
模拟降水和氮沉降对准噶尔盆地南缘梭梭光合生理的影响*
2018-01-04 01:43赵文勤谢双全吕新华贺亚玲
中国生态农业学报(中英文) 2018年1期
张 维, 赵文勤, 谢双全, 张 莉, 吕新华, 贺亚玲
模拟降水和氮沉降对准噶尔盆地南缘梭梭光合生理的影响*
张 维, 赵文勤**, 谢双全, 张 莉, 吕新华, 贺亚玲
(石河子大学生命科学学院/石河子大学化工学院 石河子 832000)
氮沉降和降水格局变化是目前全球气候变化背景备受关注的热点研究课题, 也是荒漠生态系统的两个主要限制因子。因此, 研究两者对荒漠植物的效应有助于深入了解荒漠生态系统对全球变化的响应。本文选择准噶尔盆地南缘荒漠地区的建群种梭梭()作为研究对象, 设置自然降水(W0)与增加降水30%(W1)两个水分条件和自然氮沉降(N0)、增加氮素30 kg(N)·hm-2·a-1(N1)与增加氮素60 kg(N)·hm-2·a-1(N2) 3个施氮水平, 连续处理2年, 以探究降水、氮沉降及其交互作用对梭梭光合日变化及生理生态特征的影响。结果表明: 降水、氮沉降及其交互作用对梭梭的净光合速率(n)日变化产生极显著正相关影响; 同时根据梭梭n、胞间CO2浓度(i)及气孔限制值(s)的变化方向, 推测梭梭光合“午休”主要由非气孔因素引起。此外, W0条件下, 梭梭丙二醛(MDA)含量、抗氧化酶(POD、CAT、SOD)活性、可溶性蛋白(Pr)和可溶性糖(SS)含量均随施氮量增加而显著降低, 脯氨酸(Pro)含量则呈先降低后增加的趋势; 而W1条件下, 梭梭MDA含量、抗氧化酶(POD、CAT、SOD)活性及渗透调节物质(Pro、Pr、SS)含量均随施氮量增加呈显著先增加后降低的趋势。两种水分条件下, 除W1N1处理梭梭的Pro含量高于对照组外, 其余处理均显著低于对照组; 同时梭梭的MDA含量、抗氧化酶活性、Pr及SS含量也均显著低于对照组。综合分析结果表明: 降水、增氮及其交互作用均有利于梭梭的生长, 但其交互作用效应的强弱则取决于二者间的比例。
梭梭; 降水; 氮沉降; 水氮交互; 光合生理特征; 准噶尔盆地南缘
植物叶片是进行光合作用的主要器官, 也是对外界环境变化最敏感的指示器[1]。光合作用是植物最重要的生理生化活动, 为植物提供有机营养物质和能量, 是一切生命活动赖以生存的基础, 并受养分与水热条件等环境因子的影响。其中, 水分和养分是影响植物光合作用的主要生态因子[2]; 是植物生长发育所必需的资源, 其有效性对植物叶片的光合生理功能具有重要影响; 也是限制植物生长和生产力的两个最重要因素, 且研究还证明二者之间存在显著的耦合效应[3-4]。
然而近代以来, 由于工业的快速发展和人类活动干扰的加剧, 导致陆地生态系统大气氮沉降现象显著增加, 造成不同程度的土壤酸化和植被退化[5-6]。据IPCC[7]预测: 未来中纬度地区降水将会增加, 对此我国西北干旱区降水也可能出现增加的趋势[8]; 而大气氮沉降效应也会伴随着全球降水格局的变化而发生变化。准噶尔盆地地处亚欧大陆干旱半干旱气候带, 水资源缺乏, 生态环境极其脆弱, 对全球气候变化的响应十分敏感[9], 其最明显的特点是年降水时空分布不均, 蒸发量大, 土壤贫瘠且养分有效利用率低[10]。该区的优势种及建群种梭梭()是分布最广、面积最大的荒漠植被类型之一, 为多年生灌木或小乔木, 古地中海区系遗留物种, 现被列为国家濒危三级保护植物。尽管目前国内外有关梭梭光合生理方面的研究较多, 但大多集中于单因素水平, 如: Gu等[11]研究结果表明, 梭梭的气孔敏感性随土壤水分亏缺呈线性增长, 同时导致非等氢离子气孔行为向等氢离子气孔行为转变; Kang等[12]研究结果表明, 梭梭在干旱环境下能够积累高浓度的Ca2+, 并将其直接用于渗透调节; Zhang等[13]研究表明梭梭根的保护策略是其存活的关键。而对于野外梭梭原生境下的多因素及其交互作用方面的研究还较少。因此, 本研究在尽量不破坏梭梭原生境的前提下, 对其光合生理生态学特征进行野外人工模拟降水、氮沉降及其交互作用的研究, 旨在探讨该地区梭梭种群在全球气候变化下的适应策略及其光合生理调控机制, 为该区荒漠生态系统的恢复重建及科学合理的管理提供理论指导和参考。
1 材料与方法
1.1 研究区概况
研究区位于新疆阜康县的中国科学院新疆生态与地理研究所的阜康荒漠生态研究站(87°45′E, 44°30′N, 海拔436 m), 地处天山北麓、准噶尔盆地南缘。该区年平均气温6.1 ℃, 最高气温41.1 ℃, 最低气温-34.4 ℃; 年平均降雨量117.2 mm, 年平均蒸发量1 942.1 mm, 属温带大陆性干旱荒漠气候。植被属典型荒漠灌木、半灌木及半乔木的单生或混生稀疏植被, 而梭梭是该区域主要的防风固沙稀疏植被类型之一。
1.2 试验设计
试验于2014年年初在研究站实验基地的围栏沙地内进行, 该区梭梭的平均生长密度约600株·hm-2, 郁闭度0.5~0.7, 平均株高180 cm; 选取地势平坦、梭梭分布相对均一且树龄及长势相当的地段作为本研究样区。之后在该样区内设置10 m × 10 m的小样方, 且每个样方内梭梭数至少在7株左右的对照组(W0N0)、降水处理组(W1N0)、氮沉降处理组(W0N1和W0N2)及水氮交互处理组(W1N1和W1N2)各4个重复。降水处理分为自然降水(W0)及在自然降水基础上增加降水30%(W1)(增加值为阜康过去30年年降水量平均值200 mm的30%, 即共增加降水60 mm)两个水平, 对增加的30%降水又均分到春、夏、秋3个季节进行, 即每个季节分别增加20 mm。其中春季增加降水于每年的3月25日开始, 夏季增加降水于每年的6月20日开始, 秋季增加降水则于每年的8月20日开始, 均为每周增加一次, 每次增加5 mm, 共进行4周; 且处理时间均为6:00—8:00。为保证降水处理的顺利完成和更好地模拟自然降水, 结合小区面积和模拟降水量准确计算出每次增水量, 提前一天用水车将水运送到样地, 将其存储在样地旁边的专用水箱里, 采用喷灌装置定量将其以淋洗的方式撒入样地。氮沉降处理则分为自然氮沉降(N0)、自然氮沉降基础上增加氮素30 kg(N)·hm-2·a-1(N1)和自然氮沉降基础上增加氮素60 kg(N)·hm-2·a-1(N2)3个水平, 并结合该区氮沉降的主要沉降形式, 使增加的氮素均以硝酸铵(NH4NO3)的形式添加。此外为了使植物更好地吸收和模拟氮沉降, 将NH4NO3溶于100 L水中, 同时为尽量避免NH4NO3的流失而采用喷雾器均匀地将其喷入样方中, 对照则喷入等量的水, 且也分为春、夏、秋3个季节进行, 增加日期与降水处理同日进行。
1.3 气体交换参数的测定
气体交换参数测定于2015年梭梭生长旺季(7月)的晴天进行, 利用便携式光合测定系统仪(Li-6400XT, LI-COR, USA)随机选取各处理样方中生长良好的梭梭植株进行测定。每次在每一植株上随机选取3簇(3个重复)部位相同长势相近的梭梭同化枝进行测定, 共测取15组数值, 最后取其平均值进行统计分析。测定时间为8:00—20:00, 期间每隔2 h测定1次。其各项参数数值均由光合仪系统自动同步记录, 主要包括: 梭梭同化枝净光合速率(n)、胞间CO2浓度(i)。而其同化枝瞬时水分利用效率(WUE)与气孔限制值(s)则通过以下公式计算得出: WUE=n/r[14];s=1−i/a[15](r为蒸腾速率,a为大气CO2浓度)。此外利用土钻钻取各处理样方内相同方位上60 cm深的混合土盛于铝盒中称其湿重, 后将其带回实验室进行物化特征的测定。
1.4 材料获取及各项生理指标的测定
在测定完气体交换参数后, 将所用梭梭同化枝嫩尖去掉盛于离心管中, 并立即将其保存于液氮罐中, 然后带回实验室进行各项生理指标的测定, 主要包括: 丙二醛(MDA)含量、过氧化物酶(POD)、过氧化氢酶(CAT)和超氧化物歧化酶(SOD)活性以及脯氨酸(Pro)、可溶性蛋白(Pr)和可溶性糖(SS)含量。其中, MDA含量的测定采用硫代巴比妥酸法, POD活性的测定采用愈创木酚法, CAT活性的测定采用碘量法, SOD活性的测定采用氮蓝四唑(NBT)光化学还原法[16], Pro含量的测定采用磺基水杨酸法, Pr含量的测定采用考马斯亮蓝法, SS含量的测定采用蒽酮比色法[17]。用G-250型分光光度计比色测定OD值。每组处理测取4个数值, 最后取其平均值进行统计分析。
1.5 数据分析
利用双因素方差分析(two-way ANOVA)分析各处理对梭梭生境土壤理化特征[土壤含水率(WCS)、电导率(EC)、pH、有机质(SOM)、全氮(TN)、全磷(TP)、全钾(TK)]及梭梭生理指标(MDA、POD、SOD、CAT、Pro、Pr、SS)的效应, 用单因素方差分析对各处理间的以上指标及梭梭n、i、s和WUE进行分析, 用LSD(least significant difference)法进行多重比较(<0.01和<0.05)。统计分析利用SPSS 17.0完成, 用Origin 8.0软件作图。
2 结果与分析
2.1 梭梭生境土壤理化性质对降雨和氮沉降增加的响应
方差分析结果表明, 施氮及水分处理极显著地影响了梭梭生境WCS、EC、SOM、TN及TK含量, 对土壤pH影响显著, 而降水对土壤TP含量则影响不显著(表1)。
在W0条件下, 梭梭生境WCS、EC、SOM、TN及TP含量均随施氮量的增加而极显著增加; 而pH和TK含量则随施氮量的增加呈先增后降趋势, 同时除N0和N2之间的pH差异性不显著外, 其余各处理间差异极显著。但在W1条件下, 梭梭生境土壤理化特性则表现为土壤SOM、TN及TP含量均随施氮量的增加而极显著增加, 而WCS和EC则均随施氮量的增加呈先增后降趋势, 且组间差异性极显著; TK含量的变化情况则与W0条件下相同(表2)。
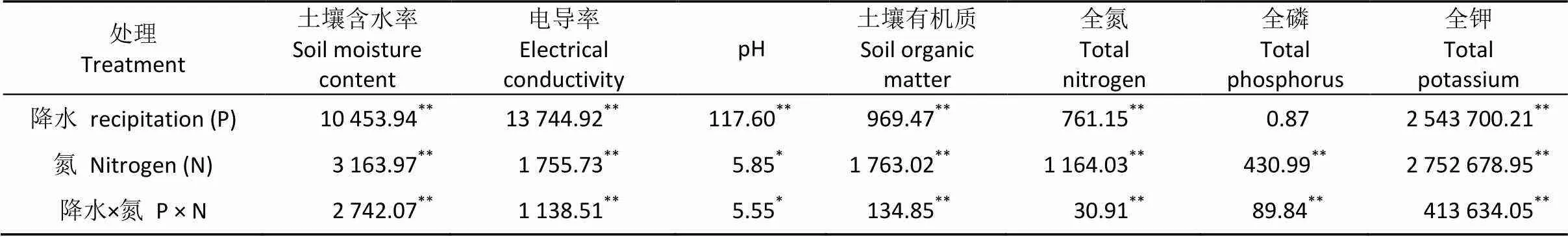
表1 水氮处理对梭梭土壤基质理化特征影响的双因素方差分析
表中数值为检验值。Values are results oftest. *< 0.05; **< 0.01.
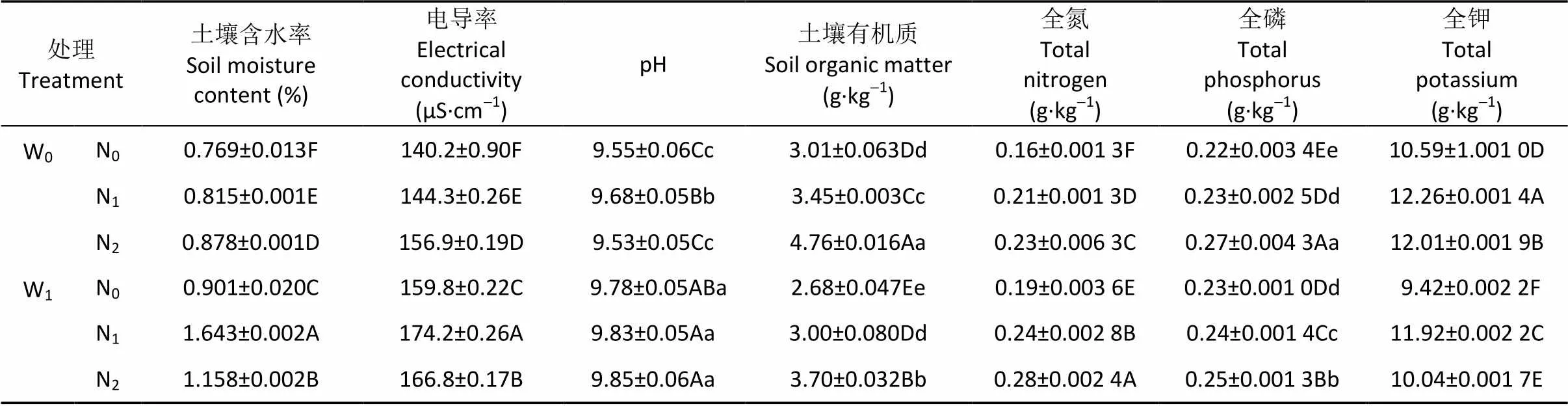
表2 不同水氮处理下梭梭样地土壤的理化特性
W0: 自然降水; W1: 自然降水基础上增加30%; N0: 自然氮沉降; N1: 自然氮沉降基础上增加30 kg(N)·hm-2·a-1; N2: 自然氮沉降基础上增加60 kg(N)·hm-2·a-1。同列不同大、小写字母分别表示在<0.01和<0.05水平上差异显著。W0: natural precipitation; W1: natural precipitation increases by 30%; N0: natural nitrogen deposition; N1: natural nitrogen deposition increases by 30 kg(N)·hm-2·a-1; N2: natural nitrogen deposition increases by 60 kg(N)·hm-2·a-1. Different capital and lowercase letters within the same column indicate significant differences at the levels of< 0.01 and< 0.05, respectively.
2.2 梭梭气体交换参数对不同处理的响应
方差分析表明, 施氮和降水处理均能够极显著地影响梭梭同化枝气体交换。W0条件下, 各氮素水平下梭梭n在8:00、18:00和20:00均随氮素增加而呈极显著地先增加后降低趋势, 其余测定时刻(除14:00的N0和N1外)则均随氮素增加而呈极显著增加。W1条件下, 各氮素水平下梭梭n在8:00和20:00的变化趋势与W0条件下相同, 其余测定时刻则均随氮素增加而呈极显著呈先降低后增加趋势; 但不同处理下梭梭n的整个日变化却均呈典型的“双峰”型趋势, 且出现峰值与峰谷值的时刻也均一致(图1a)。
叶片i受多种因素(环境及自身因素)的影响, 进而最终影响到植物的光合作用。W0条件下, 各氮素水平下梭梭i除12:00外, 其余测定时刻则均随氮素增加而呈极显著降低趋势, 整个日变化过程也均呈“双峰”型, 且出现峰值的时刻也均相同, 即8:00和16:00; 同时其峰谷值也均出现在10:00。W1条件下, 各氮素水平下梭梭i除8:00和20:00随着氮素增加而呈极显著先降低后增加趋势外, 其余各时刻则均随氮素增加呈极显著先增加后降低趋势, 且整个日变化过程同样均呈“双峰”型趋势, 峰值与峰谷值出现的时刻也均与W0条件下相一致(图1b)。
s值大小反映因气孔导度下降, 导致CO2进入气孔的减少程度。经统计分析表明, 梭梭s的日变化趋势在不同处理下存在着极显著差异。W0条件下, 各氮素水平下梭梭s除12:00随着氮素增加而呈极显著先增加后降低趋势变化外, 其余时刻则均随氮素增加而极显著增加。但W1条件下, 各氮素水平下梭梭s在8:00随着氮素增加而降低, 20:00则随着氮素增加而呈极显著先增加后降低趋势, 其余时刻随氮素增加的变化情况均与20:00时相反。各处理下梭梭s的整个日变化趋势均呈“双峰”型。其中第1个峰值均出现于10:00, 第2个峰值除W0N0、W1N0和W1N2出现于18:00外, 其余各处理则均出现于20:00; 两水分条件下的峰谷值分别出现在14:00和16:00(图1c)。
WUE反映植物耗水与干物质生产之间的关系, 是评价植物生长适宜程度的综合生理生态指标[18]。不同水分条件下梭梭WUE随氮素逐渐增加的变化趋势除14:00外, 其余时刻均一致, 且不同处理组的整个日变化过程也均呈明显的“双峰”型趋势。其中, 第1个峰值除W0N1出现于8:00外, 其余处理组均出现于10:00, 而峰谷值则均出现于14:00; 第2个峰值则均出现于18:00(图1d)。
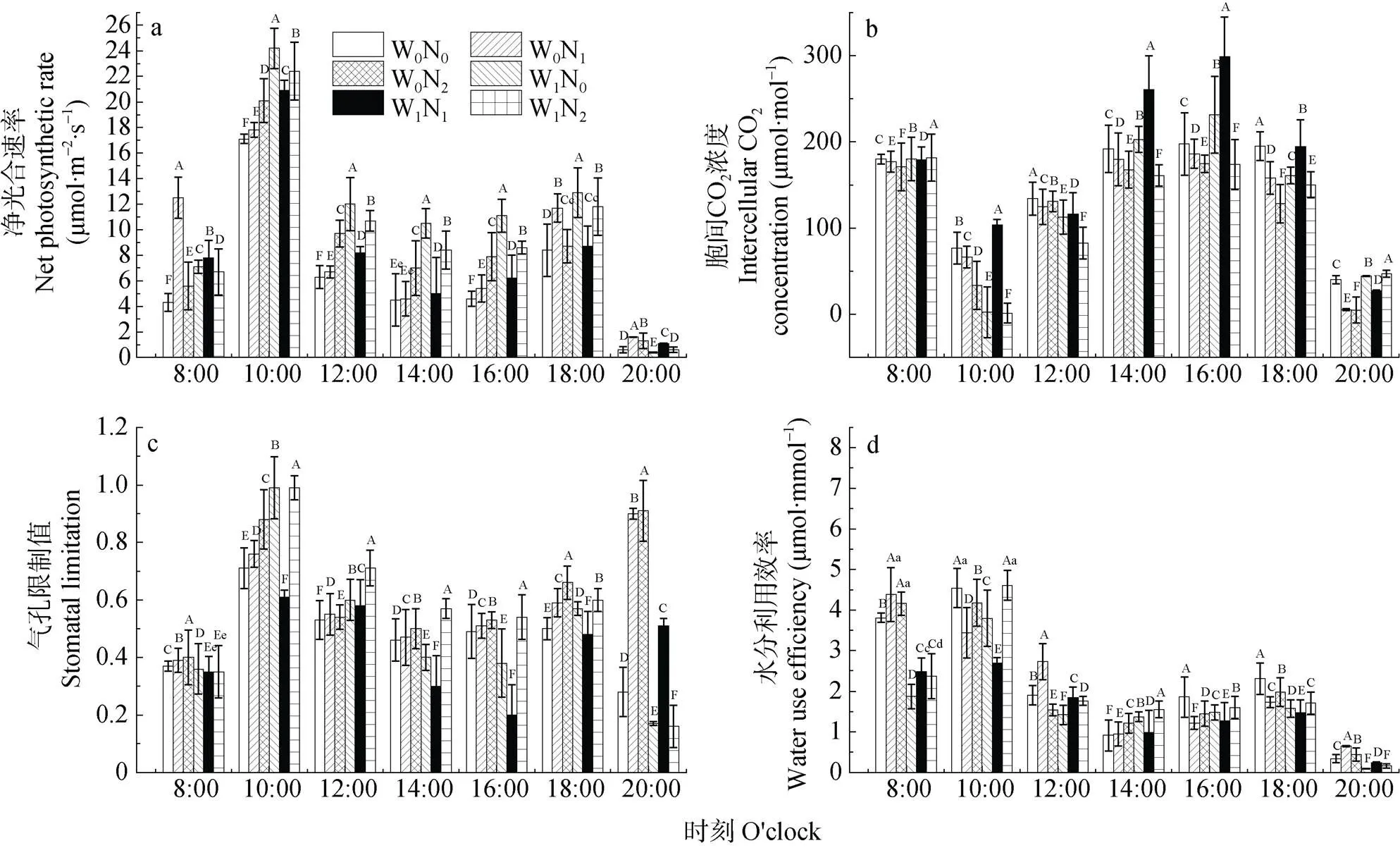
图1 不同水氮处理下梭梭气体交换日变化
W0: 自然降水; W1: 自然降水基础上增加30%; N0: 自然氮沉降; N1: 自然氮沉降基础上增加30 kg(N)·hm-2·a-1; N2: 自然氮沉降基础上增加60 kg(N)·hm-2·a-1。同时刻不同大、小写字母分别表示处理间差异极显著(<0.01)和显著(<0.05)。W0: natural precipitation; W1: natural precipitation increases by 30%; N0: natural nitrogen deposition; N1: natural nitrogen deposition increases by 30 kg(N)·hm-2·a-1; N2: natural nitrogen deposition increases by 60 kg(N)·hm-2·a-1. Different capital and lowercase letters at the same time indicate significances at< 0.01 and< 0.05 among treatments, respectively.
2.3 梭梭MDA含量、抗氧化酶活性及渗透调节物质含量在不同处理下的响应
方差分析结果表明, 施氮和降水处理均能够极显著地影响梭梭同化枝MDA、Pro、Pr和SS含量以及POD、SOD和CAT活性(表3)。
通常MDA被作为衡量植物细胞受逆境胁迫伤害程度的重要生理指标。在W0条件下, 梭梭同化枝MDA含量随施氮量增加而显著降低; 而W1条件下则随施氮量增加呈极显著先增加后降低趋势, 但均极显著高于N0水平下梭梭MDA含量, 分别是N0水平下的2.3倍和1.4倍。无论何种处理梭梭MDA含量均极显著地低于对照组(图2a)。
在抗氧化酶系统中, POD和SOD是植物体内清除及减少活性氧积累的重要保护酶, 同时前者还能抵御膜脂过氧化和维持膜结构完整性, 后者则进一步清除潜在过氧化氢对植物的伤害。而CAT也对维护植物细胞氧化自由基代谢、水分代谢平衡及光合作用有着极其重要的作用。在W0和W1条件下, 梭梭同化枝POD、CAT及SOD活性随施氮量增加的变化情况均一致, 即W0条件下均随施氮量增加而显著降低, W1条件下则呈先增加后降低的变化趋势, 然而无论何种处理, 其梭梭同化枝的各种酶活性均低于对照组(图2b, 2c, 2d)。
除保护酶系统外, 渗透调节也是植物抵御各种逆境胁迫伤害的重要生理机制之一, 其中脯氨酸、可溶性蛋白及可溶性糖是最常见最重要的渗透调节物质。在W0条件下, 梭梭同化枝脯氨酸含量随施氮量增加呈极显著先降低后增加的变化趋势, 但N2水平下的脯氨酸含量仍极显著低于N0水平; 而W1条件下则随施氮量增加呈极显著先增加后降低趋势, 且N2水平下的脯氨酸含量是对照组的1.4倍(图2e)。相对脯氨酸而言, 梭梭同化枝的可溶性蛋白及可溶性糖含量的变化情况则表现出: W0条件下两者均随施氮量增加而呈极显著降低趋势, 而W1条件下则呈先增加后降低趋势, 且W1N1水平下的含量均显著高于W1N2水平而极显著低于对照组(图2f, 2g)。
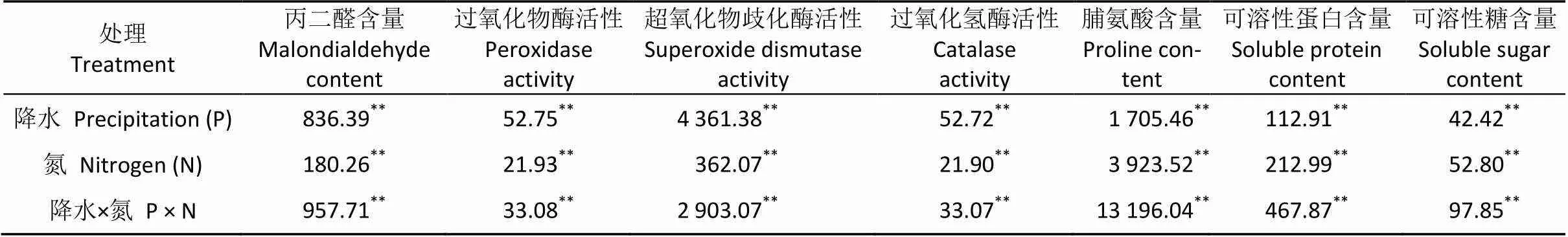
表3 水氮处理对梭梭生理影响的双因素方差分析
表中数值为检验值。Values are results oftest. **< 0.01.
3 讨论
土壤的理化性质不仅能反映土壤结构、营养状况及蓄水能力, 还是影响植物生长发育及生物量的基本要素, 受植被类型、土壤形成机制、生态系统及人类活动的影响[19]。氮沉降及降水格局变化是当前全球气候变化的主要特征之一, 能直接影响土壤含水率、pH、电导率以及养分状况, 且不同处理对其产生的结果存在很大差异[20]。本研究中, 在干旱的W0条件下, 梭梭生境土壤SCM、EC、SOM、TN及TP含量均随施氮量增加而呈极显著性增加趋势, 而pH和TK含量则随施氮量增加呈极显著先增加后降低趋势变化, 说明在干旱胁迫下, 氮的增加有利于提高土壤的保水能力及改善植物根际的营养条件, 促进植物生长; 同时也能在一定氮水平下促进植物对钾素的吸收[21], 其原因可能与植物的生长状况及氮钾之间的交互作用相关[22]; 但过量施氮会导致土壤pH下降[23]。而在湿润的W1条件下, 除土壤SOM、TN、TP及TK含量的变化情况与W0条件下相同外, 其余各项土壤成分指标均表现出明显差异。其中SCM和EC均随施氮量的增加呈极显著先增加后降低趋势变化, 而土壤pH则随施氮量的增加呈极显著性增加, 其主要原因可能在于降水处理打破了土壤原营养元素间的平衡, 同时氮含量增加又进一步导致土壤中微生物活性的提高, 从而加速了土壤中无机元素的转变与有机质等的形成[24]。
植物叶片是进行光合作用的主要器官[1], 也是对环境变化较为敏感的营养器官, 其生理生化特征的变化均能体现环境因子对植物的影响程度或植物对环境的适应状况[25]。而水分与氮素不仅是植物生长发育的重要环境因子, 也是植物生物量形成的重要组成元素[20], 因此, 水分与氮素的变化必定对植物产生一定的影响。大量研究表明[26], 降水、增氮均能明显提高植物n, 而其交互作用的效应则取决于两者间的比例。在本研究中, 各处理下梭梭n的整体日变化趋势均呈典型的“双峰”型, 即所谓的“午休”现象。对此, 我们根据植物n降低时, 若i降低而s升高, 则表明气孔导度降低是导致其n降低的主要原因; 反之, 则表明主要原因为非气孔因素这一判断依据, 证明了梭梭“午休”现象主要是受非气孔性限制的影响, 与田媛等[27]对梭梭的研究结果不同。其次, 刘晓宏等[28]对春小麦(L.)的研究结果表明氮的增加可提高作物水分利用效率; 而王海江等[29]在对棉花(spp.)的研究中则得到水分利用效率随灌溉水量、施氮量的增加呈先增后降趋势。本研究结果与前人报道相同。此外, 本研究结果还表明梭梭WUE随施氮量的增加呈先降低后增加的趋势变化。而出现这一系列结果的主要原因可能与测定时大气温度差异所引起的土壤水分蒸发程度不同相关。证实了在干旱条件下, 氮的增加能在一定程度上起到补偿水分亏缺对植物所导致的胁迫伤害这一结论, 与柴仲平等[30]对红枣(L.)的研究结果类似; 进一步表明了不同水氮处理对植物水分利用效率所起作用的差异性, 也与李银坤等[31]的研究结果相同。
在正常生长环境条件下植物体内活性氧的产生与清除处于一种动态平衡状态[32], 仅在受到胁迫伤害时才会迫使原有的平衡被打破, 进而导致植物体内活性氧自由基积累并引发细胞膜脂过氧化反应, 而MDA即其氧化的终产物, 其积累量可表示细胞膜脂过氧化的程度[33]。本研究中, 在干旱的W0条件下, 梭梭MDA含量随施氮量增加而显著降低; 但在湿润的W1条件下, 则随施氮量增加呈先增加后降低的趋势变化。说明在干旱胁迫下, 降水与施氮均有利于缓解植物因水分亏缺所造成的伤害, 但二者交互作用时的效应则应考虑其具体的比例。此外, 植物体为清除过剩活性氧及减轻胁迫对细胞的伤害, 也会增强自身体内的保护酶活性。然大量研究表明, 胁迫条件下, 适量施氮能显著降低植物体内的POD、CAT及SOD活性以及显著降低其MDA含量[34]; 但施氮量过多则会加剧植株细胞膜的氧化反应, 从而降低植株对活性氧的清除能力[34]。在本研究中, POD、CAT及SOD活性在不同水分条件下随施氮量增加的变化趋势均与MDA含量的变化一致。即均在干旱的W0条件下随施氮量增加而显著降低; 但在湿润的W1条件下则随施氮量增加呈显著先增加后降低的趋势变化, 这与Li等[35]的研究结果不同。其主要原因可能与不同的植物或逆境条件下, 植物保护酶活性对逆境的响应特征不同有关。植物体内的脯氨酸与可溶性糖通常被认为是渗透调节的主要物质, 其含量的变化与植物的抗逆性相关[36]。至于可溶性蛋白由于其成分多为酶蛋白, 则多被认为与代谢水平相关[37]; 但其含量的增加也有利于提高植物的抗胁迫能力与适应性[38]。如杨晓清等[39]对土沉香()的研究表明, 在同一水分条件下其幼苗脯氨酸含量随施氮量的增加而显著增加, 这与本研究结果有一定差异, 其主要原因可能与不同物种间对环境因子的响应机理不同有关。此外王景燕等[38]对汉源花椒()的研究表明, 幼苗可溶性糖与可溶性蛋白含量均随土壤含水量的增加呈先增后降趋势变化, 均随施肥量的增加而增加。而本研究中, 梭梭可溶性糖与可溶性蛋白含量在干旱的W0条件下均随施氮量的增加而呈现极显著降低; 而在湿润的W1条件下则均随施氮量的增加呈现先增加后降低的变化趋势, 与王景燕等[38]的研究结果不同。其主要原因可能在于可溶性糖不仅是逆境胁迫下的渗透调节物质, 同时也是植物光合作用产物[38], 而可溶性蛋白则受胁迫下植物代谢水平的影响。
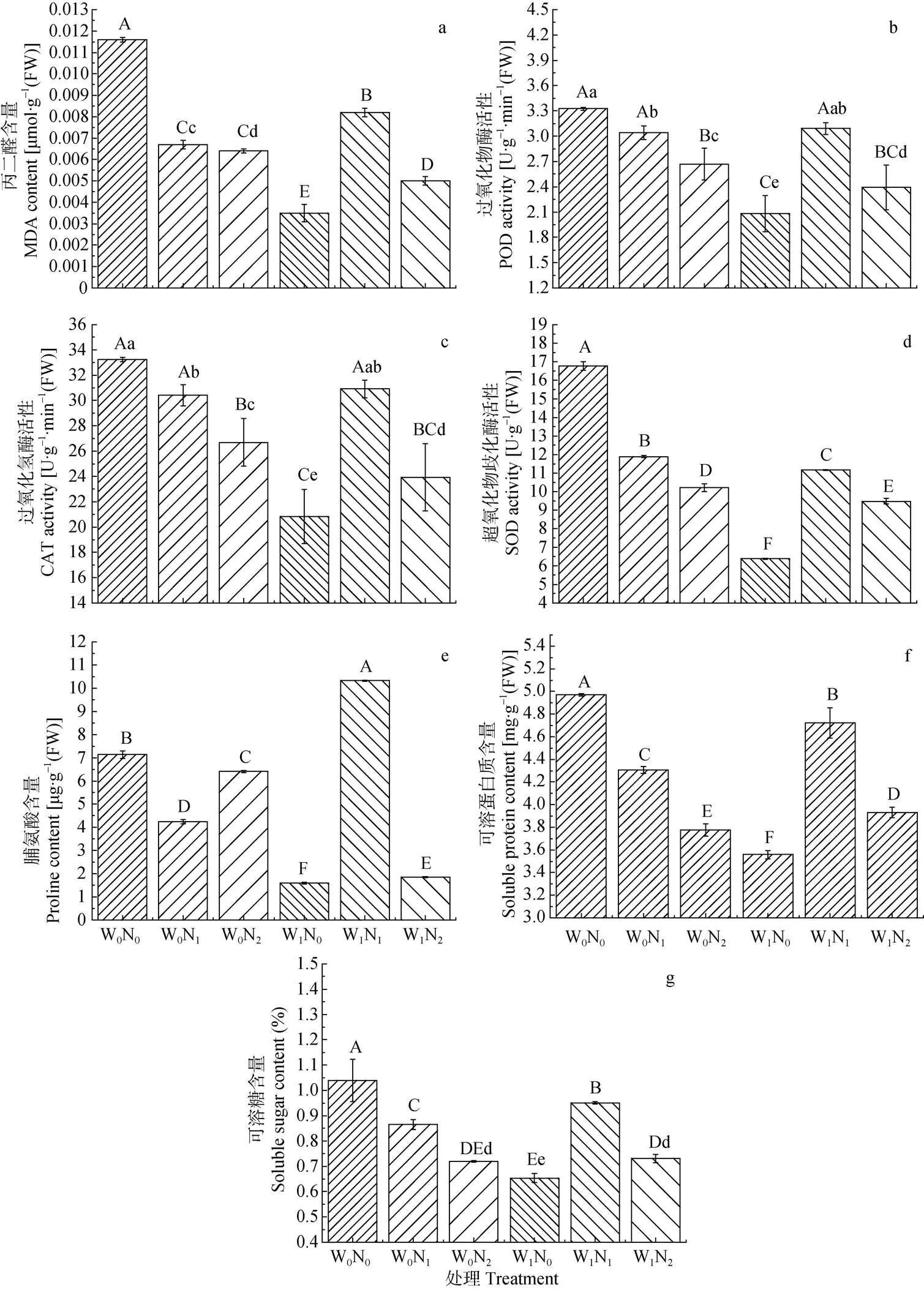
图2 不同水氮处理下梭梭同化枝丙二醛含量及过氧化物酶、过氧化氢酶和超氧化物歧化酶活性以及脯氨酸、可溶性蛋白和可溶性糖含量
W0: 自然降水; W1: 自然降水基础上增加30%; N0: 自然氮沉降; N1: 自然氮沉降基础上增加30 kg(N)·hm-2·a-1; N2: 自然氮沉降基础上增加60 kg(N)·hm-2·a-1。不同大、小写字母分别表示处理间差异极显著(<0.01)和显著(<0.05)。W0: natural precipitation; W1: natural precipitation increases by 30%; N0: natural nitrogen deposition; N1: natural nitrogen deposition increases by 30 kg(N)·hm-2·a-1; N2: natural nitrogen deposition increases by 60 kg(N)·hm-2·a-1. Different capital and lowercase letters indicate significances at< 0.01 and< 0.05 among treatments, respectively.
4 结论
降水与施氮对梭梭生境土壤的理化特性、营养状况及梭梭光合生理特征的影响显著。干旱条件下, 氮素的添加能极显著地改善梭梭根际的营养状况及在一定程度上缓解干旱对其的影响; 同时还可相对提高植物体内主要的保护酶(POD、CAT、SOD)活性及渗透调节物质(Pro、SS、Pr)的含量, 使MDA含量维持在相对较低水平, 促进其光合速率、水分利用效率的增加, 从而促进植物生长。而湿润条件下, 氮素添加对植物抗逆性所起的作用则取决于水氮交互作用时的比例。而本研究中, 增加施氮30 kg(N)·hm-2·a-1与60 kg(N)·hm-2·a-1对梭梭抗逆性所起的作用均较自然氮沉降显著增强, 说明在未来全球气候变化下单就降水与氮沉降增加而言, 将有利于准噶尔盆地南缘地区梭梭的生长。
[1] FU G, SHEN Z X. Response of alpine soils to nitrogen addition on the Tibetan plateau: A meta-analysis[J]. Applied Soil Ecology, 2017, 114: 99–104
[2] ZHANG X L, TAN Y L, LI A, et al. Water and nitrogen availability co-control ecosystem CO2exchange in a semiarid temperate steppe[J]. Scientific Reports, 2015, 5: 15549
[3] YIN C Y, PANG X Y, CHEN K, et al. The water adaptability ofis modulated by soil nitrogen availability[J]. Biomass and Bioenergy, 2012, 47: 71–81
[4] WANG W F, ZONG Y Z, ZHANG S Q. Water-and nitrogen-use efficiencies of sweet sorghum seedlings are improved under water stress[J]. International Journal of Agriculture & Biology, 2014, 16: 285–292
[5] BOBBINK R, HICKS K, GALLOWAY J, et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis[J]. Ecological Applications, 2010, 20(1): 30–59
[6] SPARRIUS L B, SEVINK J, KOOIJMAN A M. Effects of nitrogen deposition on soil and vegetation in primary succession stages in inland drift sands[J]. Plant and Soil, 2012, 353(1/2): 261–272
[7] IPCC. Summary for policymakers of the synthesis report of the IPCC fourth assessment report[R]. Cambridge: Cambridge University Press, 2007
[8] YANG P, XIA J, ZHANG Y Y, et al. Temporal and spatial variations of precipitation in Northwest China during 1960-2013[J]. Atmospheric Research, 2017, 183: 283–295
[9] 张影, 徐建华, 陈忠升, 等. 中亚地区气温变化的时空特征分析[J]. 干旱区资源与环境, 2016, 30(7): 133–137 ZHANG Y, XU J H, CHEN Z S, et al. Spatial and temporal variation of temperature in central Asia[J]. Journal of Arid Land Resources and Environment, 2016, 30(7): 133–137
[10] 辛福梅, 杨小林, 赵垦田, 等. 干旱胁迫对拉萨半干旱河谷主要乔木树种幼树耗水及光合特性的影响[J]. 浙江大学学报: 农业与生命科学版, 2016, 42(2): 199–208 XIN F M, YANG X L, ZHAO K T, et al. Effect of drought stress on characteristics of water consumption and photosynthesis for main arbor species in semi-arid valley of Lhasa[J]. Journal of Zhejiang University: Agriculture & Life Sciences, 2016, 42(2): 199–208
[11] GU D X, WANG Q, OTIENO D. Canopy transpiration and stomatal responses to prolonged drought by a dominant desert species in central Asia[J]. Water, 2017, 9(6): 404
[12] KANG J J, ZHAO W Z, ZHENG Y, et al. Calcium chloride improves photosynthesis and water status in the C4succulent xerophyteunder water deficit[J]. Plant Growth Regulation, 2017, 82(3): 467–478
[13] ZHANG Y, XIE J B, LI Y. Effects of increasing root carbon investment on the mortality and resprouting ofseedlings under drought[J]. Plant Biology, 2017, 19(2): 191–200
[14] Peñuelas J, Filella I, LlusiÀ J, et al. Comparative field study of spring and summer leaf gas exchange and photobiology of thetreesand[J]. Journal of Experimental Botany, 1998, 49(319): 229–238
[15] DEWAR R C. A simple model of light and water use evaluated for[J]. Tree Physiology, 1997, 17(4): 259–265
[16] 李合生. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 1999: 164–260 LI H S. Principles and Techniques of Plant Physiological Biochemistry Experiment[M]. Beijing: Higher Education Press, 1999: 164–260
[17] 邹琦. 植物生理学实验指导[M]. 北京: 中国农业出版社, 2000: 104–160 ZOU Q. Instructional on Plant Physiology[M]. Beijing: China Agriculture Press, 2000: 104–160
[18] 李银坤, 武雪萍, 吴会军, 等. 水氮条件对温室黄瓜光合日变化及产量的影响[J]. 农业工程学报, 2010, 26(13): 122–129 LI Y K, WU X P, WU H J, et al. Effects of water and nitrogen conditions on the diurnal variation of photosynthesis and yield of cucumber in greenhouse[J]. Transactions of the CSAE, 2010, 26(13): 122–129
[19] 刘振香. 不同水肥处理对夏玉米田土壤理化性质及微生物特性的影响[D]. 泰安: 山东农业大学, 2014 LIU Z X. Effects of irrigation and nitrogen on physical and chemical properties and microbiologic properties of soil on summer corn field[D]. Tai’an: Shandong Agricultural University, 2014
[20] 陈亚, 代先强, 袁玲, 等. 水氮耦合对土壤理化性状及作物生长的影响研究进展[J]. 河南农业科学, 2009, 38(5): 11–15 CHEN Y, DAI X Q, YUAN L, et al. Advances of researches on effects of coupling water with nitrogen fertilizers on the physicochemical property of soil and crops growth[J]. Journal of Henan Agricultural Sciences, 2009, 38(5): 11–15
[21] 肖新, 储祥林, 邓艳萍, 等. 江淮丘陵季节性干旱区灌溉与施氮量对土壤肥力和水稻水分利用效率的影响[J]. 干旱地区农业研究, 2013, 31(1): 84–88 XIAO X, CHU X L, DENG Y P, et al. Effects of irrigation and nitrogen fertilization on soil fertility and water use efficiency of rice[J]. Agricultural Research in the Arid Areas, 2013, 31(1): 84–88
[22] 王聪宇, 曹国军, 耿玉辉, 等. 氮钾配施对吉林省湿润冷凉区玉米产量及钾素吸收积累的影响[J]. 吉林农业大学学报, 2015, 37(3): 332–337 WANG C Y, CAO G J, GENG Y H, et al. Effect of nitrogen and potassium interaction on yield and accumulation of spring maize in cold and humid region of Jilin Province[J]. Journal of Jilin Agricultural University, 2015, 37(3): 332–337
[23] 陈亚. 水氮耦合对植烟土壤理化生物特性及烤烟生长的影响[D]. 重庆: 西南大学, 2009 CHEN Y. Effects of water-nitrogen coupling on the growth of flue-cured tobacco and physicochemical and biological properties of soil[D]. Chongqing: Southwest University, 2009
[24] 李轶凡. 增氮增雨对贝加尔针茅草甸草原土壤氮矿化及土壤理化性质的影响[D]. 长春: 东北师范大学, 2015 LI Y F. Effects of stimulating nitrogen deposition and increasing precipitation on nitrogen mineralization in ameadow steppe[D]. Changchun: Northeast Normal University, 2015
[25] 韦泽秀. 水肥对大棚黄瓜和番茄生理特性及土壤环境的影响[D]. 咸阳: 西北农林科技大学, 2009 WEI Z X. Effect the condition of soil water and fertilizer on the physiological characteristics of plant and soil environments of cucumber and tomato in greenhouse[D]. Xianyang: North West Agriculture and Forestry University, 2009
[26] 闫慧, 吴茜, 丁佳, 等. 不同降水及氮添加对浙江古田山4种树木幼苗光合生理生态特征与生物量的影响[J]. 生态学报, 2013, 33(14): 4226–4236 YAN H, WU Q, DING J, et al. Effects of precipitation and nitrogen addition on photosynthetically eco-physiological characteristics and biomass of four tree seedlings in Gutian Mountain, Zhejiang Province, China[J]. Acta Ecologica Sinica, 2013, 33(14): 4226–4236
[27] 田媛, 塔西甫拉提•特依拜, 徐贵青. 梭梭与白梭梭气体交换特征对比分析[J]. 干旱区研究, 2014, 31(3): 542–549 TIAN Y, TAXIPLAT T, XU G Q. Gas exchange ofand.[J]. Arid Zone Research, 2014, 31(3): 542–549
[28] 刘晓宏, 肖洪浪, 赵良菊. 不同水肥条件下春小麦耗水量和水分利用率[J]. 干旱地区农业研究, 2006, 24(1): 56–59 LIU X H, XIAO H L, ZHAO L J. Effects of water and nitrogen condition on water consumption and water use efficiency of spring wheat[J]. Agricultural Research in the Arid Areas, 2006, 24(1): 56–59
[29] 王海江, 崔静, 侯振安, 等. 膜下滴灌棉花水氮耦合对其干物质和水分利用效率的影响[J]. 西北农业学报, 2010, 19(3): 76–80 WANG H J, CUI J, HOU Z A, et al. Impact of coupling of water and nitrogen on dry matter and water use efficiency of cotton under plastic mulching by drip irrigation[J]. Acta Agriculturae Boreali-occidentalis Sinica, 2010, 19(3): 76–80
[30] 柴仲平, 王雪梅, 孙霞, 等. 水氮耦合对红枣光合特性与水分利用效率的影响研究[J]. 西南农业学报, 2010, 23(5): 1625–1630 CHAI Z P, WANG X M, SUN X, et al. Photosynthetic characteristics and water use efficiency ofunder coupling of water and N[J]. Southwest China Journal of Agricultural Sciences, 2010, 23(5): 1625–1630
[31] 李银坤, 武雪萍, 武其甫, 等. 不同水氮处理对温室黄瓜产量、品质及水分利用效率的影响[J]. 中国土壤与肥料, 2010, (3): 21–24 LI Y K, WU X P, WU Q P, et al. Effects of different water and nitrogen treatment on the yield and quality and water use efficiency of cucumber in greenhouse[J]. Soils and Fertilizer Sciences in China, 2010, (3): 21–24
[32] 李静静, 陈雅君, 张璐, 等. 水氮交互作用对草地早熟禾生理生化与坪用质量的影响[J]. 中国草地学报, 2016, 38(4): 42–48 LI J J, CHEN Y J, ZHANG L, et al. Effects of water-nitrogen interaction on physiological-biochemical indexes and turf quality ofL.[J]. Chinese Journal of Grassland, 2016, 38(4): 42–48
[33] 李鑫, 张永清, 王大勇, 等. 水氮耦合对红小豆根系生理生态及产量的影响[J]. 中国生态农业学报, 2015, 23(12): 1511–1519 LI X, ZHANG Y Q, WANG D Y, et al. Effects of coupling water and nitrogen on root physio-ecological indices and yield of adzuki bean[J]. Chinese Journal of Eco-Agriculture, 2015, 23(12): 1511–1519
[34] 米美多, 慕宇, 代晓华, 等. 花后高温胁迫下不同施氮量对春小麦抗氧化特性的影响[J]. 江苏农业科学, 2017, 45(1): 52–56 MI M D, MU Y, DAI X H, et al. Effect of different nitrogen amounts on antioxidant properties of spring wheat after anthesis under high temperature stress[J]. Jiangsu Agricultural Sciences, 2017, 45(1): 52–56
[35] LI J, QU H, ZHAO H L, et al. Growth and physiological responses ofto sand burial stress[J]. Journal of Arid Land, 2015, 7(1): 94–100
[36] MITTAL S, KUMARI N, SHARMA V. Differential response of salt stress on: Photosynthetic performance, pigment, proline, D1 and antioxidant enzymes[J]. Plant Physiology and Biochemistry, 2012, 54: 17–26
[37] 翁伯琦, 郑向丽, 赵婷, 等. 不同生育期花生叶片蛋白质含量及氮代谢相关酶活性分析[J]. 植物资源与环境学报, 2014, 23(1): 65–70 WENG B Q, ZHENG X L, ZHAO T, et al. Analyses on protein content and enzyme activity involved in nitrogen metabolism of peanut () leaf at different growth stages[J]. Journal of Plant Resources and Environment, 2014, 23(1): 65–70
[38] 王景燕, 龚伟, 李伦刚, 等. 水肥对汉源花椒幼苗抗逆生理的影响[J]. 西北植物学报, 2015, 35(3): 530–539 WANG J Y, GONG W, LI L G, et al. Effects of water and fertilizer on physiological characteristics of stress resistance ofMaxim.‘Hanyuan’seedling[J]. Acta Botanica Boreali-Occidentalia Sinica, 2015, 35(3): 530–539
[39] 杨晓清, 周再知, 梁坤南, 等. 氮素对模拟胁迫下土沉香幼苗抗旱生理的影响[J]. 热带作物学报, 2013, 34(6): 1121–1127YANG X Q, ZHOU Z Z, LIANG K N, et al. Effects of nitrogen nutrition on physiological traits of theseedlings under simulated water stress[J]. Chinese Journal of Tropical Crops, 2013, 34(6): 1121–1127
Combined effect of simulated precipitation and nitrogen deposition on photosynthetic physiology ofin southern margin of Junggar Basin, China*
ZHANG Wei, ZHAO Wenqin**, XIE Shuangquan, ZHANG Li, LYU Xinhua, HE Yaling
(College of Life Sciences / College of Chemical Technology, Shihezi University, Shihezi 832000, China)
Precipitation and nitrogen deposition are two of the most investigated factors of global climate change which also are the controlling factors of desert ecosystems. This study examined the combined effects of precipitation and nitrogen deposition on arid ecosystems in order to provide insight into the responses of ecosystems to global climate change. We selected plantation ofin the southern margin of Junggar Basin. Two precipitation conditions [natural precipitation (W0) and 30% increased precipitation (W1)] and three levels of nitrogen depositions [natural nitrogen deposition (N0), 30 kg(N)·hm-2·a-1(N1) and 60 kg(N)·hm-2·a-1(N2) increased natural nitrogen deposition] were randomly added to simulate nitrogen deposition and precipitation for two years to explored the effects of precipitation, nitrogen deposition and the combined effects on diurnal change in photosynthetic and physio-ecological traits of. The results showed that precipitation, nitrogen deposition and the combined effects had direct significant correlation with diurnal change in net photosynthetic rate of. Based on the variation in net photosynthetic rate, intercellular CO2concentration and stomatal limitation of, “midday depression” of photosynthesis was mainly caused by non-stomatal factors. Moreover, with increased nitrogen deposition, there were decreasing physiological indexes (MDA content, POD activity, CAT activity, SOD activity, soluble protein content and soluble sugar content) under W0precipitation condition. However, proline content initially decreased before eventually increasing. There were initial increases followed by decreases in physiological indexes (MDA content, POD activity, CAT activity, SOD activity, soluble protein content, proline content and soluble sugar content) under W1precipitation condition. With the exception of W1N1, proline content was lower under nitrogen deposition increase treatments than under the natural deposition treatment. Also MDA content, antioxidant enzymes activities, soluble protein content and soluble sugar content ofwere also significantly lower under the other water/nitrogen treatments than under the control regimes. The results from the comprehensive analysis indicated that precipitation, nitrogen deposition and the combined effects benefited the growth of, but the strength of the combined effect depended on the ratio between the nitrogen deposition and precipitation.
; Precipitation; Nitrogen deposition; Interaction of water and nitrogen; Photosynthetic physiological traits; Southern margin of Junggar Basin
Corresponding author, E-mail: shz211985@163.com
Jun. 6, 2017; accepted Aug. 6, 2017
10.13930/j.cnki.cjea.170529
Q945.79
A
1671-3990(2018)01-0106-10
通信作者:赵文勤, 主要从事干旱区植物与植被研究。E-mail: shz211985@163.com 张维, 主要从事干旱区植物与植被研究。E-mail: 1182652644@qq.com
2017-06-06
2017-08-06
*The study was supported by the National Basic Research Program of China (973 Program) (2014CB954203) and the National Natural Science Foundation of China (31360139, 41561010, 31560177).
*国家重点基础研究发展计划(973计划) (2014CB954203)和国家自然科学基金项目(31360139, 41561010, 31560177)资助
张维, 赵文勤, 谢双全, 张莉, 吕新华, 贺亚玲. 模拟降水和氮沉降对准噶尔盆地南缘梭梭光合生理的影响[J]. 中国生态农业学报, 2018, 26(1): 106-115
Zhang W, Zhao W Q, Xie S Q, Zhang L, LYU X H, He Y L. Combined effect of simulated precipitation and nitrogen deposition on photosynthetic physiology ofin southern margin of Junggar Basin, China[J]. Chinese Journal of Eco-Agriculture, 2018, 26(1): 106-115
猜你喜欢
作物学报(2022年2期)2022-11-06
灌溉排水学报(2022年9期)2022-09-28
中国农业科学(2022年17期)2022-09-19
作物学报(2022年9期)2022-07-18
哈哈画报(2022年4期)2022-04-19
养殖与饲料(2021年4期)2021-04-17
科学大众(2020年17期)2020-10-27
河南农业·综合版(2020年10期)2020-10-20
河南农业(2020年10期)2020-10-15
安徽农学通报(2020年3期)2020-02-28
