
LAZY1通过油菜素内酯途径调控水稻叶夹角的发育
2018-01-22 02:03:07张晓琼王晓雯田维江张孝波李杨羊何光华桑贤春
作物学报 2017年12期
张晓琼 王晓雯 田维江 张孝波 孙 莹 李杨羊 谢 佳 何光华 桑贤春*
通过油菜素内酯途径调控水稻叶夹角的发育
张晓琼 王晓雯 田维江 张孝波 孙 莹 李杨羊 谢 佳 何光华 桑贤春*
西南大学水稻研究所 / 转基因植物与安全控制重庆市重点实验室, 重庆 400716
叶夹角的大小直接影响水稻叶面积指数, 进而调控群体光合作用, 是水稻株型育种中重要的指标, 研究其发育机制对水稻株型育种具有重要的意义。利用EMS诱变籼型水稻保持系西大1B, 获得一个植株散生且叶夹角变大的突变体。田间种植条件下, 苗期的叶夹角极显著大于野生型; 分蘖期的分蘖角极显著增大, 株型松散; 成熟期整个植株呈匍匐状生长。而野生型株型在整个生育期均保持相对紧凑, 叶夹角较小。石蜡切片分析显示,叶夹角增大是由叶枕近轴面细胞变大造成的。的主要农艺性状与野生型相比无明显变化。遗传分析表明该性状受1对隐性核基因控制, 利用SSR标记进行基因定位, 最终将定位在第11染色体标记RM4746和RM26742之间324 kb的物理范围内, 包含散生基因。测序结果显示突变体在第3外显子上发生了一个T到C的碱基替换, 导致第143位氨基酸由野生型的缬氨酸突变为丙氨酸, 表明是一个新的等位突变体。对外源油菜素内酯(BR)的敏感性降低, BR信号传导途径关键基因在中的表达上调了近10倍, 早期研究表明基因的过表达可导致叶夹角变大。推测可能通过BR信号传导途径调控水稻叶夹角的发育。
水稻(L.); 叶夹角;; 油菜素内酯(BR)信号传导;
水稻产量除受外界不可控的环境因素影响外, 还与水稻的株型相关, 而叶夹角和分蘖角度是水稻株型的重要组成。叶夹角和分蘖角度通常会影响植物的光合效率以及植株间的温度和气体的含量, 是水稻品种选育、栽培研究和生产实践上的重要性状。通常认为分蘖角度属于数量性状, 已定位[1]、[2]、[2]、[3]等多个主效QTL, 并克隆了[4]。也有研究表明分蘖角为质量性状, 并克隆了其调控基因[5-6]和[7]。表达具有时空性, 主要在分蘖基部不伸长节、茎的节部与叶鞘交接部位的叶鞘枕处表达, 属于发育时期调控基因, 其过量表达可导致分蘖角度增大[4];通过调节生长素极性运输影响水稻地上部的向重性, 最终控制分蘖角的大小[5-6], 属于重力信号传导途径基因;是油菜素内酯(BR)响应途径中的负向调节子, 抑制的表达, 会导致叶夹角和分蘖角度明显增加[7]。
叶夹角对外界的信号比较敏感, 光、植物激素等都可能改变其大小。目前研究认为, 叶夹角主要与植物激素有关, 已克隆了[8]、[9]、[10]等多个基因; 另外叶夹角还受重力响应和叶枕机械强度等因素的影响, 相关基因如[11]、[12]等。对生长素不敏感, 但对外源BR异常敏感, 可通过BR的合成和信号传导途径调节叶枕近轴面细胞大小, 影响叶夹角发育[8];的表达受脱落酸、赤霉素、生长素、BR等多激素的诱导[9], 控制近轴面细胞的分裂速度, 最终影响叶夹角大小;介导生长素与BR信号传导通路的交叉网络, 共同增强叶枕近轴面细胞的分裂[10];编码一个具有独特保守特征的ID蛋白, 调控淀粉质体沉降率,进而影响胚芽鞘的重力感应和信号转导[11];在叶枕维管束中表达, 其与一个核蛋白家族的互作调节叶枕机械组织的形成, 影响叶枕机械强度, 调控叶夹角[12]。这些叶角度、分蘖角度相关基因的定位克隆与功能研究为揭示水稻株型发育的分子机制奠定了良好基础, 但仍有待更深入的研究。
从EMS诱变籼型水稻保持系西大1B后代中鉴定到一个叶角度变大的散生突变体, 我们对其进行了形态鉴定、细胞学分析和基因精细定位等研究, 发现它是新的等位突变体, 通过BR信号传导途径影响叶夹角发育。
1 材料与方法
1.1 试验材料
连续种植2代,突变性状均能稳定遗传。
1.2 农艺性状鉴定
田间种植和常规管理条件下, 全生育期观察野生型和的株型变化。成熟期随机选择中间10株, 测量株高和节间长, 并调查穗长、有效穗、穗粒数、穗实粒数、结实率、千粒重等主要农艺性状。
1.3 组织表型分析
取三叶期野生型和突变体全展叶的叶枕, 依据文献[13]稍作改动进行石蜡切片分析。即FAA固定液固定后, 依次用一定梯度浓度的乙醇脱水、二甲苯透明、石蜡包埋、切片、烘干和番红固绿染色, 最后中性树胶封片, 显微镜下观察并拍照。
1.4 遗传分析和定位群体构建
在西南大学水稻研究所歇马基地配制日本晴/杂交组合, 同年秋在海南加代种植F1, 观察植株形态并收获F2种子。次年夏在西南大学水稻研究所歇马基地种植杂交组合的亲本和F2群体, 进行遗传分析; 同时, 利用日本晴/杂交组合的F2隐性单株进行基因定位。
1.5 DNA的提取
采用改进后的CTAB法[14]提取亲本、F2正常植株基因池、F2突变株基因池及定位群体单株的基因组DNA。
1.6 PCR分析
根据日本晴和1B的DNA序列差异, 参照http://www.gramene.org/microsat/等网站设计SSR和InDel标记。PCR体系总体积12.5 μL, 包括1.0 μL的模板DNA, 1.0 μL的10 μmolL–1引物, 0.65 μL的25 mmolL–1MgC12, 0.5 μL 2.5 mmolL–1dNTPs, 8.0 μL ddH2O, 1.25 μL的10 × PCR 缓冲液和0.1 μL的5 U μL–1DNA聚合酶。PCR程序为94℃预变性5 min, 94℃变性20 s, 56℃退火20 s, 72℃延伸40 s, 35个循环后, 72℃再充分延伸10 min。PCR产物经10%的非变性聚丙烯酰胺凝胶电泳, 0.1%质量浓度的AgNO3染色10 min, 去离子水漂洗2次, 再以1%质量浓度NaOH和0.1%体积质量浓度甲醛的混合液显色, 观察记录带型并照相。
1.7 BR敏感实验
取野生型和突变体种子, 催芽露白后播种至营养土中, 分别进行黑暗和正常培养。至三叶一心期时, 调查长势正常的野生型和突变体植株的叶夹角, 进行统计分析。同时, 剪取正常培养的野生型和突变体植株叶枕处前后2 cm左右的材料, 分别置0、0.01、0.1、1和10 μmol L–124-表油菜素内酯(24-eBL)溶液中, 暗培养3 d后统计各浓度下叶夹角大小。以上实验均3次生物学重复。
1.8 RNA提取及qPCR
三叶一心期, 分别取野生型和突变体植株的叶枕, 放进无核酸酶的2 mL离心管中, 液氮速冻后研磨成粉, 根据试剂盒描述的方法提取RNA并反转录成cDNA, 试剂盒购自天根生化科技(北京)有限公司, 然后再参照文献[13]的方法对BR信号传导途径相关基因[15]、[16]、[17]、[18]和BR合成相关基因[19]、[20]进行qPCR定量分析。qPCR引物序列见表1。
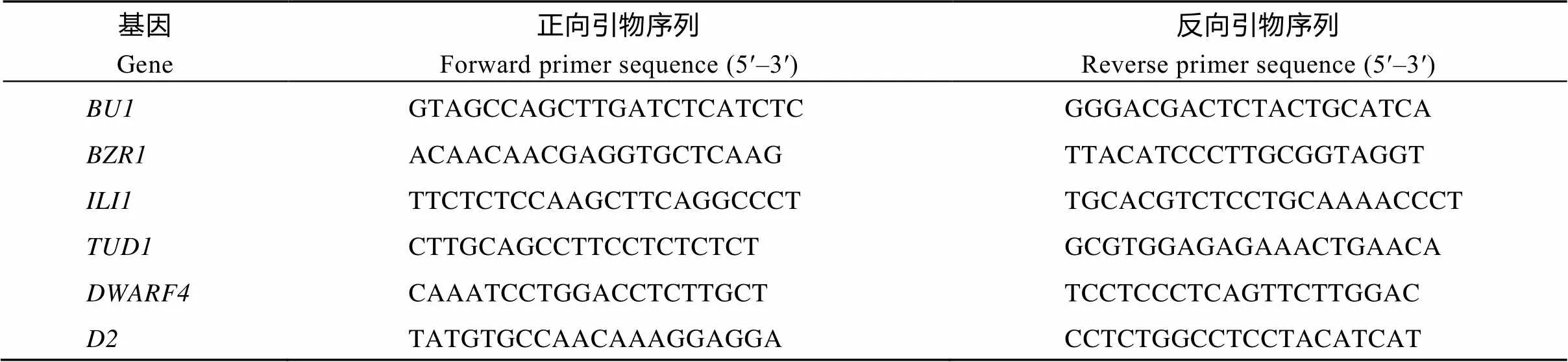
表1 qPCR定量引物序列
2 结果与分析
2.1 表型鉴定
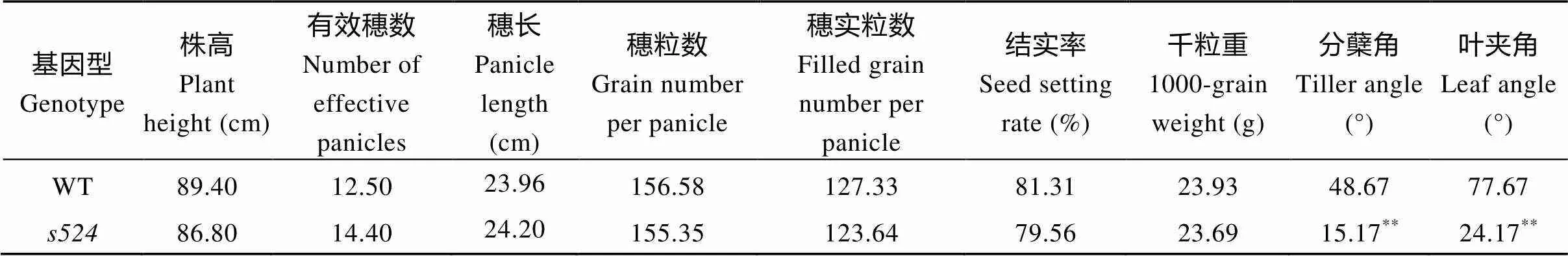
田间种植条件下, 突变体整个生育期均表现叶夹角增大和植株散生。三叶期, 野生型的叶夹角为10.31°,的则是41.62°, 相比野生型, 突变体的叶夹角增大4倍, 达极显著差异水平; 进入分蘖期,的分蘖角度显著大于野生型, 且随生长发育进程变得更大(图1-A, B); 灌浆期整个植株呈匍匐状(图1-D)。而野生型西大1B整个生育期株型均保持紧凑, 分蘖角度正常(图1-C)。与野生型相比,除叶夹角和分蘖角度显著变大外, 株高、有效穗、穗长、穗粒数、穗实粒数、结实率、千粒重等农艺性状均无显著变化(表2)。
2.2 细胞学分析
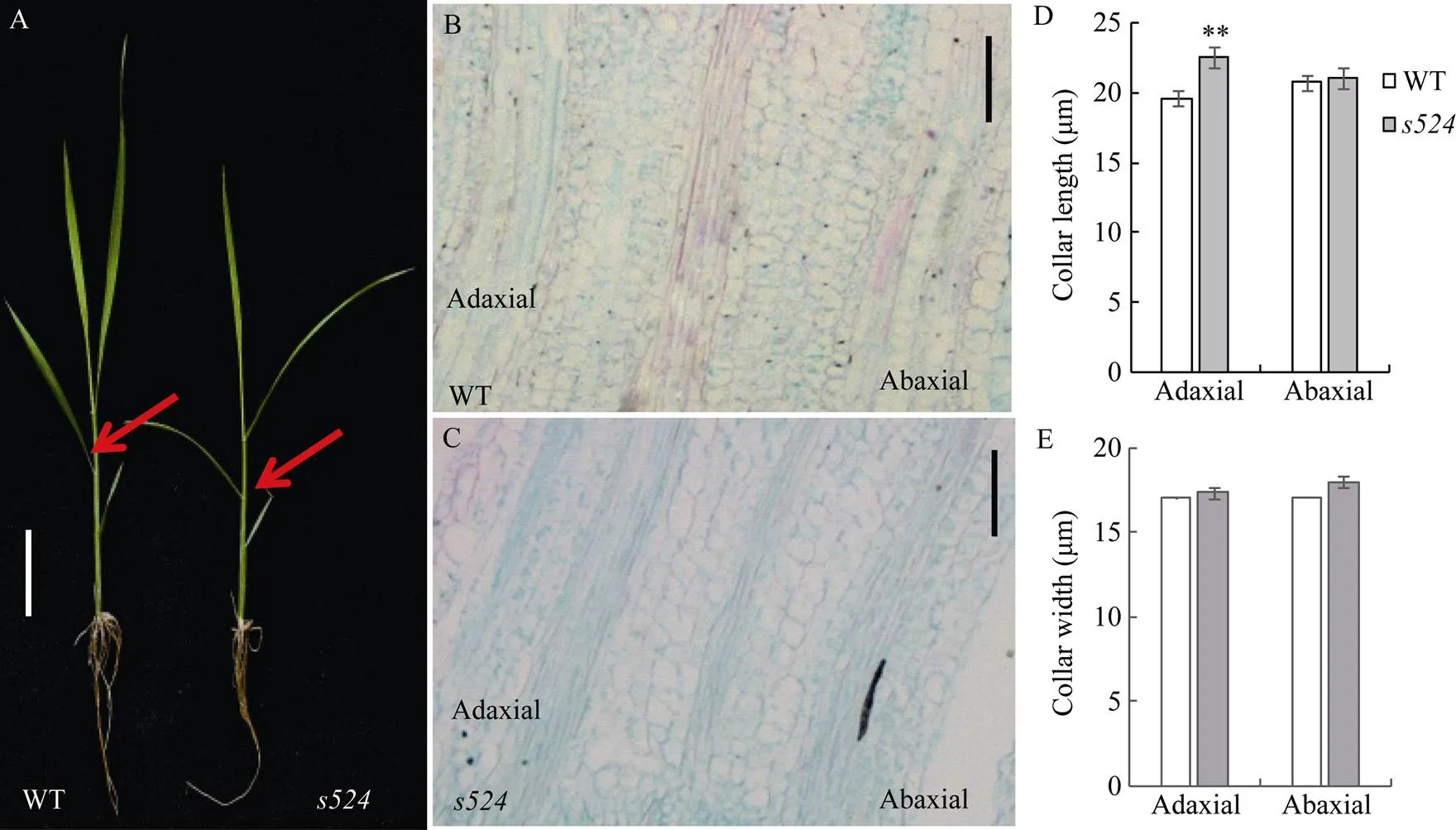
形态学观察表明的叶夹角变大, 为进一步分析其原因, 利用石蜡切片观察了三叶期倒一全展叶叶枕部位的细胞结构(图2-A)。结果显示, 野生型(图2-B)近轴面细胞长度平均为19.63 μm, 突变体(图2-C)的约为22.49 μm, 即相对于野生型, 突变体近轴面细胞长度增加10%, 达显著差异水平; 野生型植株远轴面细胞长度为20.72 μm, 相应的突变体的约为21.01 μm, 比野生型增加1%, 差异不显著(图2-D); 野生型与突变体的近轴面细胞宽度分别为17.08 μm和17.38 μm, 远轴面细胞宽度分别是17.11 μm和18.01 μm (图2-E), 细胞宽度变化均不显著。由此推测, 突变体叶夹角变大可能是由近轴面细胞变长导致的。
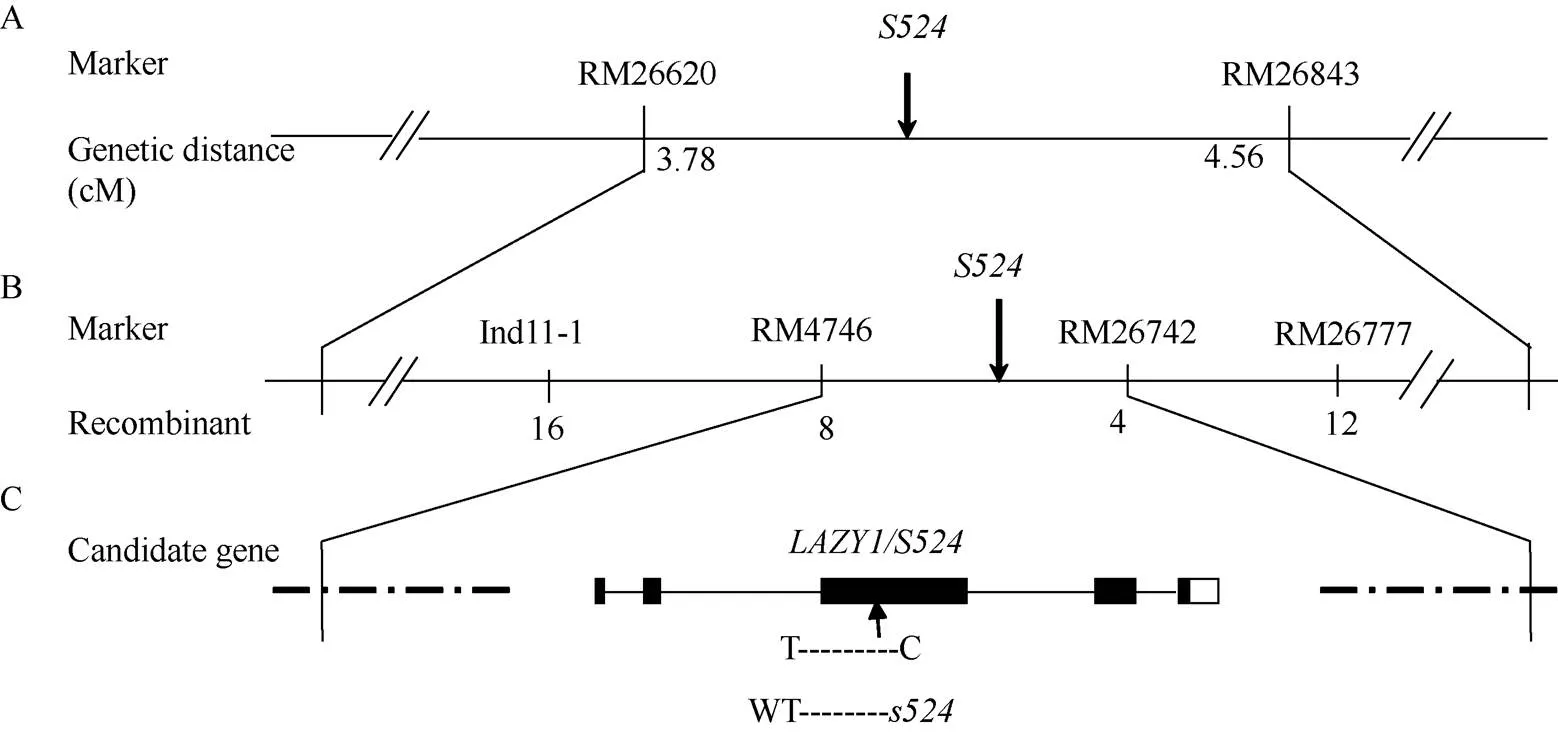
2.3 基因的精细定位
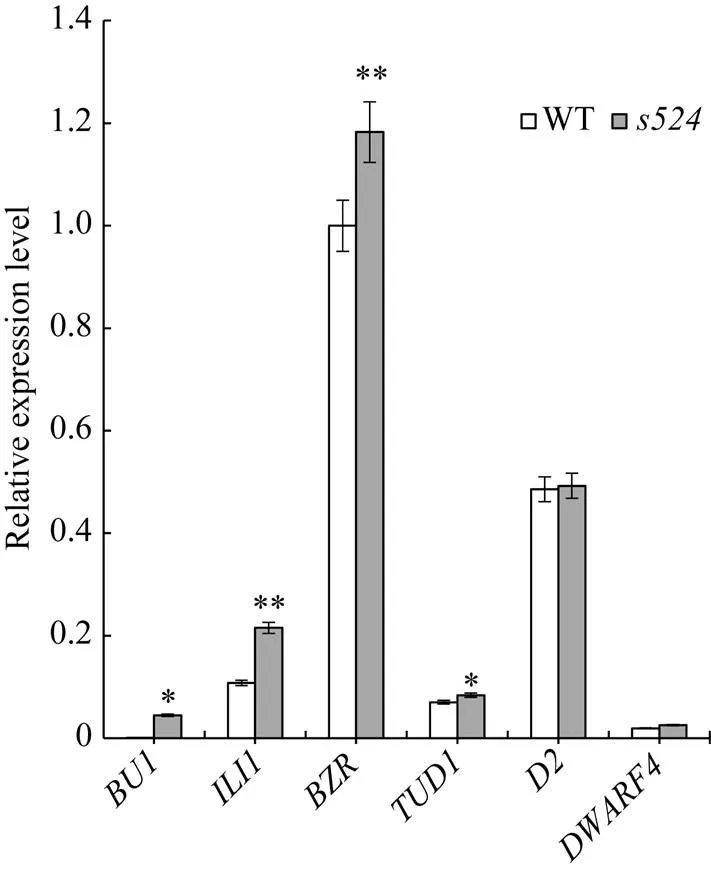
日本晴/杂交组合的F1群体正常; F2群体中有2564株正常单株和826株突变单株, 突变单株均表现为散生和叶夹角变大, 卡方检验(c2=0.73 图1 突变体s524和野生型的表型观察 A: 分蘖期野生型, Bar=10 cm; B: 分蘖期突变体, Bar=10 cm; C: 灌浆期野生型, Bar=15 cm; D: 灌浆期突变体, Bar=15 cm; E: 为图1C和图1D灌浆期放大的分蘖基部。 A: plant phenotype of the wild type at tillering stage, Bar=10 cm; B: plant phenotype of theat tillering stage, Bar=10 cm; C: plant phenotype of the wild type at filling stage, Bar=15 cm; D: plant phenotype of theat filling stage, Bar=15 cm; E: magnified tiller base at filling stage in Figs. 1C and 1D. 表2 突变体s524与野生型(WT)的农艺性状 **在0.01水平上差异显著;=10。 **Significant difference at<0.01 by-test;=10. 图2 近轴面细胞变长导致s524叶夹角变大 A: 三叶一心期野生型(WT)和突变体, 箭头所指为叶枕部位, Bar=10 cm; B, C: 野生型(B)和突变体(C)的叶枕细胞, Bar=50 μm; D, E: 野生型和突变体近轴端与远轴端细胞长度(D)和宽度(E)统计图。3次生物学重复;=50。**在0.01水平上差异显著(测验)。 A: plant phenotype of the wild type(WT) and mutant type at stage with three whole and one under-developing leaf blades, Bar=10 cm; B and C: comparison of the leaf occipital cells of WT(B) andmutant(C), Bar=50 μm; D and E: statistical chart of length (D) and width (E) of the adaxial and abaxial cells in WT and. The experiments are biologically repeated three times and the data are presented as means±SE,=50. ** Significant difference at<0.01 by-test. 油菜素内酯(BR)是一种高效的植物生长调节剂, 广泛存在于花、茎和根部, 生理活性极强, 是植物第六大激素, 尤其在单子叶植物叶夹角发育中具有重要功能[21]。不同植株对BR的敏感程度不同, 引起的叶夹角变化也不相同。随着外施24-eBL浓度的升高, 野生型和突变体的叶夹角都随之变大; 当浓度增大至10 μmol L–1时, 野生型的夹角度数继续增大, 而突变体的则保持不变(图4-A)。在1 μmol L–1浓度下, 突变体叶夹角即达到最大值, 其叶夹角变化量(∆S)为野生型(∆B)的60%, 达极显著差异水平(图4-B)。这表明对BR敏感性降低。 qPCR分析表明, 与野生型相比, BR信号传导途径基因、、和在突变体中的表达量均极显著或显著上调, 其中,在中的表达量近乎为野生型的10倍; 但BR合成途径调控基因和在突变体和野生型之间的表达变化则未达到统计学差异水平(图5)。 图3 LAZY1/S524在水稻第11染色体上的分子定位 图4 s524叶夹角对外源BR的敏感性降低 A: 野生型和突变体叶夹角对激素BR的敏感性检测; B: 野生型和突变体叶夹角变化。∆B和∆S分别为1 μmol L–1外源24-eBL处理前后野生型和突变体叶夹角的差值。**在0.01水平上差异显著(测验)。 A:BR sensitivity tests of the wild type and mutant type; B:comparison of the leaf inclination of WT andmutant. ∆B and ∆S represent the difference between the angle of the wild type and the mutant respectively, under 1 μmol L–124-eBL treatment. ** Significant difference at< 0.01 by-test. 叶夹角和分蘖角是水稻理想株型育种的重要选择指标, 本文EMS诱变鉴定了一个散生且叶夹角变大的突变体, 精细定位及候选基因分析表明其为的等位突变体。目前已经报道了5个的等位突变体,在第4外显子上有8个碱基的缺失, 导致蛋白质翻译提前终止[5];的开放阅读框则整体丢失[5];、和为T-DNA插入突变体, 在基因内部发生了大片段的插入或/和缺失[6]。突变体仅在第3外显子区域发生一个碱基的替换, 导致氨基酸的转换, 是一个弱等位突变体, 其散生性状与已报道的等位突变体类似, 但株高、有效穗和穗粒数等农艺性状无显著变化。 突变体除植株散生外, 另一典型特征是叶夹角变大。目前已报道了[8]、[21]、[22]等多个叶夹角发育调节基因。其中编码一个生长素氨基合成酶GH3-1, 通过增加近轴面细胞的长度调控叶夹角大小[8];编码一个类VIL3蛋白, 叶枕近轴面上表皮细胞分裂能力增强导致叶角变大[9];是一个细胞周期调控基因, 表达量降低致使水稻叶片远轴面细胞数量减少, 进而叶夹角变大[23];的功能缺失会导致表达上调(编码一个木葡聚糖内糖基转移酶, 是细胞伸长过程中使细胞壁松弛所必需的酶), 进而引起植株叶枕近轴面细胞膨大使叶夹角增大[22]。与已报道的突变体类似,叶角处远轴面细胞与野生型相比无明显变化, 近轴面细胞则极显著变长, 从而导致突变体叶夹角变大。 图5 与叶夹角相关BR合成与信号传导途径中的基因在野生型(WT)和突变体s524的表达分析 基因作为内参, 3次生物学重复。**在0.01水平上差异显著; *在0.05水平上差异显著(测验)。 Comparison of the expression of genes related to leaf angle in BR signalling. Usinggene as a reference and the experiments are biologically repeated three times. ** Significant difference at< 0.01 by-test; * significant difference at<0.05 by-test. 油菜素内酯是调控单子叶植物叶夹角发育的重要激素。油菜素内酯响应实验表明对BR的敏感性降低, 暗示可能参与BR的信号传导途径。qPCR分析显示, BR生物合成基因、在和野生型中的表达差异不显著, BR信号传导基因和在突变体中的表达量均上调, 进一步佐证了通过BR信号传导途径调控水稻叶夹角发育的推测。 是BR信号途径中的早期应答基因, 参与和介导的BR信号传导通路。抑制的表达使水稻叶片出现直立的表型, 而过量表达该基因则导致叶夹角变大[15]。本研究发现,在突变体中的表达量与野生型相比升高了近10倍(图5)。因此推测可能通过影响等BR信号传导上游通路基因调控叶枕近轴面细胞的发育, 进而影响水稻叶夹角的大小。 全生育期均表现叶夹角变大和植株散生, 农艺性状无显著变化。叶枕近轴面细胞变长是导致突变体叶夹角变大的主要原因。该性状受单隐性核基因调控, 定位在第11染色体SSR标记RM4746和RM26742之间约324 kb的物理范围内。是的新等位突变体, 在第3外显子上产生了一个T到C的碱基替换, 致使第143位氨基酸由野生型的缬氨酸变为了突变体的丙氨酸。对BR的敏感性降低, 上游途径基因的表达上调了近10倍, 推测可能通过影响BR信号传导通路调控水稻叶夹角的发育。 [1] Li Z K, Paterson A H, Pinson S R, Stansel J W. RFLP facilitated analysis of tiller and leaf angles in rice (L.)., 1999, 109: 79–84 [2] 钱前, 何平, 滕胜, 曾大力, 朱立煌. 水稻分蘖角度的QTLs分析. 遗传学报, 2001, 28: 29–32 Qian Q, He P, Teng S, Zeng D L, Zhu L H. QTLs analysis of tiller angle in rice (L.)., 2001, 28: 29–32 (in Chinese with English abstract) [3] 余传元, 刘裕强, 江玲, 王春明, 翟虎渠, 万建民. 水稻分蘖角度的QTL定位和主效基因的遗传分析. 遗传学报, 2005, 32: 948–954 Yu C Y, Liu Y Q, Jiang L, Wang C M, Zhai H Q, Wan J M. QTLs mapping and genetic analysis of tiller angle in rice (L.)., 2005, 32: 948–954 (in Chinese with English abstract) [4] Yu B, Lin Z, Li H, Li X, Li J, Wang Y, Zhang X, Zhu Z, Zhai W, Wang X, Xie D, Sun C., a major quantitative trait locus controlling tiller angle in rice., 2007, 52: 891–898 [5] Li P, Wang Y, Qian Q, Fu Z, Wang M, Zeng D, Li B, Wang X, Li J.controls rice shoot gravitropism through regulating polar auxin transport., 2007, 17: 402–410 [6] Yoshihara T, Lino M. Identification of the gravitropism-related rice geneand elucidation ofdependent and independent gravity signaling pathways., 2007, 48: 678–688 [7] Zhang C, Xu Y Y, Guo S Y, Zhu J Y, Huan Q, Liu H H, Wang L, Luo G Z, Wang X J, Chong K. Dynamics of brassinosteroid response modulated by negative regulator LIC in rice., 2012, 8: e1002686 [8] Zhao S Q, Xiang J J, Xue H W. Studies on the rice LEAF INCLINATION1 (LC1), an IAA-amido synthetase, reveal the effects of auxin in leaf Inclination control., 2013, 6: 174–187 [9] Zhao S Q, Hu J, Guo L B, Qian Q, Xue H W. Rice leaf inclination2, a VIN3-like protein, regulates leaf angle through modulating cell division of the collar., 2010, 20: 935–947 [10] Zhang S N, Wang S K, Xu Y X, Yu C L, Shen C J, Qian Q, Geisler M, Jiang D A, Qi Y H. The auxin response factor, OsARF19, controls rice leaf angles through positively regulating OsGH3-5 and OsBRI1., 2015, 38: 638–654 [11] Wu X R, Tang D, Li M, Wang K J, Cheng Z K. Loose plant architecture 1, an indeterminate domain protein involved in shoot gravitropism, regulates plant architecture in rice., 2013, 161: 317–329 [12] Ning J, Zhang B C, Wang N L, Wang N, Zhou Y H, Xiong L Z. Increased leaf angle 1, a Raf-Like MAPKKK that interacts with a nuclear protein family, regulates mechanical tissue formation in the lamina joint of rice., 2011, 23: 4334–4347 [13] Sang X C, Li Y F, Luo Z K, Ren D Y, Fang L K, Wang N, Zhao F M, Ling Y H, Yang Z L, Liu Y S, He G H. CHIMERIC FLORAL ORGANS1, encoding a monocot-specific MADS box protein, regulates floral organ identity in rice., 2012, 160: 788–807 [14] Porebski S, Bailey L G, Baum B R. Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components., 1997, 15: 8–15 [15] Tanaka A, Nakagawa H, Tomita C, Shimatani Z, Ohtake M, Nomura T, Jiang C J, Dubouzet J G, Kikuchi S, Sekimoto H, Yokota T, Asami T, Kamakura T, Mori M. BRASSINOSTEROID UPREGULATED1, encoding a helix-loop-helix Protein, is a novel gene involved in brassinosteroid signaling and controls bending of the lamina joint in rice., 2009, 151: 669–680 [16] Bai M Y, Zhang L Y, Gampala S S, Zhu S W, Song W Y, Chong K, Wang Z Y. Functions ofand 14-3-3 proteins in brassinosteroid signaling in rice., 2007, 104: 13839–13844 [17] Zhang L Y, Bai M Y, Wu J, Zhu J Y, Wang H, Zhang Z, Wang W, Sun Y, Zhao J, Sun X, Yang H, Xu Y, Kim S H, Fujioka S, Lin W H, Chong K, Lu T, Wang Z Y. Antagonistic HLH/bHLH transcription factors mediate brassinosteroid regulation of cell elongation and plant development in rice and arabidopsis., 2009, 21: 3767–3780 [18] Hu X M, Qian Q, Xu T, Zhang Y E, Dong G J, Gao T, Xie Q, Xue Y B. The U-box E3 ubiquitin ligase TUD1 functions with a heterotrimeric G α subunit to regulate brassinosteroid-mediated growth in rice., 2013, 9: e1003391 [19] Sakamoto T, Morinaka Y, Ohnishi T, Sunohara H, Fujioka S, Ueguchi-Tanaka M, Mizutani M, Sakata K, Takatsuto S, Yoshida S, Tanaka H, Kitano H, Matsuoka M. Erect leaves caused by brassinosteroid deficiency increase biomass production and grain yield in rice., 2006, 24: 105–109 [20] Hong Z, Ueguchi-Tanaka M, Umemura K, Uozu S, Fujioka S, Takatsuto S, Yoshida S, Ashikari M, Kitano H, Matsuok M. A rice brassinosteroid-deficient mutant,, is caused by a loss of function of a new member of cytochrome p450., 2003, 15: 2900–2910 [21] Sun S, Chen D, Li X, Qiao S, Shi C, Li C, Shen H, Wang X. Brassinosteroid signaling regulates leaf erectness invia the control of a specific U-type cyclin and cell proliferation., 2015, 34: 220–228 [22] Duan K, Li L, Hu P, Xu S P, Xu Z H, Xue H W. A brassinolide- suppressed rice MADS-box transcription factor,, has a negative regulatory role in BR signaling., 2006, 47: 519–531 [23] Sakamoto T, Morinaka Y, Inukai Y, Kitano H, Fujioka S. Auxin signal transcription factor regulates expression of the brassinosteroid receptor gene in rice., 2013, 73: 676–688 Regulates Development of Rice Leaf Angle through Brassinolide Pathway ZHANG Xiao-Qiong, WANG Xiao-Wen, TIAN Wei-Jiang, ZHANG Xiao-Bo, SUN Ying, LI Yang-Yang, XIE Jia, HE Guang-Hua, and SANG Xian-Chun* Rice Research Institute of Southwest University / Chongqing Key Laboratory of Application and Safety Control of Genetically Modified Crops, Chongqing 400716, China The angle of rice leaf, as an important agronomic trait, influences leaf area index and yield in rice. Leaf angle mutant identification is of significant importance in rice breeding based on ideal plant architecture. A mutant, derived from the progeny of EMS-treatedrice Xida 1B, showed significantly larger leaf angle than the wild type. At tillering stage, the tiller angle and leaf angle ofincreased and the whole plant crept, while the wild type in whole growth period maintained relatively compact phenotype. Longitudinal sections of the epidermis of pulvinus showed that the enlarged leaf angle of the mutant was mainly due to the much-elongated adaxial cells. The main agronomic traits ofdid not change significantly compared with those of the wild-type. Genetic analysis suggested that the mutational traits were controlled by a recessive nuclear gene, which was finely mapped between SSR markers RM4746 and RM26742 on chromosome 11 with a physical distance of 324 kb.in the restricted region regulates rice tiller angle development. Sequencing revealed that a base substitution from T to C occurred on the third exon of, leading to the amino acid change from valine to alanine at 143th protein sequence. Hormone treatment test indicated that BR sensitivity was reduced in. Quantitative analysis demonstrated that the expression of BR signaling related genewas obviously up-regulated. The previous study showed that over-expression ofcould enlarge rice leaf angle. Our research suggests thatmay regulate the development of leaf angle through BR conduction pathway. Rice (L.); Tiller angle;;Signal transduction of brassinolide (BR); 10.3724/SP.J.1006.2017.01767 本研究由中央高校基本科研业务费(XDJK2015C117)和重庆市基础科学与前沿技术研究(一般)项目(cstc2015jcyjA1517)资助。 This study was supported by the Fundamental Research Funds for the Central Universities (XDJK2015C117) and Chongqing Research Program of Basic Research and Frontier Technology (cstc2015jcyjA1517). 桑贤春, E-mail: sangxianchun@163.com E-mail: zxq1964265210@163.com 2017-04-05; Accepted(接受日期): 2017-07-23; Published online(网络出版日期): 2017-08-07. http://kns.cnki.net/kcms/detail/11.1809.S.20170807.1537.002.html
2.4 油菜素内酯响应实验

3 讨论
4 结论
猜你喜欢
放射学实践(2023年9期)2023-09-20 08:24:16
哈哈画报(2022年3期)2022-05-06 09:23:31
放射学实践(2021年9期)2021-09-22 05:42:30
小学阅读指南·高年级版(2019年11期)2019-06-15 06:31:27
放射学实践(2017年5期)2017-06-01 12:19:24
阅读(科学探秘)(2017年3期)2017-05-30 10:48:04
安徽医科大学学报(2016年12期)2017-01-15 14:21:44
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
小天使·四年级语数英综合(2015年5期)2015-05-25 18:29:04
山东医药(2015年40期)2015-02-28 14:28:45
