
精子相关抗原9 B细胞及T细胞表位富集区预测
2018-02-07 08:36毛晨晨叶思思张特明韩政薛向阳沈贤
温州医科大学学报 2018年1期
毛晨晨,叶思思,张特明,韩政,薛向阳,沈贤,3
(1.温州医科大学附属第一医院 胃肠外科,浙江 温州 325015;2.温州医科大学 基础医学院,浙江温州 325035;3.温州医科大学附属第二医院 胃肠外科,浙江 温州 325027)
肿瘤/睾丸抗原(cancer/testis antigen,CTA)主要表达在人类生殖系统组织和恶性肿瘤细胞表面,在正常组织中不表达或者很少表达。这其中,精子相关抗原9(sperm associated antigen 9,SPAG-9)是最常见的一种CTA,且可通过调控MAPK信号通路导致肿瘤的发生[1-2]。研究表明,SPAG-9在肾癌、乳腺癌、前列腺癌、肝细胞癌和大肠癌细胞上均存在过表达的现象[3-7],且在肾细胞癌、甲状腺癌和结肠癌等细胞株表面可以检测到SPAG-9,表明SPAG-9可能为分布于肿瘤细胞表面的膜蛋白[8-9]。另外,在宫颈癌、卵巢癌、结肠癌患者体内均可检测到针对SPAG-9的抗体[3-5]。由此,SPAG-9可能成为肿瘤免疫治疗研究的新靶点[9]。
抗原表位,即抗原分子中决定抗原特异性的特殊结构或数个氨基酸残基。但是对于SPAG-9尚未有相关研究。本研究利用生物信息软件系统等预测分析了SPAG-9的B细胞优势表位以及人主要组织相容性复合体(major histocompatibility complex,MHC)限制的细胞毒性T淋巴细胞(cytotoxic T lymphocyte,CTL)表位,首次得到了SPAG-9的表位富集区,为SPAG-9表位疫苗的设计和研究提供理论依据。
1 材料和方法
1.1 SPAG-9优势B细胞表位预测
1.1.1 SPAG-9的氨基酸序列:SPAG-9的氨基酸全长序列检索自SwissProt蛋白质数据库。
1.1.2 SPAG-9的二级结构预测:利用EXPASY服务器提供的GOR、SOPMA、Scratch Protein Predictor(SPP)方法(http://www.expasy.ch/tools)对SPAG-9全长氨基酸序列的二级结构进行预测分析。
1.1.3 SPAG-9跨膜区域预测:利用CBS服务器上的TMHMM方法(http://www.cbs.dtu.dk/services/TMHMM/)对SPAG-9全长氨基酸序列的跨膜区域进行预测分析。

表1 GOR4、SPP、SOPMA 3种方法预测SPAG-9二级结构的构成比[ n(%)]
1.1.4 SPAG-9亲水性、极性、表面可及性、抗原性及柔韧性参数预测:综合EXPASY服务器提供的Hopp&Woods(亲水性参数)、Zimmerman(极性参数)(http://web.expasy.org/cgi-bin/protscale/protscale.pl)与DNAstar软件的Protein进行的Emini(表面可及性参数)、Jameso-Wolf(抗原性参数)与Karplus-Schulz(柔韧性指数)分析来对SPAG-9的B细胞表位进行预测。
1.1.5 SPAG-9抗原性指数分析:结合上述预测结果,运用吴玉章等[10]建立的抗原性指数确定SPAG-9全长氨基酸的优势B细胞表位富集区。
1.2 SPAG-9 T细胞表位预测 应用SYFPEITHI服务器上的EPITOPE PREDICTION方法分析预测SPAG9的CTL表位,选取评分较高(score≥20)的多肽。应用NetCTL预测HLA-A2及HLA-A24限制的CTL表位获得HLA-A2高评分(threshold for epitope identification≥0.75)的多肽和HLA-A24高评分的多肽。应用IEDB预测HLA-A2、HLA-A24限制的CTL表位,选取高评分的多肽(percentile_rank<5)。综合SYFPEITHI、Net-CTL及IEDB 3种方法,选取均高评分的HLA-A2限制以及HLA-A24限制的CTL表位肽。
1.3 SPAG-9表位富集区预测 综合上述预测得到的优势B细胞表位和HLA限制的CTL表位,得到SPAG-9的表位富集区。
2 结果
2.1 SPAG-9全长氨基酸序列 SPAG-9共1 321个氨基酸,相对分子质量146 kDa。
2.2 SPAG-9优势B细胞表位预测结果
2.2.1 SPAG-9的二级结构预测:GOR4、SOPMA、SPP 3种方法预测SPAG-9的二级结构,均提示SPAG-9的二级结构以无规则卷曲为主,亦可见30%~40%的α-螺旋,而少见β-折叠(见表1)。在这3种方案的预测结果中,以至少2种方案相一致的重叠区域为准,可见无规则卷曲主要位于N端的1-5,11-17,42-43,182-186,197-232,236-244,2 468-255,268-284,288-300,306-311,323-335,347-372,376-378,391-392,396,443,488-496,539-542,547-551,561-575,578-604,640-642,646-655,658-672,675-684,695-705,715-727,763-768,775-780,788-790,797-819,826-835,840-841,846-887,896-905,910-913,920-934,936-943,951-954,969-970,976-979,986-989,995,998-1 002,1 010-1 011,1 018-1 020,1 028-1 038,1 044-1 049,1 058,1 064-1 066,1 072-1 076,1 081-1 088,1 099-1 100,1 108-1 109,1 117-1 119,1 124-1 129,1 136-1 144,1 154-1 156,1 162-1 165,1 171-1 187,1 194-1 215,1 219-1 225,1 232,1 234-1 235,1 238-1 266,1 274-1 277,1283-1 289,1 293-1 304,1 321区域。
2.2.2 SPAG-9跨膜区的预测:使用TMHMM方法对其跨膜区进行预测分析,结果显示SPAG-9为膜外蛋白,无跨膜区。
2.2.3 SPAG-9亲水性、极性、表面可及性、抗原性及柔韧性参数预测:SPAG-9的亲水性、极性、表面可及性、抗原性参数、柔韧性参数的预测详见图1和表2,综合各种参数的预测结果,SPAG-9 B细胞表位所在的优势区段为197-200,227-232,237-244,249-255,292-298,306-311,323-327,332-335,348-350,353-358,488-494,539-542,547-550,565-572,588-595,601-604,661-667,677-684,698-702,722-727,801-808,816-819,869-873,875-881,884-887,938-943,1173-1177,1194-1201(抗原性指数≥0,亲水性指数≥0,表面可及性指数≥1,极性指数≥12)。
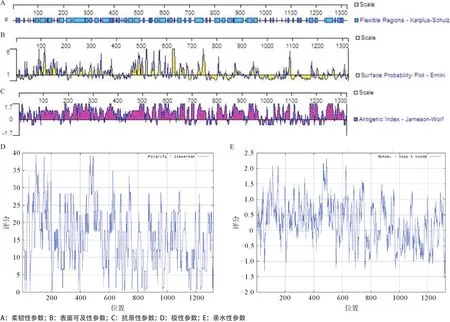
图1 SPAG-9不同参数的预测结果
2.2.4 SPAG-9抗原性指数分析:综合SPAG-9的二级结构、亲水性、表面可及性、极性、抗原性参数、柔韧性参数的预测一致区域,计算SPAG-9的B细胞表位区域的平均抗原性指数,结果显示N端的197-201,236-243,249-253,291-299,306-312,324-327,332-337,348-352,488-494,539-542,546-550,565-573,699-702,802-807,869-873,875-880,882-887,1 173-1 178,1 195-1 200区段的抗原性指数相对较高,可能为SPAG-9优势B细胞表位,见表3。
2.3 SPAG-9 T细胞表位预测结果 综合SYFPEITHI、Net-CTL及IEDB 3种方法均高评分的HLA-A2限制的CTL表位肽为N端的19-27,49-57,50-58,56-64,343-351,381-389,413-421,447-455,521-529,665-673,733-741,837-845,926-934,950-958,964-972,998-1 006,1 016-1 024,1 094-1 102,1 140-1 148,1 150-1 158(见表4)。均高评分的HLA-A24限制的CTL表位肽为31-39,137-145,517-525,943-951(见表5)。
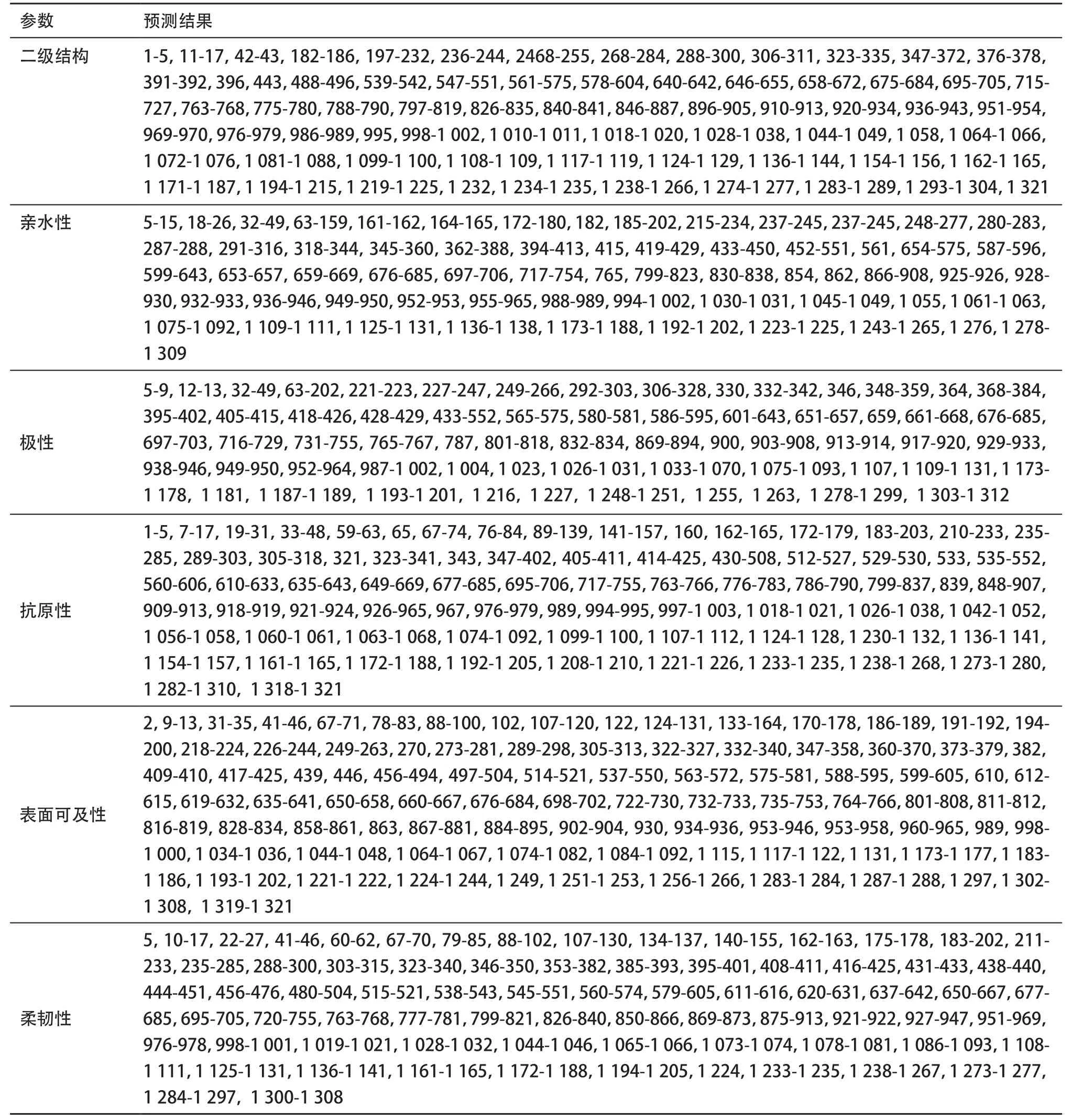
表2 不同参数预测SPAG-9蛋白B细胞表位的肽段区域
2.4 SPAG-9表位富集区预测结果 综合SPAG-9 B细胞表位和CTL表位的预测结果,SPAG-9表位富集区为N端的332-352,551-573,见图2。
3 讨论
近年来,随着基因组学、蛋白质组学的飞速发展,对于肿瘤的治疗不仅仅局限于手术治疗以及放化疗治疗,肿瘤特异性治疗开始受到广泛关注。众所周知,人体免疫系统通过免疫监视功能识别和清除突变细胞维持机体的生理平衡稳定,而免疫系统所能识别的就是癌变细胞表达的肿瘤抗原,包括肿瘤相关抗原或肿瘤特异性抗原。研究表明仅15.4%的肿瘤为病原体持续感染所引起,所以肿瘤抗原将是肿瘤特异性免疫治疗的理想靶点[11]。
抗原表位是抗原分子中决定抗原特异性的一小段序列,能够被机体免疫效应细胞特异性识别并诱导后续的免疫反应。而且作为肿瘤特异性治疗的靶点,能够避免天然抗原中对机体产生不利影响的抗原成分,诱导具更强的特异性和更高的免疫效应的特异性免疫应答。以此类特异性抗原表位为基础所研究的疫苗分子已经展现出了良好的研究前景。SHEN等[12]通过对MAGE-3 B细胞表位的预测和原核表达表位多肽,免疫小鼠之后成功地诱发了小鼠体液免疫。另外,LIU等[13]将预测的Ewing’s肉瘤的B细胞表位免疫新西兰白兔,ELISA检测发现新西兰白兔产生了良好的抗体滴度。最近也有研究表明携带CTL表位的DNA疫苗能诱发机体产生很强的针对该表位特异性的CTL反应,通过释放胞浆内的颗粒(含穿孔素和颗粒酶)等,达到杀伤和清除肿瘤细胞的目的[14-15]。
SPAG-9为一种较新的肿瘤特异膜蛋白,目前尚未见研究以此作为肿瘤特异性免疫治疗的靶点。本研究综合生物信息学方法预测出来的B细胞表位和CTL表位,筛选出的表位富集区包含多个B细胞表位和CTL表位,其既能够被B细胞受体以及抗体特异性识别,诱导B细胞活化产生特异性抗体,通过激活补体,介导ADCC作用和发挥调理作用发挥抗肿瘤效应[16],也可以被T细胞受体特异性识别,诱导产生特异性CTL,通过释放细胞毒颗粒介导细胞毒性反应,引起肿瘤细胞凋亡[17]。而且,不同于以往的单一抗原表位,本研究得到的表位富集区包含多个抗原表位,大大提高了其作为肿瘤特异性免疫治疗靶点的有效性。
对于B细胞表位的预测,本研究综合二级结构、亲水性、抗原性、表面可及性、柔韧性等参数,并将预测结果与与吴玉章等[10]建立的抗原指数相结合,从而对做出科学、合理的预测分析。通常,亲水性和表面可及性是形成B细胞表位的首要条件,但决定表位是否具有抗原性尚需综合考虑多种因素,蛋白质序列中肽链的柔韧性与相应部位的抗原性也密切相关。另外,蛋白质的二级结构对抗原表位也有很大影响。α-螺旋、β-折叠的化学键键能比较高,在牢固维持蛋白质高级结构上发挥重要的作用,但是却很难合适地与抗体嵌合,且经常处于蛋白质的内部,因而很少位于抗原表位所在区域。相反,β-转角及无规则卷曲则比较松散,且易于发生扭曲盘旋,并展示在蛋白质的表面,成为抗原表位的可能性较大。另外,亲水性较高的区域可使抗原抗体更好地结合,柔韧性较强的氨基酸残基更易形成抗原表位。所以综合上述方法,本研究结果发现SPAG-9氨基酸序列N端的197-201,236-243,249-253,291-299,306-312,324-327,332-337,348-352,488-494,539-542,546-550,565-573,699-702,802-807,869-873,875-880,882-887,1 173-1 178,1 195-1 200综合评分较高。

表3 SPAG-9的B细胞表位的平均抗原性指数
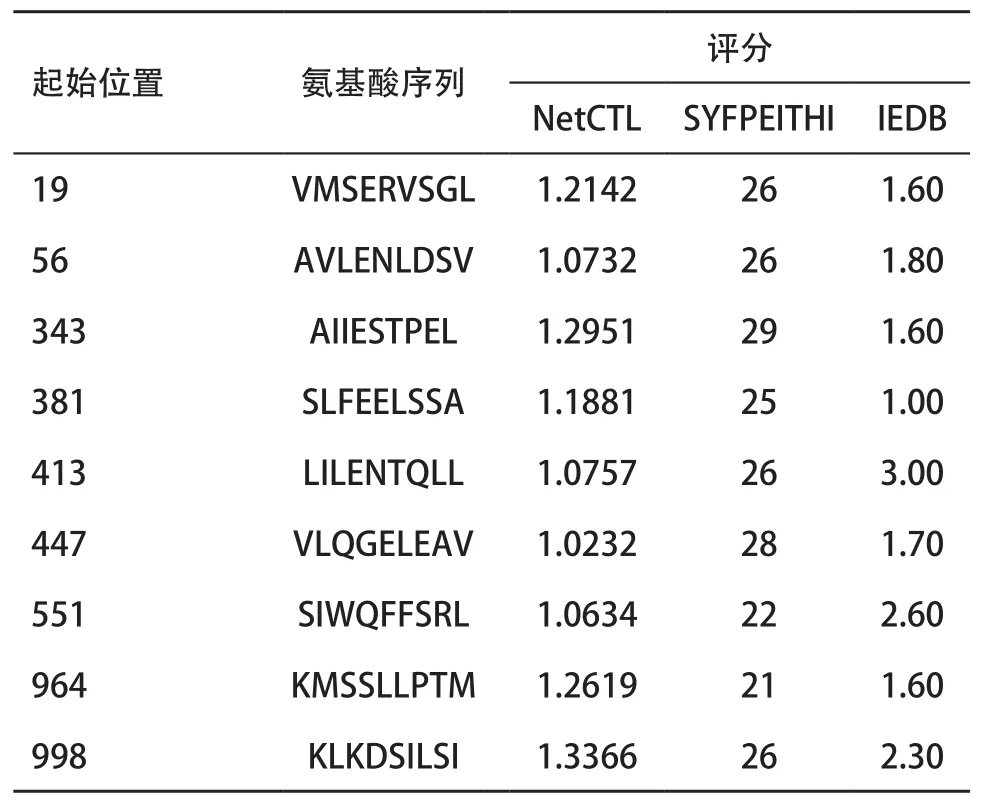
表4 SPAG-9 HLA-A2限制CTL表位3种方法评分均高的预测结果
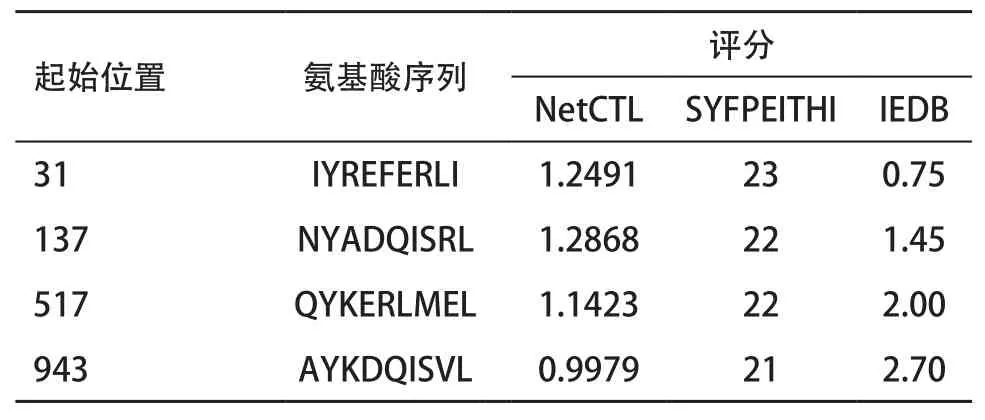
表5 SPAG-9 HLA-A24限制CTL表位3种方法评分均高的预测结果

图2 SPAG-9表位富集区预测结果
而对于T细胞表位的预测,基于已经有研究发现HLA-A2是我国人群的优势基因(HLA-A24在我国人群也有较高的频率分布[18],本研究综合SY-FPEITHI、Net-CTL及IEDB 3预测方法,筛选HLA-A2限制性的CTL表位19-27,49-57,50-58,56-64,343-351,381-389,413-421,447-455,521-529,551-559,665-673,733-741,837-845,926-934,950-958,964-972,998-1 006,1 016-1 024,1 094-1 102,1 140-1 148,1 150-1 158,A24限制性的CTL表位为31-39,137-145,517-525,943-951。
综合上述预测结果,优先考虑HLA-A2限制的T细胞表位,我们发现SPAG-9氨基酸序列N端332-352,551-573富含B、T细胞表位,即SPAG-9氨基酸序列N端332-352,551-573为其表位富集区,这能够为下一步SPAG-9的表位疫苗的研究提供理论基础。
猜你喜欢
农业工程学报(2022年8期)2022-08-08
化学工业与工程(2022年1期)2022-03-29
数学物理学报(2021年6期)2021-12-21
江苏农业科学(2020年24期)2020-02-22
温州医科大学学报(2019年4期)2019-04-28
中国免疫学杂志(2017年1期)2017-01-17
中国实用医药(2016年14期)2016-05-20
橡塑技术与装备(2016年7期)2016-02-25
中国学术期刊文摘(2016年2期)2016-02-13
畜牧兽医学报(2015年3期)2015-07-05
