
眉斑并脊天牛生物学特性研究
2018-01-04 18:42:32董子舒张玉静赖开平郑霞林
环境昆虫学报 2017年6期
董子舒,张玉静,赖开平,郑霞林,王 乔,陆 温*
(1.广西大学农学院,南宁 530004;2.广西化工研究院,南宁 530001;3.梅西大学农业与环境学院,新西兰北帕 11122)
眉斑并脊天牛生物学特性研究
董子舒1,张玉静1,赖开平2,郑霞林1,王 乔3,陆 温1*
(1.广西大学农学院,南宁 530004;2.广西化工研究院,南宁 530001;3.梅西大学农业与环境学院,新西兰北帕 11122)
眉斑并脊天牛GleneacantorFabricius是我国华南地区园林绿化植物木棉树的重要蛀干害虫。本试验通过室内饲养观察和野外调查,对该虫的生物学特性进行了研究。结果表明该虫在南宁一年发生4代,平均世代历期为70 d,主要以4龄幼虫在受害枝干内越冬。成虫羽化后出木前期为5.97±1.11 d。雄虫较雌虫提早羽化2-3 d,成虫期需补充营养,且具弱趋光性和假死习性。在日节律中,9 ∶00-11 ∶00和14 ∶00-16 ∶00为产卵高峰期,16 ∶00-19 ∶00为交配高峰期。雌虫出木后2 d开始交配,雄虫出木后4 d开始交配。雌雄虫一生可交配15.12±7.02次,每次交配时长为3.23±0.97 h,一天内仅交配一次。成虫主要趋向长势较弱的木棉树及枝条上产卵。产卵前期为13.50±3.15 d,平均产卵量为117.31±33.25粒。雌雄寿命分别为72.34±15.60 d和46.22±14.38 d。
眉斑并脊天牛;生活史;习性;木棉;天牛科
眉斑并脊天牛GleneacantorFabricius(鞘翅目Coleoptera:天牛科Cerambycidae:沟胫天牛亚科Lamiinae:并脊天牛属Glenea),又称眉斑楔天牛,分布于我国广西、广东、海南、贵州、云南、香港等地(Gressitt,1951;蒲富基,1980;华立中,2002),以及越南、老挝、菲律宾等东南亚国家。该虫寄主种类达7科9属(陆温等,2006),尤其喜食木棉GossampinusmalbaricaDC.(黄华等,2007)。木棉是广西重要的园林绿化树种,近年来该虫对木棉的危害呈上升趋势,导致大批木棉树枯死,严重影响城市园林景观和生态环境。前人曾对其形态特征、人工饲养、发生和防治等开展了相关研究(陆温等,2006;黄华等,2007;赖开平等,2008;李岳诗等,2015),但有关该虫许多重要的生物学特性尚缺乏相关的研究报道。鉴此,本文对该虫的生物学特性进行了系统研究,旨在为有效控制其发生危害提供科学依据。
1 材料与方法
1.1 年生活史的观察
2015年1-2月份从南宁市郊(108.4°E,22.8°N)木棉树上采集受害枝条,剖查越冬虫态。将部分虫枝置于50 cm3×40 cm3×60 cm3饲养箱内,待成虫出木时,将其集中于50 cm3×50 cm3×60 cm3饲养笼内,20对/笼,在室内用新鲜木棉枝叶饲养,每2 d更换一次食料。成虫开始产卵时,放入直径约3.0 cm的木棉枝条供产卵。每天用刀片在刻槽周围(1.0 cm2×1.0 cm2)将卵连同枝条皮层切下,置于直径2.0 cm的孵化管内待孵化。
幼虫孵化后,转接幼虫至直径大于4.0 cm的木棉枝条上,于幼虫培养箱内培养,直至成虫羽化出木。将初羽化成虫集中于饲养笼内继代饲养,观察其年发生代数。在25℃±1℃,相对湿度75%±5%,光周期14 L ∶10 D的条件下观察各虫态发育历期。
1.2 生活习性观察
取同一天出木的越冬代成虫配对于玻璃瓶(直径10.0 cm,高12.0 cm)中饲养,采用维克高清IPC-N82摄像头录像及人为观察并记录其取食、交配和产卵等行为。
将初孵幼虫接入木棉枝条内,置于平底试管(直径1.8 cm,长7.5 cm)内遮光培养,每天剖查幼虫取食及发育等情况。幼虫化蛹后,将蛹移入另一试管内,每日观察蛹的发育过程,直至羽化,共观察60头幼虫。
1.3 成虫昼夜活动节律
取出木15-20 d的健康雌雄虫20对成虫放入50 cm3×50 cm3×80 cm3有机玻璃养虫笼内,用带叶木棉枝条饲养,并放置直径约3.0 cm,长10.0 cm的枝条供成虫产卵。每隔1 h更换一次着卵枝条,并观察记录成虫飞翔、取食、交尾和产卵等情况,连续观察3 d。
1.4 成虫取食选择性的研究
取出木15-20 d的健康雌雄成虫20对放入50 cm3×50 cm3×80 cm3有机玻璃养虫笼内,经1 d饥饿处理后,放入带有叶片、叶柄的木棉枝条,24 h后分别检查叶片、叶柄、叶脉和韧皮部被取食情况,以硫酸纸影印食痕,并换算成取食面积,重复3次。
1.5 成虫耐饥力的研究
在实验种群中取出木15-20 d的健康雌雄成虫各90头,分别称重编号后,按每5头一组放入饲养瓶内(高12 cm,直径10 cm)。实验环境条件同1.1。
取9组雌虫,分别放在饲养瓶内,不提供任何食物,24 h起观察雌虫存活情况,直至雌虫全部死亡;另取9组雌虫,仅在饲养瓶内提供清水(放置一块长2.0 cm,宽1.0 cm的海绵,蘸取适量的蒸馏水),每天更换同样的吸水海绵,24 h起观察雌虫存活情况,直至雌虫都死亡。雄虫测定方法同前。
1.6 数据分析
实验数据用软件SPSS 16.0进行方差分析,成虫对木棉不同营养器官取食选择性和产卵期采用Turkey法进行多重比较,显著水平P≤0.05。
2 结果与分析
2.1 年生活史
眉斑并脊天牛在南宁一年发生4代,有世代重叠现象(表1),完成一个世代的发育需要70 d。雄虫寿命为46.22±14.38 d,雌虫寿命为72.34±15.60 d。第4代一部分发育较早的幼虫可继续发育至成虫并产卵且发育至2-3龄幼虫越冬,但多以第4代4龄幼虫越冬。

2.2 越冬场所及虫态
该虫在南宁主要以老龄幼虫在木棉受害枝条或主干的木质部内越冬,部分中低龄幼虫亦可在主干的韧皮部内越冬,气温回暖时仍可继续取食,少量蛹亦能越冬,越冬虫态与室内饲养的结果基本一致。2月中下旬在蛹室内有成虫羽化,但不出木取食(表2)。在室内温度25℃±1℃,终年均可繁殖,无休眠或滞育现象。
2.3 成虫习性
2.3.1羽化及活动节律
成虫在各个时段均可羽化,其中7 ∶00-19 ∶00占49.53%,19 ∶00-7 ∶00占50.47%。雄虫一般较雌虫提早出来2-3 d。雌雄性比为1 ∶1.12。成虫羽化后在蛹室存在滞留期,滞留期长短取决于环境温度,25℃为5.97±1.11 d,15℃最长可达21 d。成虫出木时,啃咬木质部和树皮呈与树干垂直的椭圆形羽化孔爬出。出木后,可迅速爬行、飞翔至树冠觅食。
成虫具弱趋光性及假死习性,飞翔能力较强,可在高大的木棉树间转株为害。成虫主要在白天活动,5 ∶00-7 ∶00极少活动,雌虫的产卵活动多在9 ∶00-11 ∶00和14 ∶00-16 ∶00进行,交配活动多在16 ∶00-19 ∶00进行。而取食活动在各时段均可进行。
2.3.2取食选择性
成虫需补充营养,且对木棉不同器官的取食具明显的选择性(F=52.94;df=3,11;P<0.000),尤其喜食叶柄(图1),不取食已枯死的树叶及树枝皮层。
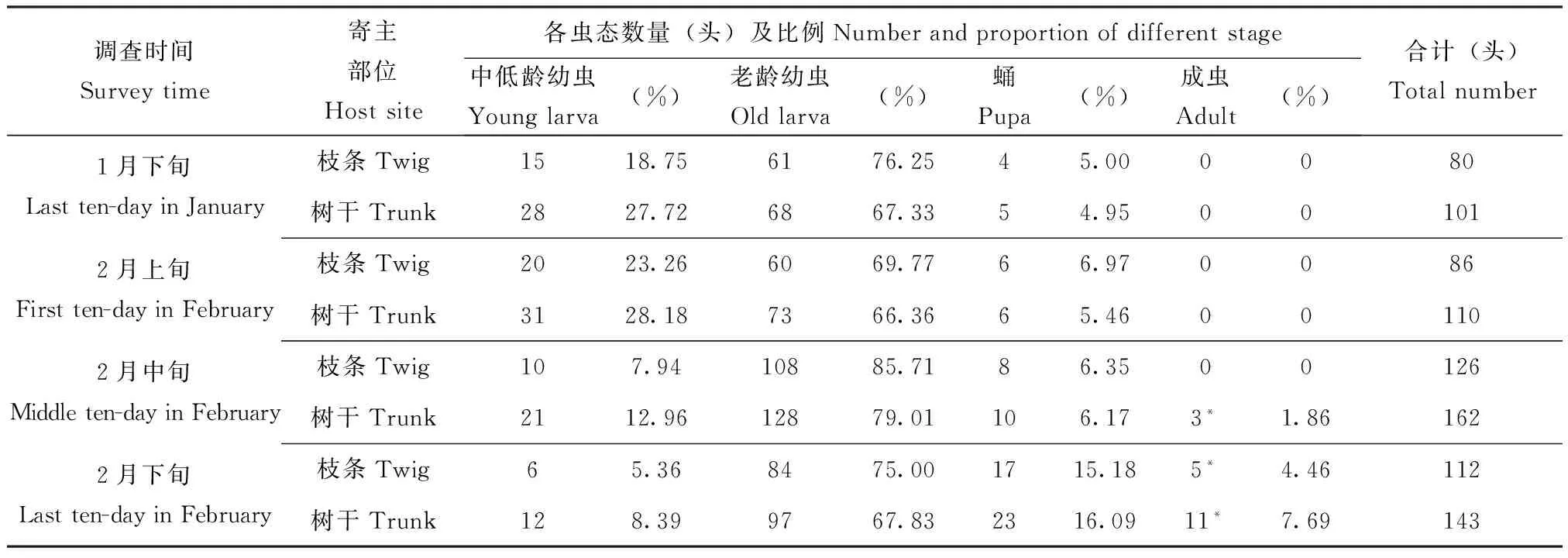
表2 眉斑并脊天牛越冬情况(南宁市,2015年)
注:*, 成虫滞留在蛹室内。Note:*, Adults remained in pupal cells.
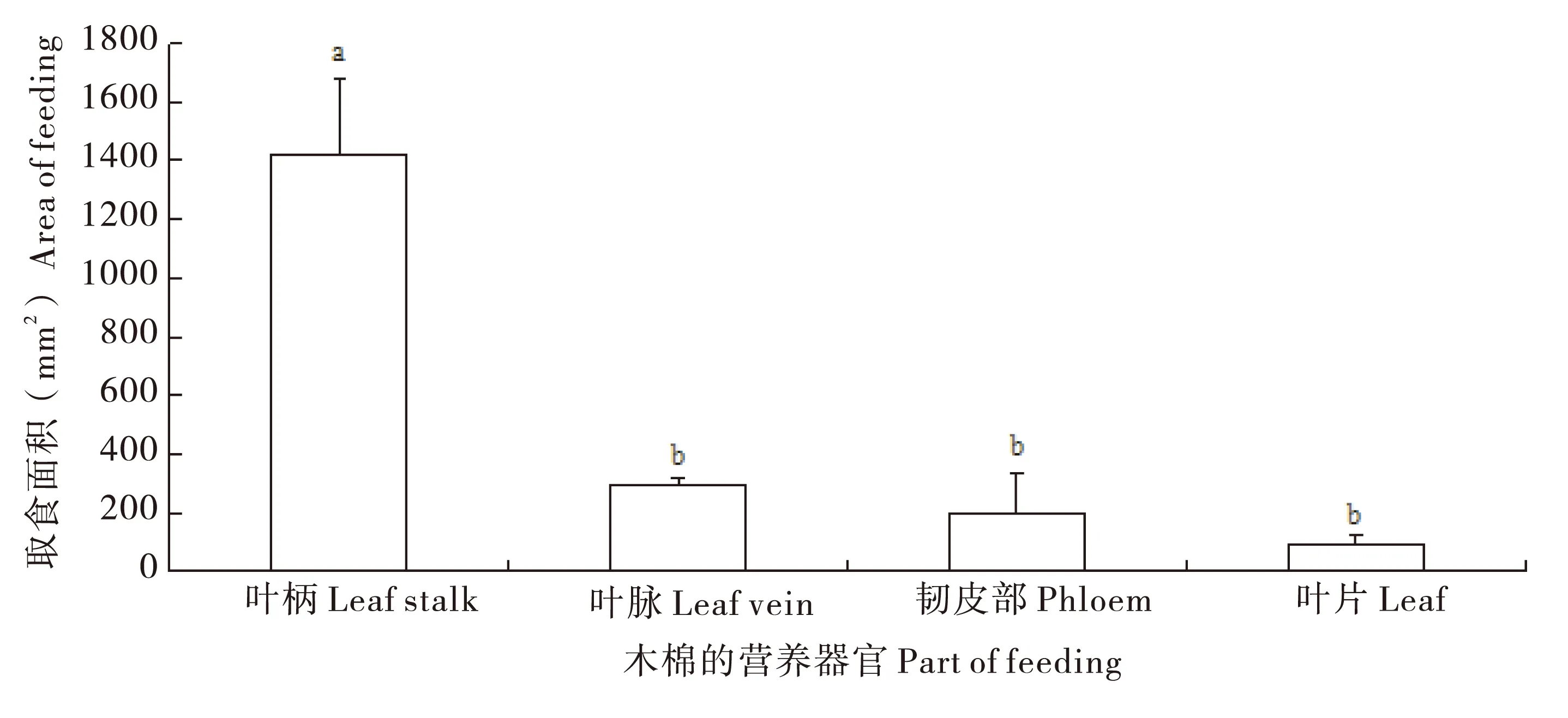
图1 眉斑并脊天牛成虫对木棉各器官的取食选择性Fig.1 Fed preference of adults on different parts of kapok in Glenea cantor Fabricius注:不同字母表示差异性显著(P<0.001)。Note: Values (mean±SE) followed by different letters are significantly different according to the Turkey's test at P<0.001.
2.3.3交配行为
雌虫出木后2 d即可交配,雄虫一般在出木后4 d才开始交配。雄虫较活跃,主动寻找、追逐雌虫,并迅速冲向雌虫,抱合后一般即可立即曲腹交尾。成虫一次交配需3.23±0.97 h。野外交配场所主要在叶片或叶柄。交配初期雌虫剧烈摆动身体,5-7 min后停止摆动。交尾结束后,雄虫主动离开雌虫,少部分雄虫仍保持抱合姿势。雌、雄虫均可多次交配,一生可交配15.12±7.02次。一般雌雄虫一天只交配一次,有短时交配现象,时间为8-20 s不等,通常都因雌虫的强烈拒绝而中断。经饲养观察发现,短时交配所产的卵均不能孵化。
2.3.4产卵与孵化
在实验条件下,雌虫出木后10 d即可产卵,但以出木后12-16 d产卵较多。在野外,雌虫通常选择长势较弱的树木或侧枝上产卵,亦喜趋向于受机械损伤的枝条上产卵。雌虫产卵前在树干或树枝上爬行,寻找合适的产卵部位,用上颚在皮层上咬“一”字形长6.0-10.0 mm的刻槽,刻槽多与枝干垂直,然后掉头将产卵器插入刻槽,将卵产在刻槽中部附近且与虫体同一方向的位置。完成产卵后,腹端将分泌出果冻状的粘稠物涂抹在刻槽上。产一粒卵约需5 min。一般一个刻槽内仅产一粒卵,偶尔多达6粒。雌虫1 d可产卵6-10粒。产卵量为117.31±33.25粒/雌。成虫产卵期为56.45±22.52 d。不同世代间成虫产卵期无显著差异(F=2.23,P>0.05),但越冬代成虫的产卵期略长。25℃±1℃卵历期为5.13±0.61 d,卵孵化率为70%-80%。
2.3.5成虫耐饥力
成虫的耐饥力与性别、有无供水等因素有关。温度25℃±1℃,相对湿度75%±5%,不供水条件下,雄虫一般存活2.47±0.87 d,而在供水条件下,雄虫存活时间明显延长,平均为5.29±1.49 d。雌虫的耐饥能力明显强于雄虫,不供水和供水条件下存活的平均时间分别为4.71±1.22 d和6.93±1.99 d。
2.4 幼虫
幼虫孵化后,即在韧皮部钻蛀取食,近老熟时开始蛀入木质部,先横向钻蛀约1.5-2.0 cm,再向下钻蛀2.0-4.0 cm构筑蛹室,上端的蛀道以木梢堵塞。蛀入孔呈椭圆形,亦被木梢堵塞住。如受害枝条直径在1.5 cm以下,蛹室一般构筑在枝条的髓部。构筑蛹室初期,如蛹室被破坏,幼虫可转移另造蛹室,但不再取食。在25℃下幼虫平均发育历期为41.13±10.89 d。
3 结论与讨论
并脊天牛属是天牛科的一个大属,种类多,分布广,全世界已知1000多种(华立中,2002),我国已知69种。但迄今为止,有关该属种类的生物学方面的报道较少,我国尚未有本属相关种类的生物学的研究报道。本文通过室内饲养,对眉斑并脊天牛的生物学特性进行了系统研究,明确了该虫年生活史、越冬虫态、耐饥能力及生活习性。研究发现该虫的世代历期与Caresche(1937)和Duffy(1968)的报道基本一致。但该虫在南宁地区一年可发生4代,且3-11月均能在野外采集到各虫态,与Duffy(1968)所报道的“该虫的产卵季节为9-10月”不符,其原因可能与饲养条件和该虫所处的地理环境不同有关。
眉斑并脊天牛喜欢集中在衰弱树木或枝条上产卵,而并不在已枯死的树木或枝条上产卵。国外已报道的该属其它种类均可危害活的或衰弱的树木,如GleneagalatheaThomson(Beeson,1920)、GlenealefebureiSorong(Schreurs and Thomas,1961;Duffy,1968)、GleneaaluensisGahan(Duffy,1968)、GleneamultiguttataGuérin-Méneville(Duffy,1968)、GleneanovemguttataGuérin-Méneville(Duffy,1968)和GleneaceliaPasc.(Pang,1979)等。成虫主要出现在5-11月份,基于该虫一次交配即可满足一代卵粒受精的特性(李岳诗,2015),可在3-4月份采用衰弱木作为诱木引诱眉斑并脊天牛产卵而后集中杀灭的物理防治法,避免在城区绿化环境中使用化学防治等对居民生活环境造成污染。此外,野外调查和室内饲养发现花绒坚甲DastarcuslongulusSharp和一种寄蝇(种名待定)是幼虫的寄生性天敌,寄生率达2.1%-3.2%,但对眉斑并脊天牛种群的控制作用尚需进一步研究。
本研究为室内观察结果,尽管这种饲养条件下获得的幼虫大小、蛹重、成虫产卵量等均与野外直接采集的个体无显著差异(韩然勤等,2011),但由于野外环境条件较为复杂,因此对眉斑并脊天牛的发生为害与环境间的关系等仍需进一步研究。
)
Beeson CFC. Insects attacking Yemané [J].IndianForester, 1920, 46 (2): 85-86.
Caresche L. Première note sur lws insects nuisibles au kapokier dans le Sud-Indochinois [J].CRInstituteResearchofAgronomyandForestIndochine, 1937, 2: 175-194.
Duffy EAJ. A Monograph of the Immature Stages of Oriental Timber Beetles (Cerambycidae) [M]. London: British Museum (Natural History), 1968.
Gressitt JL. Longicornia II: Longicorn Beetles of China[M]. Paris: Paul Lechevalier, 1951.
Han RQ, Jia B, Cai JJ,etal. Rearing method forGleneacantorFabricius larvae [J].GuangxiPlantProtection, 2011, 24 (4): 7-9. [韩然勤, 贾豹, 蔡晶晶, 等. 眉斑并脊天牛幼虫的饲养方法[J]. 广西植保, 2011, 24 (4): 7-9]
Huang H, Lu W, Nong C,etal. Studies on artificial diet forGleneacantorFabricius [J].GuangxiPlantProtection, 2007, 20 (1): 9-11. [黄华, 陆温, 农春, 等. 眉斑楔天牛的人工饲料初步研究[J]. 广西植保, 2007, 20 (1): 9-11]
Hua LZ. The Identification Data of Foreign Beetles [M]. Guangzhou: Sun Yat-sen University, 2002. [华立中. 国外天牛鉴定资料[M]. 广州: 中山大学, 2002]
Lai KP, Lu W, Liu DX,etal. The larval instars and stadia of the longhorn beetleGleneacantor[J].ChineseBulletinofEntomology, 2008, 45 (1): 138-140. [赖开平, 陆温, 刘德星, 等. 眉斑并脊天牛幼虫龄数及各龄特征[J]. 昆虫知识, 2008, 45 (1): 138-140]
Lei YM. The damage to kapok trees ofGleneacantorand IPM [J].PlantProtection, 2015, 64: 52-54. [雷艳梅. 眉斑楔天牛对木棉树的为害及综合防治[J]. 植物保护, 2015, 64: 52-54]
Li YS. Studies on Ovarian Development and Sperms Transfer in Female Body inGleneacantor(Fabricius) [D]. Nanning: Guangxi University, 2015. [李岳诗. 眉斑并脊天牛卵巢发育和精子在雌虫体内的转移研究[D]. 南宁: 广西大学, 2015]
Li YS, Li CF, Zhang YJ,etal. Evaluation on effect of a new rearing method forGleneacantorFabricius larvae [J].JournalofSouthernAgriculture, 2015, 46 (8): 1420-1424. [李岳诗, 黎淳锋, 张玉静, 等. 一种新的眉斑并脊天牛幼虫饲养方法效果评价[J]. 南方农业学报, 2015, 46 (8): 1420-1424]
Lu W, Tian MY, Lai KP,etal. The occurrence and prevention ofGleneacantorFabricius [J].GuangxiPlantProtection, 2006, 19 (3): 1-4. [陆温, 田明义, 赖开平, 等. 眉斑楔天牛的发生及防治[J]. 广西植保, 2006, 19 (3): 1-4]
Pang TC, Pan KY. New cocoa stem borerGleneaceliaPasc. (Laminae) in Sabah [J].TechnicalBulletin-Dept.ofAgriculture,Malaysia, 1979.
Pu FJ. Economic Insect Fauna of China Vol.19 Coleoptera Cerambycidae (II) [M]. Beijing: Scince Press, 1980. [蒲富基. 中国经济昆虫志·第十九册·鞘翅目天牛科(二) [M]. 北京: 科学出版社, 1980]
Schreurs J, Simon TRT. Cacao pests in Netherlands New Guinea [J].BulletinsoftheAgriculturalResearchStation,Manokwari(Agricultural Series), 1961, 3: 1-16.
BiologicalcharacteristicsofGleneacantorFabricius(Coleoptera:Cerambycidae)
DONG Zi-Shu1, ZHANG Yu-Jing1, LAI Kai-Ping2, ZHENG Xia-Lin1, WANG Qiao3, LU Wen1*
(1. College of Agriculture, Guangxi University, Nanning 530004, China; 2. Guangxi Research Institute of Chemical Industry, Nanning 530001, China; 3. Institute of Natural Resources, Massey University, Palmerston North 11122, New Zealand)
GleneacantorFabricius is an important stem borer ofGossampinusmalbaricaDC. in southern China. Biological characteristics of this species was studied through indoor rearing and field investigation. Results showed thatG.cantorhad 4 generations a year in Nanning City, Guangxi Zhuang Autonomous Region, and mean generation duration was 70 d.G.cantorusually overwintered as the fourth instar larvae in host trunks or branches. Adults stayed in the pupal cells after eclosion for 5.97±1.11 d. Emergence of males was earlier than females for 2-3 d. Both males and females displayed the behaviors of complementary nutrition, weak phototaxis, and suspended animation. The diel oviposition was observed from 9 ∶00 to 11 ∶00 a.m and from 14 ∶00 to 16 ∶00 p.m, and the diel mating between 16 ∶00 and 19 ∶00 p.m. Mating behavior of females was observed after emergence for 2 d, but that of males for 4 d. Mating times was 15.12±7.02 times during their life span. Both sexes ofG.cantormate only once a day, and each mating duration was 3.23±0.97 h. Females preferred the stems and branches of weak trees to oviposit. The preoviposition period was 13.50±3.15 d. Oviposition number of per female was 117.31±33.25. The average longevity of both female and male was 72.34±15.60 d and 46.22±14.38 d.
GleneacantorFabricius; life history; habits; kapok; Cerambycidae
董子舒,张玉静,赖开平,等.眉斑并脊天牛生物学特性研究[J].环境昆虫学报,2017,39(6):1313-1318.
Q964;S433.5
A
1674-0858(2017)06-1313-06
国家自然科学基金(31260432,31660626)
董子舒,男,1992年生,在读硕士研究生,研究方向为昆虫行为生态,E-mail:dongzishu1668@163.com
*通讯作者Author for correspondence, E-mail: luwenlwen@163.com
Received: 2016-09-01; 接受日期Accepted: 2016-11-03
猜你喜欢
热带生物学报(2022年6期)2022-11-29 14:14:44
甘肃农业科技(2021年6期)2021-07-25 16:11:05
歌海(2021年6期)2021-02-01 11:27:18
幼儿园(2020年15期)2020-11-24 13:00:20
环境昆虫学报(2020年5期)2020-11-11 08:51:06
艺术家(2020年5期)2020-07-09 09:49:18
科学导报(2019年35期)2019-09-03 04:35:11
海峡姐妹(2019年5期)2019-06-18 10:40:40
西北农林科技大学学报(自然科学版)(2018年5期)2018-05-24 12:33:34
百科知识(2016年13期)2016-08-05 18:23:30
