菌落,挑取边缘菌丝于新的PDA培养基上,继续培养。共分离出96个菌株,待培养3~4 d左右根据形态特征选出16个代表性菌株进行单孢分离纯化。表1
1.2.2 病原鉴定1.2.2.1 形态学鉴定
将单孢分离的16个代表性菌株在PDA培养基上培养7 d后,观察菌落颜色和形态,并镜检其分生孢子形态及颜色;测量孢子大小。采用玻片培养法,25℃下,培养6 d,观察分生孢子梗的形态及分生孢子在分孢梗顶端的着生状态。并拍照,作为病原鉴定的主要形态学依据。用测微尺测量50个孢子和孢子梗的大小。
表1 菌株编号及来源
Table 1 The strain ideutifiers and sources origin
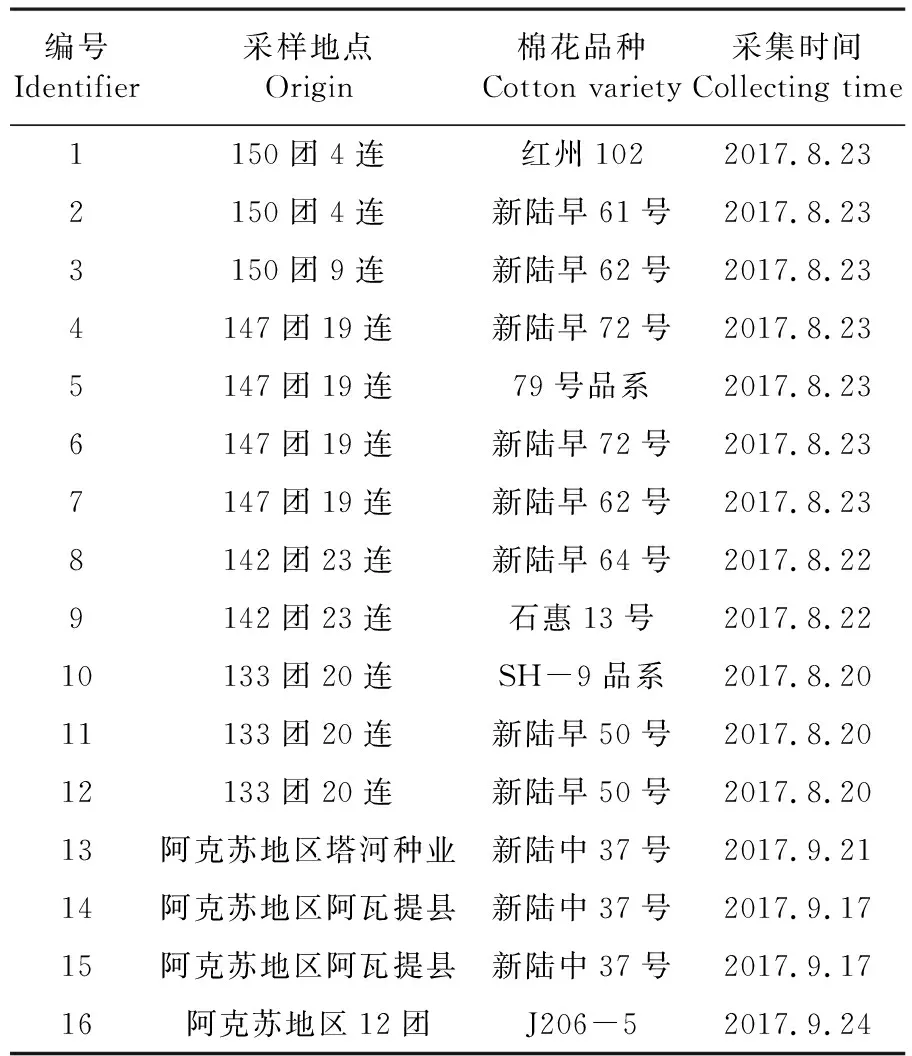
编号Identifier采样地点Origin棉花品种Cotton variety采集时间Collecting time1150团4连红州1022017.8.232150团4连新陆早61号2017.8.233150团9连新陆早62号2017.8.234147团19连新陆早72号2017.8.235147团19连79号品系2017.8.236147团19连新陆早72号2017.8.237147团19连新陆早62号2017.8.238142团23连新陆早64号2017.8.229142团23连石惠13号2017.8.2210133团20连SH-9品系2017.8.2011133团20连新陆早50号2017.8.2012133团20连新陆早50号2017.8.2013阿克苏地区塔河种业新陆中37号2017.9.2114阿克苏地区阿瓦提县新陆中37号2017.9.1715阿克苏地区阿瓦提县新陆中37号2017.9.1716阿克苏地区12团J206-52017.9.24
1.2.2.2 分子生物学
将供试的16个单孢菌株在PDA平板上培养7 d后,用刀片刮取菌丝体,用Bio Flux试剂盒提取供试菌株的基因组DNA。将提取的DNA置于-20℃冰箱中保存,备用。以提取的菌株基因组DNA为模板,用真核生物rDNA-ITS区通用引物ITS1(5’-TCCGTAGGTGAACCTGCGG-3’)和ITS4(5’-TCCTCCGCTTATTGATATGC-3’)进行PCR扩增,用1%的琼脂糖凝胶检测扩增结果,后用回收试剂盒回收纯化扩增产物(DNA回收试剂盒)。回收产物经连接转化后,筛选出具有代表性的阳性克隆,送上海生工(Sangon Biotech)公司测序。将所测得的序列利用Gen Bank BLAST进行同源性比对,并在NCBI核酸数据库中下载其他相关病菌的ITS区序列,用DNAMAN 8.0软件进行序列的比对并构建系统发育树,分析菌株间的亲缘关系。
1.2.2.3 致病性鉴定
分别采用菌丝块接种和喷雾接种两种方法进行。
菌丝块接种:在棉田随机选取健康、大小均匀的棉铃,用70%酒精擦拭后再用无菌水冲洗3次,供接种用。分有伤和无伤接种于准备好的棉铃上,每菌株接种3个棉铃,并取不接种棉铃作为对照,将其置于保湿缸中室温下观察,并记录其发病情况和症状表现,7 d后结束并拍照。
孢子悬浮液接种:按常规方法配置孢子悬浮液(1×106cfu/mL),装入小型喷雾器内,分别在室内和田间进行喷雾接种,室内接种条件同菌丝块接种,田间接种为自然条件下田间棉株上的棉铃,均以不接种棉铃为对照,观察并记录其发病情况及症状,7 d后结束并拍照。同时对发病株进行再分离,观察再分离菌株与接种菌株的异同。
1.2.3 气候条件对供试菌株生长及产孢量影响1.2.3.1 温度
按1.2.2.3的方法制取菌饼,将其移植到PDA培养基平板中央,分别在5、10、15、20、 25、30、35和40℃ 8个梯度的培养箱中恒温培养,3次重复,并分别于24、48、72、96、120、144和168 h测量菌落直径,共测量7 d。在第7 d用打孔器打取7 mm的菌饼,并按每毫升无菌水3个菌饼的量置于2 mL离心管中,充分震荡,洗下分生孢子。用血球计数板计量分生孢子量,观察温度对菌落生长及产孢量的影响。
1.2.3.2 光照
将上述供试菌株菌饼分别移植于PDA培养基上,设置完全光照、12 h光暗交替和完全黑暗3个处理,在 25℃条件下进行培养,每组3次重复,并分别于24、48、72、96、120、144和168 h测量菌落直径,分生孢子量的计算方法同1.2.3.1。
1.2.3.3 温度对孢子萌发的影响
用挑针挑取培养7 d的菌落边缘分生孢子于凹形载玻片上,将载玻片置于相对湿度为100%的培养盒中,并将培养盒分别置于5、10、15、20、25、30、35和40℃的人工气候箱中黑暗培养,每种温度均三次重复。每12 h随机观察1次各供试温度中100个分生孢子的萌发情况,48 h结束,共观察4次。用SPSS 8.0分析温度对孢子萌发的显著性。
1.2.3.4 光照对孢子萌发的影响
用挑针挑取培养7 d的菌落边缘分生孢子于凹形载玻片上,将载玻片置于相对湿度为100%的培养盒中,之后将培养盒分别放于完全光照、完全黑暗和光暗交替的25℃人工气候箱中。每12 h随机观察100个分生孢子萌发情况,48 h结束,共观察4次。用SPSS 8.0分析光照对孢子萌发的显著性。
1.2.3.5 湿度对孢子萌发的影响
用挑针挑取培养7 d的菌落边缘分生孢子于凹形载玻片上,并将载玻片置于40%、60%、80%、100%四个相对湿度梯度的培养盒中;同时挑取分生孢子于无菌水中,盖上盖玻片置于培养盒中作为对照,重复3次。培养盒均置于25℃人工气候箱中黑暗培养,每12 h随机观察100个分生孢子萌发情况,48 h结束,共观察4次。
1.3 数据处理
用SPSS 8.0分析湿度对孢子萌发的显著性。
2 结果与分析
2.1 棉花红粉病发病情况的调查和致病性测定
研究表明,棉铃红粉病危害的棉铃一般表现为,初期在棉铃铃壳或棉铃缝隙处形成褐色小点,后逐渐扩大成圆形或近圆形,黑褐色病斑,湿度较大时,病斑处初生白色(菌丝体),后呈粉红色、厚且紧密的霉层。严重时霉层可局部或全部布满铃面。使棉铃不能正常吐絮、甚至形成僵铃、僵瓣,且僵瓣上也会分布有粉红色霉层、使纤维变为粉红色或浅褐色(图1-A:a、b、c)。
致病性测定表明,菌丝块接种后无论有伤接种还是无伤接种都可使棉铃发病,但有伤接种比无伤接种发病早,病情也较重。接种48 h后在棉铃接种处出现褐色小点,后褐色区域逐渐扩大呈圆形或近圆形、下陷的黑褐色病斑,湿度大时在病斑中心长出白色粉状物,随着病斑的扩大,霉层扩增,颜色加深,呈粉白色或粉红色(图1-B:b、c)。用孢子悬浮液进行室内离体棉铃喷雾接种16 h后,棉铃表面出现水浸状褐色小点,后不断扩大成黑褐色斑点;若田间接种,发病较迟,第4 d出现水浸状褐色小点;而室内、田间没接种对照棉铃均不发病。将接种后的病铃进行再分离, 得到与接种菌株形态相同的病原物。图1
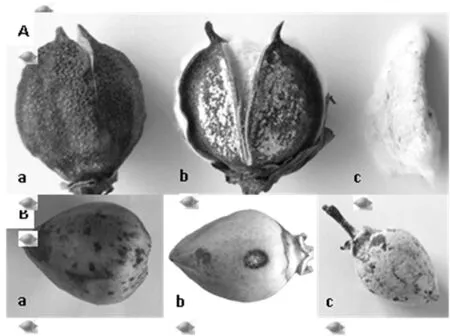
注:A田间症状:a、b田间病铃症状;c棉纤维感染症状;B人工接种症状:a喷雾接种症状;b菌丝块接种症状; c后期症状
Note: A field symptoms:a, b: cotton boll symptom in field; c: symptoms of cotton fiber infection; B artificial inoculation symptom:a: spray inoculation symptom; b: symptoms of hyphae block Inoculation; c: the late symptoms
图1 棉铃红粉病田间与接种症状
Fig.1 Field and inoculation symptoms of cotton boll red powder disease
2.2 棉花红粉病菌的形态学鉴定
将单孢分离纯化的16个菌株移植到PDA培养基上,置于28℃恒温培养箱中培养。第1 d长出浅白色气生菌丝;第2 d出现少量分生孢子,菌落边缘呈浅白色;第3 d,由于菌落上产生大量分生孢子,菌落开始呈现粉红色;后菌落边缘呈不规则齿状或波状,有的具明显的同心轮纹,菌落正面为粉红色,背面为粉黄色,边缘浅白色(图2-A、B)。由小室培养进行镜检,分生孢子梗直立不分枝,具几个隔膜,长50~200 μm,平均132.75 μm。分生孢子单生或聚生于分生孢子梗顶端,无色,聚集时呈粉色,梨形或卵圆形,双孢,中间隔膜处稍溢缩,孢基有一偏向乳头状突起,大小为(12.5~22.5)μm×(5~12.5)μm,平均大小为16.7μm×8.8 μm(图2-C、D)。孢子萌发时芽管可从分生孢子基部、顶部、侧面等部位萌发,可一端或两端同时萌发,芽管分支或不分支(图2-E)。经形态学鉴定该病病原为粉红聚端孢菌(Trichotheciumroseum)。图2
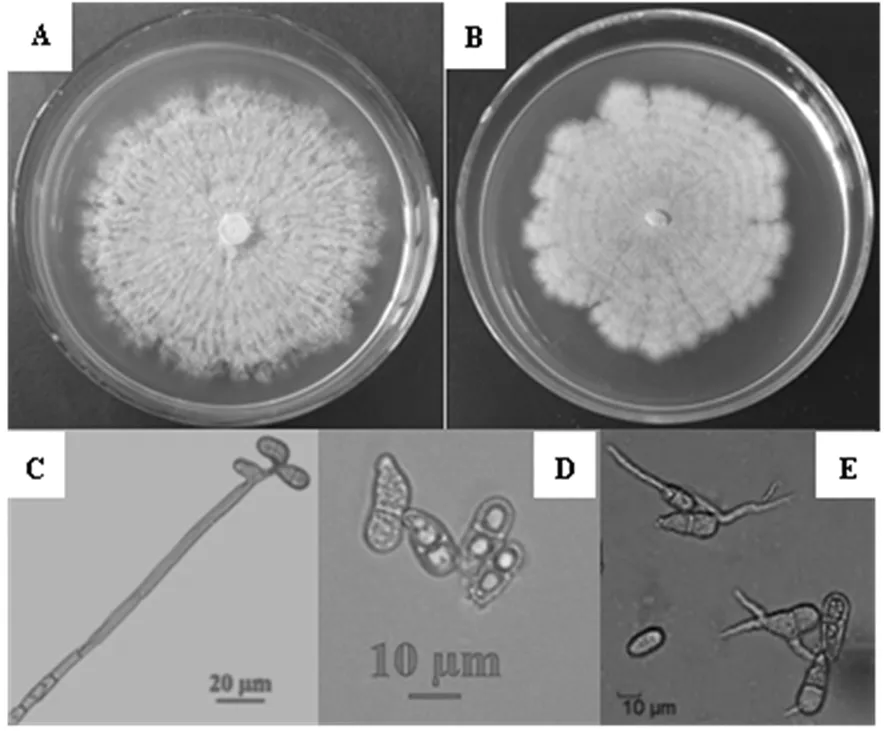
注:A、B菌落形态;C分生孢子梗;D分生孢子;E 孢子萌发
Note : A/B Colony morphology;C Conidiophores;D Conidia;E Spore germination
图2 棉铃病原菌形态特征
Fig.2 Morphological characteristics of cotton boll pathogenic fungus
2.3 分子生物学检测
提取16个供试菌株DNA,利用rDNA-ITS区引物ITS1/ITS4进行PCR扩增,扩增片段为611~613 bp,对扩增产物进行琼脂糖凝胶电泳检测并回收DNA,将回收的DNA做连接、转化,筛选阳性单克隆菌斑进行培养,后送上海生工生物公司测序。对所测得的序列在NCBI Blast 中进行同源比对。比对结果表明,1、3~10、13~16菌株与T.roseum(KP317992.1) 相似性为100%;2、11与T.roseum(JQ898156.1)的相似性为100%,而12与T.roseum(JQ898156.1) 的相似性达到98%。图3
通过使用DNAMAN构建系统发育树发现,其中15个供试菌株与T.roseum都归在同一分支,只有12不在T.roseum的分支上,但与聚端孢属(Trichothecium.spp)的其他种如T.ovalisporum)及F.solani不在同一分支上。在对12号菌株的菌落形态、分生孢子、分生孢子梗分析中发现其与其他15个菌株基本相似;且在NCBI Blast同源比对中与T.roseum(JQ898156.1)的相似性为98%。结合各菌株的形态学特征和NCBI Blast同源比对结果,初步认为供试菌株都为T.roseum。图4
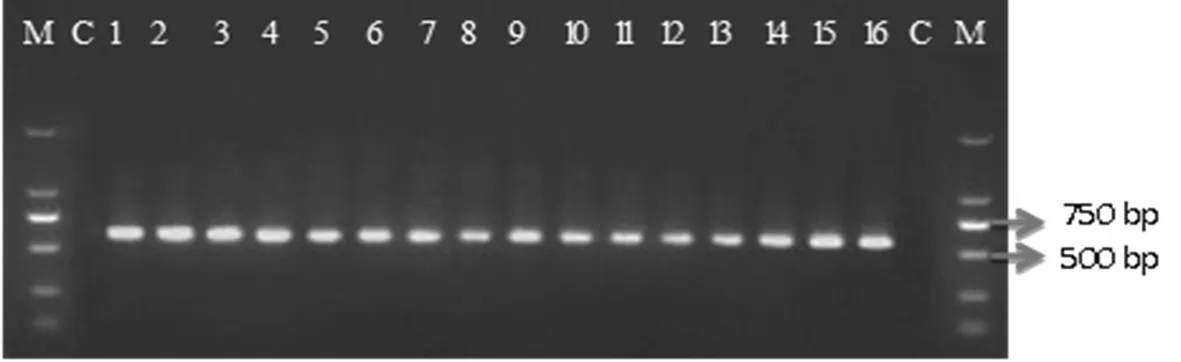
注:M 标准DL 2000 bp;C阴性对照;H1-H16供测菌株
Note: M Molecular size marker DL,2000 bp,C Negative control;H1-H16 Test strain
图3 供测菌株rDNA-ITS区PCR扩增
Fig.3 PCR amplification in the rDNA- ITS region of the test strain
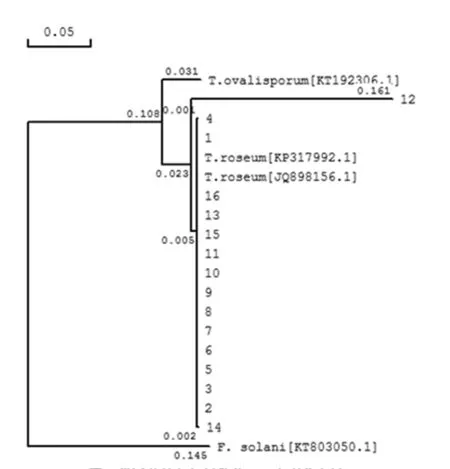
图4 供试菌株与相似种的ITS序列发育树
Fig.4 ITS sequence Phylogenetic tree of the pathogen and similar species
2.4 气候条件对病菌生长、产孢量和孢子萌发的影响
2.4.1温度条件对病菌生长及产孢量的影响
研究表明,温度对T.roseum的菌落生长有显著的影响,该菌在5和40℃不生长,10和35℃生长极其缓慢,其生长的最适温度是25℃左右。10和35℃菌落均小于2.5 cm。对生长168 h菌落上的分生孢子进行产孢量检测,以25℃条件下生长的菌落的分生孢子量最多,达到了8.65×106cfu/mL,其次是在20℃时的分生孢子量,而15和30℃的分生孢子量明显减少,分别为1.75×106cfu/mL和1.9×106cfu/mL;10℃以下和35℃以上不产孢或产孢极少。表2,图5
表2 不同温度条件下棉铃红粉病菌菌落生长及产孢量变化
Table 2 Effects of different temperatures on colony growth and sporulation capacity of cotton boll pathogenic fungus
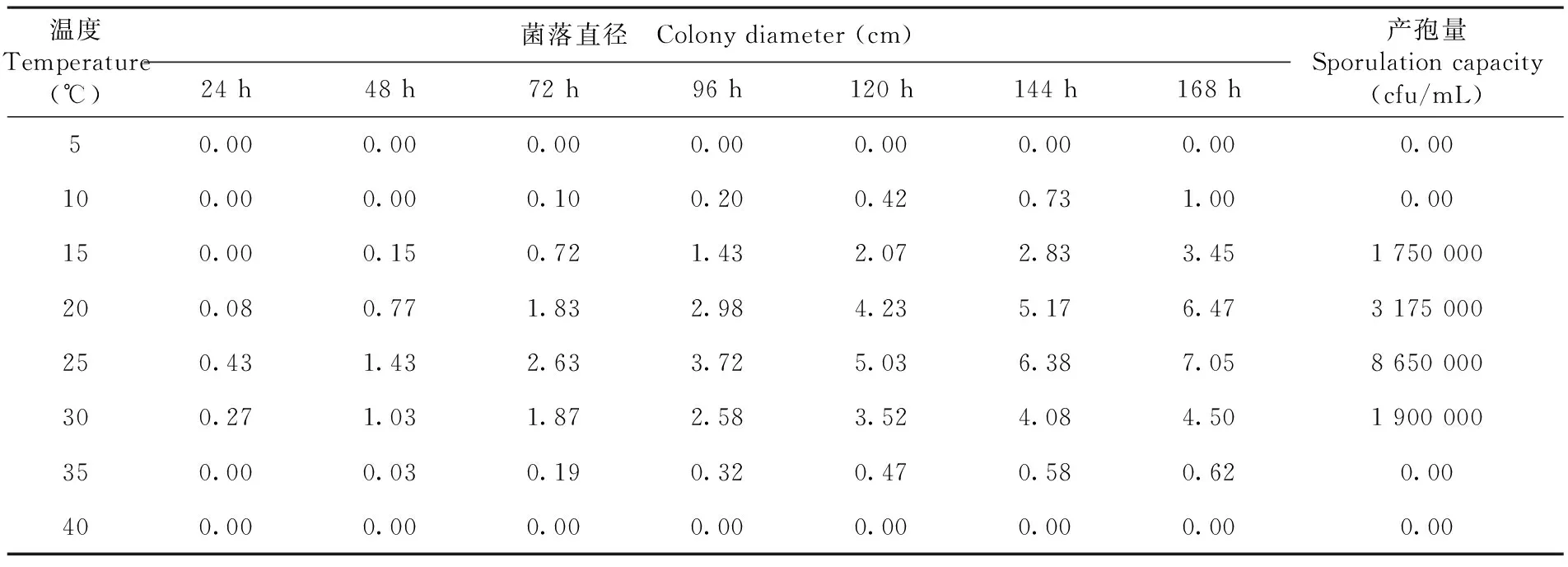
温度Temperature(℃)菌落直径 Colony diameter (cm)24 h48 h72 h96 h120 h144 h168 h产孢量Sporulation capacity(cfu/mL)50.000.000.000.000.000.000.000.00100.000.000.100.200.420.731.000.00150.000.150.721.432.072.833.451 750 000200.080.771.832.984.235.176.473 175 000250.431.432.633.725.036.387.058 650 000300.271.031.872.583.524.084.501 900 000350.000.030.190.320.470.580.620.00400.000.000.000.000.000.000.000.00

图5 不同温度下棉铃红粉病菌的菌落形态
Fig.5 Colony morphology of cotton boll pathogenic fungus at different temperatures
2.4.2 光照条件对菌落生长及产孢量的影响
研究表明,在三种光照处理条件下,对T.roseum的生长没有明显的差异,但是对产孢量的影响很大,其中以完全光照产孢量最少,为1.85×106cfu/mL;完全黑暗产孢量最大,为8.93×106cfu/mL;光暗交替处于两者之间,其产孢量为5.63×106cfu/mL。光线对产孢有明显的抑制作用。不同光照条件对菌落的形态特征也有明显影响,完全光照和完全黑暗所长出的菌落不产生轮纹;而光暗交替条件下所长出的菌落,由于光暗条件产孢量的明显不同,则产生明显的轮纹。表3,图6
表3 不同光照条件下棉铃红粉病菌菌落生长及产孢量变化
Table 3 Effects of different illumination on colony growth and sporulation capacity of cotton boll pathogenic fungus
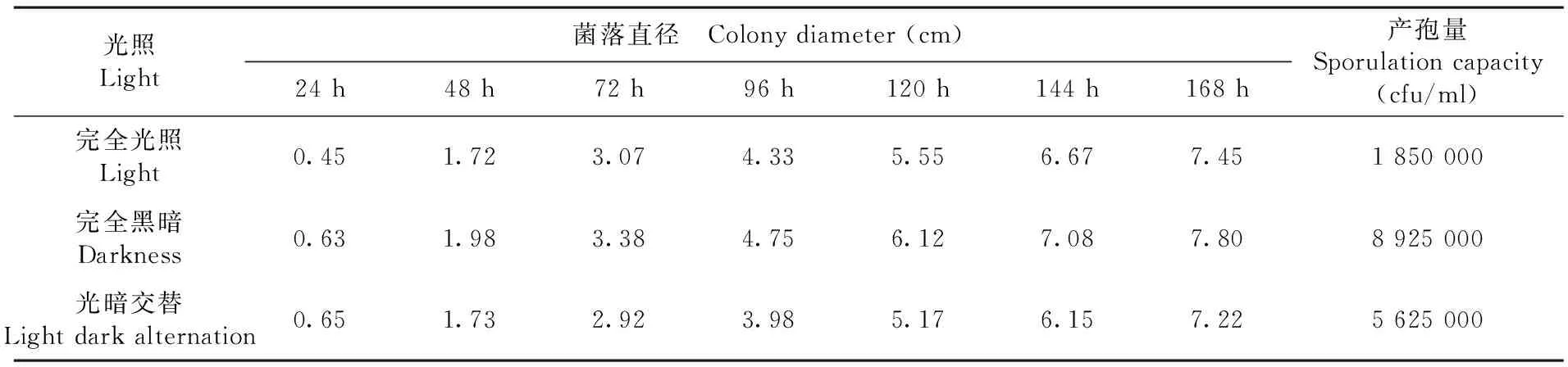
光照Light菌落直径 Colony diameter (cm)24 h48 h72 h96 h120 h144 h168 h产孢量Sporulation capacity(cfu/ml)完全光照Light0.451.723.074.335.556.677.451 850 000完全黑暗Darkness0.631.983.384.756.127.087.808 925 000光暗交替Light dark alternation0.651.732.923.985.176.157.225 625 000
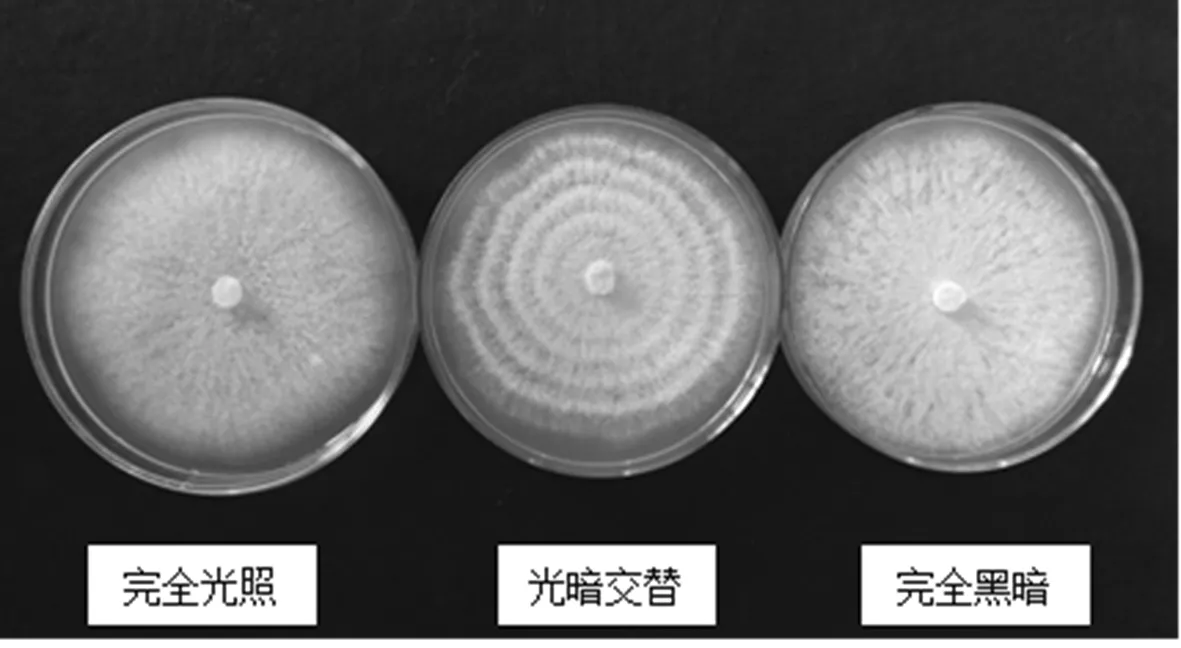
图6 不同光照条件下菌落形态
Fig.6 Colony morphology under different light conditions
2.4.3 温度条件对孢子萌发的影响
不同温度对T.roseum的分生孢子萌发具有显著影响。试验表明,小于5℃和大于40℃时该菌的分生孢子不能萌发;在10~35℃孢子都可萌发,其萌发的最适温度为20~25℃,48 h时其萌发率为94%~97.67%,15℃以下和35℃以上萌发率明显下降。表4
表4 不同温度条件下棉铃红粉病菌孢子萌发变化
Table 4 Effect of temperature on spore germination of cotton boll pathogenic fungus
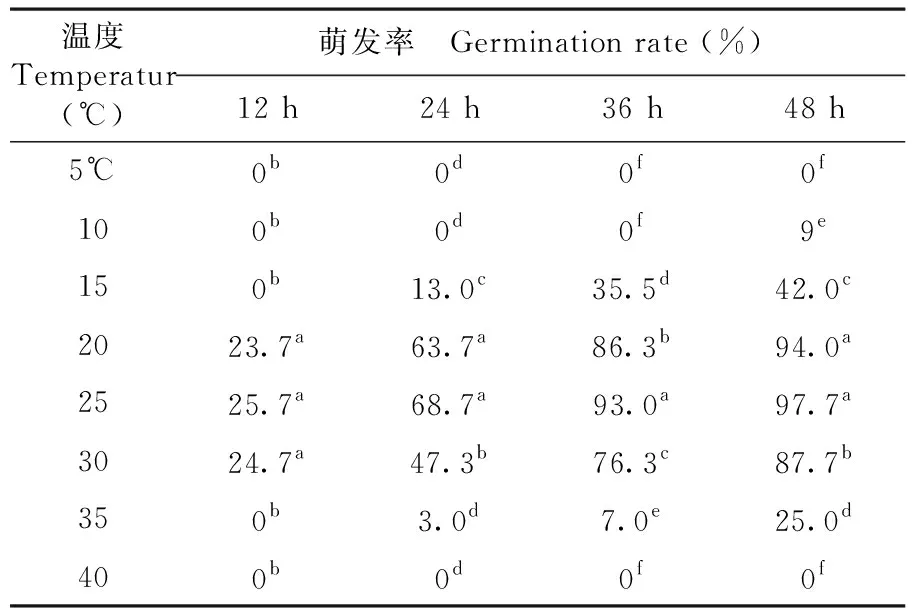
温度Temperatur(℃)萌发率 Germination rate (%)12 h24 h36 h48 h5℃0b0d0f0f100b0d0f9e150b13.0c35.5d42.0c2023.7a63.7a86.3b94.0a2525.7a68.7a93.0a97.7a3024.7a47.3b76.3c87.7b350b3.0d7.0e25.0d400b0d0f0f
注:数据后小写英文字母为0.05水平显著性差异
Note: little letters after date indicate at 0.05 levels
2.4.4 光照条件对孢子萌发的影响
研究证明,在供试三种条件下T.roseum的孢子均可萌发,且萌发率都较高。在光暗交替和完全黑暗条件下,孢子萌发率均可达到93.33%以上,完全光照条件下孢子萌发率低于其他两种条件下的萌发率,为85%。完全光照条件对棉铃红粉病菌T.roseum的分生孢子萌发具有一定的抑制作用,而完全黑暗和光暗交替则有利T.roseum的孢子萌发。表5
表5 不同光照条件下棉铃红粉病菌孢子萌发变化
Table 5 Effect of light on spore germination of cotton boll pathogenic fungus
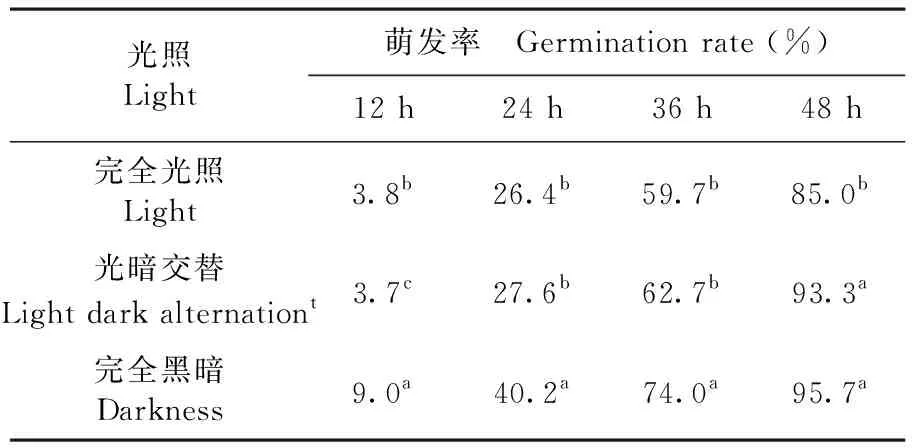
光照Light萌发率 Germination rate (%)12 h24 h36 h48 h完全光照Light3.8b26.4b59.7b85.0b光暗交替Light dark alternationt3.7c27.6b62.7b93.3a完全黑暗Darkness9.0a40.2a74.0a95.7a
注:数据后小写英文字母为0.05水平显著性差异
Note: letters after date indicate at 0.05 levels
2.4.5 湿度条件对孢子萌发的影响
研究表明,在供试的四个相对湿度和水中,48 h后,在100%相对湿度下孢子萌发率为100%;相对湿度在80%时孢子萌发率为80.33%;相对湿度为60%时的萌发率为51.5%;相对湿度为40%时孢子萌发率最低为28.50%;而水中的孢子萌发率为58.7%,与相对湿度60%条件下接近。在40%~100%的相对湿度条件下,湿度越大,孢子的萌发率越高。表6
表6 不同湿度条件下棉铃红粉病菌孢子萌发变化
Table 6 Effect of humidity on spore germination of cotton boll pathogenic fungus
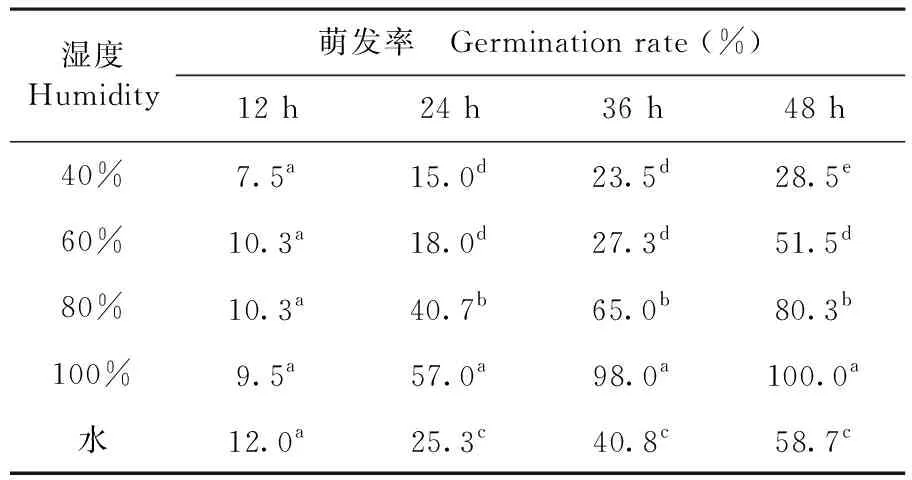
湿度Humidity萌发率 Germination rate (%)12 h24 h36 h48 h40%7.5a15.0d23.5d28.5e60%10.3a18.0d27.3d51.5d80%10.3a40.7b65.0b80.3b100%9.5a57.0a98.0a100.0a水12.0a25.3c40.8c58.7c
注:数据后小写英文字母为0.05水平显著性差异
Note: little letters after date indicate at 0.05 levels
3 讨 论
经鉴定,新疆石河子棉区和阿克苏棉区的棉铃红粉病是由粉红聚端孢菌(T.roseum)所致,这一结果与潘月敏(2005)等[7]及黎鸿慧等(2011)的报道相同[8]。经分子生物学鉴定,在16个代表性菌株中,其中13个菌株与T.roseum(KP317992.1)相似性为100%;2个菌株与T.roseum(JQ898156.1)的相似性为100%,只有12号菌株与T.roseum(JQ898156.1) 的相似性为98%,初步认为在钟内可能存在生物多样性。
国内关于棉铃红粉病的研究较少,对其学名应用也比较混乱。如当前有的使用Trichotheciumroseum, 但也有的用Cephalotheciumroseum,有的甚至把它们作为为害棉铃的两种菌[11]。张中义等[12]在《植物病原真菌学》中就指出Trichotheciumroseum和Cephalotheciumroseum是同物异名。陆家云[13]在所著植物病原真菌学一书中再次强调Trichotheciumlink=CephalotheciumCorda;Trichotheciumroseum=Cephalotheciumroseum,故不能将其作为两种不同的菌。Cephalotheciumroseum只是Trichotheciumroseum的同物异名,目前国内大部分研究单位都使用Trichotheciumroseum,以此为好。
研究查明,该菌在10~35℃条件下均可生长,但10和35℃时生长极其缓慢,25℃左右为其生长的最适温度,5和40℃不能正常生长。对生长7 d的菌落进行产孢量检测,以生长在25℃条件下的分生孢子量最多,达到了8.65×106cfu/mL;其次是在20℃时的分生孢子量;15和30℃的分生孢子量明显减少;10℃以下和35℃以上不产孢或产孢极少。该结果与Yun等[3]和王勇等[7]研究结果基本一致,不同点为试验中5℃菌落不能生长,而王勇等研究中5℃条件下148 h时开始生长。
三种光照条件对T.roseum的菌落生长没有明显的差异,但是对产孢量的影响很大。其中完全光照条件下产孢量最少,完全黑暗产孢量最大,光暗交替处于两者之间,说明光线对产孢有明显的抑制作用;此外,不同光照条件对菌落的形态特征也有一定的影响:完全光照和完全黑暗所长出的菌落不产生轮纹;而光暗交替条件下所长出的菌落,由于光暗条件产孢量的明显不同,则产生明显的轮纹。
分生孢子萌发是病菌侵染的前提,研究气候条件对病菌孢子萌发的作用对了解该病的发生规律和防治都有重要作用。研究表明,该菌的分生孢子萌发最适温度为20~25℃,15℃以下或35℃以上孢子萌发率明显下降,萌发时间推迟,而当温度小于5℃或大于40℃时,孢子不能萌发,这一结论与殷辉等[14]在枣果贮藏期红粉病菌鉴定及环境因子对其分生孢子萌发的影响结论相似。关于光照对孢子萌发的影响国内报道很少,试验查明,该菌分生孢子在供试不同光照条件下都可萌发,且48 h萌发率达到了85%以上,但其趋势为:完全黑暗>光暗交替>完全光照,表明光线对孢子萌发有一定的抑制作用。关于相对湿度对孢子萌发的影响,试验的结果是:在40%~100%的相对湿度条件下,湿度越大,孢子的萌发率越高,当湿度大于80%时,最适该菌分生孢子萌发,但在水中的萌发率又明显下降。
由于棉铃红粉病菌产孢量很大,发芽率较高,潜育期较短,对湿度条件要求较高,故秋季一旦遇到持续降雨天气,易造成该病的发生。
4 结 论
经形态学鉴定、致病性测定和分子生物学分析查明,引起棉铃红粉病的病原菌是粉红聚端孢T.roseum。通过气候条件对菌落生长、产孢量和对孢子萌发的影响查明,棉铃红粉菌在10~35℃条件下均可生长,25℃左右为其生长的最适温度;在黑暗潮湿的条件下,25℃左右的温度,适于棉铃红粉菌产孢;该菌的分生孢子在10~35℃、40%~100%的相对湿度和有光和无光的条件下均可萌发,萌发的最适条件是:温度为20~25℃、相对湿度80%以上。在此条件下,相对黑暗的环境更有益于孢子的萌发。
猜你喜欢
红领巾·萌芽(2022年9期)2022-11-24
幼儿画刊(2022年10期)2022-10-14
散文诗(2022年14期)2022-08-08
红领巾·萌芽(2022年6期)2022-06-27
河南科技学院学报(自然科学版)(2021年6期)2021-11-15
疯狂英语·新策略(2021年9期)2021-11-02
棉花学报(2021年1期)2021-02-05
红领巾·萌芽(2019年8期)2019-08-27
热带作物学报(2019年4期)2019-06-11
中国棉花(2019年3期)2019-03-27
