
喀斯特区天峨槭(Acer wangchii)树干液流特征及其与环境因子的相关分析
2018-01-03 08:29:47杨文斌崔迎春赵文君舒德远侯贻菊丁访军
生态学报 2017年22期
吴 鹏, 杨文斌, 崔迎春, 赵文君, 舒德远, 侯贻菊,丁访军,*
1 中国林业科学研究院荒漠化研究所,北京 100091 2 贵州省林业科学研究院,贵阳 550005
喀斯特区天峨槭(Acerwangchii)树干液流特征及其与环境因子的相关分析
吴 鹏1, 2, 杨文斌1, 崔迎春2, 赵文君2, 舒德远2, 侯贻菊2,丁访军2,*
1 中国林业科学研究院荒漠化研究所,北京 100091 2 贵州省林业科学研究院,贵阳 550005
采用TDP(Thermal Dissipation Probe)热扩散探针法,于2012年2月—2014年3月,对喀斯特地区天峨槭(Acerwangchii)树干液流速率进行了长期连续的监测,并同步监测了林分空气温度(Ta)、相对湿度(RH)、太阳辐射(Solar)、风速(WS)、降雨量(Rainfall)、土壤含水量(SWC)等环境因子,探讨了不同时间尺度下天峨槭树干液流特征及其与环境因子的关系。结果表明:1)不同天气条件下的树干液流为晴天>阴天>雨天,且均呈明显的“昼高夜低”变化规律;2)天峨槭树干的日平均液流量为5.08 kg/d,不同季节表现为夏季((8.38±5.32) kg/d)>秋季((5.16±3.99) kg/d)>春季((4.86±3.77) kg/d)>冬季((1.94 ±1.40) kg/d);3)月平均液流量为153.64 kg,年平均液流量达1838.40 kg;4)小时尺度下,影响晴天、阴天、雨天全天树干液流的主要环境因子都是Solar,但各环境因子对树干液流的影响程度又因昼夜、季节及降雨量的不同而存在差异;5)从年度范围来看:小时尺度下,Solar、Ta、RH、SWC10cm和WS可以共同解释树干液流的63.50%;日尺度下,Solar、Ta、SWC10cm和Rainfall可共同解释其68.50%;月尺度下,单个因子Ta就能解释其74.80%;且随着时间尺度的缩小,各环境因子入选回归方程的个数有增加的趋势,而对树干液流的解释程度(R2)则有降低的趋势。对比其他地区研究结果,其环境因子对树干液流的影响差异都很大;但总的来说,无论在何种时间尺度上,Solar(或光合有效辐射(PAR))和Ta基本上都是影响树干液流的主要环境因子,且各环境因子对雨天树干液流的解释程度都不高,本研究亦如此;因此,以环境因子对雨天树干液流进行预测的时候可能会存在误差,应特别注意土壤含水量对其的影响。
喀斯特;天峨槭;树干液流;环境因子;不同时间尺度
植物蒸腾是土壤-植物-大气连续体水热传输过程中一个极为重要的环节[1],也是植物本身生长过程中极为重要的生理生态过程之一,植物体内超过90%的树干液流(忽略了树木新陈代谢过程中对极小部分水分的利用)都用以蒸腾[2];而水分是植物生长的主要限制性生态因子之一[3],树木对水分利用的主要动力是蒸腾需求,只有在蒸腾拉力的作用下[4- 5],这些水分才能被用于植物不同的组织器官,以保证其健康生长,因此可以用树干液流来表征植物的蒸腾量[6];通过对植物树干液流变化规律及其与相关环境因子关系的研究,对于理解植物水分需耗、估算不同时间尺度下植被蒸散具有重要意义[7- 8]。有研究表明,植物的蒸腾耗水主要受植物个体的生物学结构(树干直径、冠幅、边材面积等)[9- 10]、土壤水分[6,10-13]和气象因子[1- 2,6,8,10,12-14]等的影响;生物学结构主要决定了树干液流的潜力,土壤供水决定了树干液流的总体水平,而气象因子则决定了树干液流的瞬间变动[10,15]。
目前,已有的研究多是针对常态地貌上植物的树干液流来进行,而对喀斯特地貌上的研究甚少,仅张中峰[16]、黄玉清[17]、刘延惠[18]等人分别对岩溶地区的青冈栎(Cyclobalanopsisglauca)和短萼海桐(Pittosporumbrevicalyx)树干液流特征及其与环境因子的关系进行了研究。喀斯特地貌上的森林植被,其生境与相同气候条件下常态地貌上的明显不同[19],特殊的地貌构造使其生态水文过程更加复杂,对其树木蒸腾、生理生态过程以及对土壤间歇性干旱的适应机制尚缺乏足够认识。为此,本文以喀斯特森林顶极群落主要优势树种天峨槭(Acerwangchii)为研究对象,利用热扩散技术对其树干液流进行连续不间断的监测,并结合自动气象站对环境气象因子的同步测定数据,旨在探讨:1)不同时间尺度下天峨槭树干液流特征及其与环境因子的内在关系是否存在差异;2)不同时间尺度下环境因子对树干液流的拟合对简化分析树干液流是否具有意义;3)喀斯特地貌上树干液流特征及其对环境因子的响应与其他地区是否存在差异。以期能为喀斯特森林生态需水估算以及退化生态系统植被恢复与重建过程中树种的筛选提供基础数据。
1 研究区概况
研究区位于贵州喀斯特森林生态系统定位观测研究站,地处茂兰国家级自然保护区内(107°52.2′—108°5.7′ E,25°9.3′—25°20.8′ N)。保护区最低海拔430.0 m,最高1078.6 m,平均在550.0—850.0 m之间;属中亚热带季风湿润气候,具有春秋温暖、冬无严寒、夏无酷暑、雨量充沛的特点;年平均气温15.3℃,≥10℃积温4598.6℃;年平均降雨量1752.5 mm,集中分布于4—10月;年平均相对湿度83.0%;无霜期315 d;全年日照时数1272.8 h,日照百分率29.0%;太阳年辐射总量为63289.80 kW/m2。保护区内主要是由纯质石灰岩及白云岩构成的喀斯特地貌,裸岩率在80%以上,属于典型的喀斯特生境;土壤稀少,土层浅薄(20 cm左右)且不连片,多存于岩石缝隙之中,土壤富钙和富盐基化,有机质含量高。该保护区是目前世界上同纬度地区残存下来仅有的、原生性强且相对稳定的喀斯特森林生态系统,同时也是喀斯特区原生性森林(常绿落叶阔叶混交林)分布面积最大的区域[20]。
2 研究方法
2.1 样地调查与样木选择
2012年,对站区原有的4个固定样地(30 m×30 m)进行了调查[21],样地基本概况(表1)。

表1 样地基本概况
根据调查结果,在样地内选取径级适中(接近样地内的平均胸径)且生长良好、树干通直、无病虫害、冠幅相似的3株天鹅槭作为监测样树(表2)。
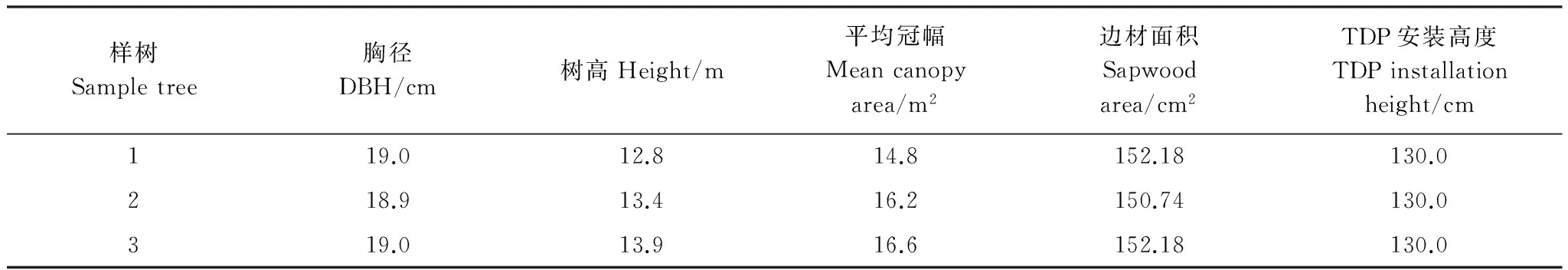
表2 天峨槭样树基本特征
2.2 树干液流测定
利用热扩散液流探针(TDP- 30,Dynamax Inc., Houston, USA)产生的温差来计算天峨槭树干液流速率,TDP探针缆线与CR1000数据采集器连接,以10 min时间间隔进行数据采集;采集时间:2012年2月25日—2014年3月22日;根据Granier经验公式[22]来换算树干边材瞬时液流速率(Js):
Js=0.0119×[(ΔTmax-ΔT)/ΔT]1.231
(1)
式中,Js是液流密度(瞬时液流速率)(g H2O/(m2/s)),即单位时间通过单位边材面积的液流量[23];ΔTmax为上下探针间的最大昼夜温差(℃),ΔT瞬时温差(℃)。
Fs=Js×SA×3600
(2)
式中,Fs为液流通量(每小时的液流速率)(g H2O/h);SA为胸径处边材面积;3600是时间换算系数[23]。
边材面积(SA)的确定:在样地外选择12株不同胸径的天峨槭,在树干胸径处用生长锥钻取样芯。将样芯带回实验后,用游标卡尺分别测量样芯的心材宽、边材宽、半径和皮厚等值,计算样木边材面积和胸径,建立了天峨槭边材面积与胸径的回归关系式:
SA=0.07426×D1.8077(R2=0.7898)
(3)
式中,D为样木的胸径,R为相关系数。
日间或夜间的树干液流对日总液流量的贡献率,计算公式如下[12]:

(4)
式中,CR为日间或夜间的树干液流对日累积量的贡献率(%);Fsi为i时间的液流速率(g H2O/h);i为时间段(1、2、…、t);t为日间(以太阳辐射大于零定义为“日间”)或夜间(太阳辐射等于零为“夜间”)的时长;24为全天时长。
基于小时尺度下的液流速率(Fs)可以计算日尺度下的液流量(Fd)和月尺度下的液流量(Fm):
(5)
(6)
式中,Fd为日液流量(kg H2O/d);0.001是单位换算系数;Fm为月液流量(kg H2O);n为每月的天数;Fdj为该月第j天的日液流量。
2.3 环境因子测定
环境因子由安装在观测塔上的Campbell自动气象站数据采集器同步采集,主要包括:太阳辐射(Solar)、气温(Ta)、相对湿度(RH)、风速(WS)、降雨量(Rainfall)、不同层次的土壤含水量(SWC10cm、SWC20cm、SWC40cm)等。
2.4 数据处理方法
1)异常值的处理:首先建立液流速率正常值与空气温度和太阳辐射的拟合模型,然后利用异常值(缺失值)所对应的空气温度和太阳辐射,反过来计算其液流速率,用该液流速率来代替其异常值(缺失值);异常值的判别方法参考文献[4,24]。
2)小时尺度下的时滞效应:采用错位对比法来确定[25]。
3)数据分析:采用Excel 2010进行绘图,SPSS 22.0进行方差分析(Repeated-measures ANOVA)、多重比较(LSD最小显著差法)、相关性分析、逐步回归分析及通径分析等;通径系数计算方法参见文献[26]。
3 结果与分析
3.1 小时尺度下树干液流特征及其与环境因子的相关性
3.1.1典型晴天和阴天树干液流特征
选取典型晴天和阴天天峨槭树干液流的平均值,按月、分不同季节(春季:3—5月,夏季:6—8月,秋季:9—11月,冬季:12—次年2月)分析树干液流的日变化(图1)。小时尺度下,树干液流的日变化表现为晴天(388.60g/h)>阴天(216.30g/h);不同季节表现为夏季(686.35g/h晴和359.06g/h阴)>春季(393.68g/h晴和219.23g/h阴)>秋季(372.19g/h晴和198.79g/h阴)>冬季(102.19g/h晴和88.00g/h阴),且不同天气条件下冬季与其他季节差异均显著(P<0.05),夏季与秋季差异均显著(P<0.05),而其他季节间差异则均不显著(P>0.05);相同天气条件下,同一季节不同月份树干液流的日变化亦存在差异。从日变化的曲线波动来看,除冬季以外,晴天树干液流的日变化多呈平滑的“单峰”曲线;阴天则多为“单峰”“双峰”和“多峰”曲线并存。

图1 不同季节、不同天气条件下(典型晴天和阴天)天峨槭树干液流的日变化Fig.1 Diurnal variation of stems sap flow in Acer wangchii under different weather conditions of different seasons
由表3可知,不同季节晴天树干液流从“‘启动’→达到‘峰值’→回落到‘低值’”的整个过程较阴天均呈现出:启动时间较早、到达峰值和回落到低值时间较晚、整个过程持续时间较长的变化规律;由“夏季→冬季→夏季”,不同天气条件下树干液流的启动和到达峰值的时间均表现为先“推后”后“提前”,回落到低值的时间均为先“提前”后“推后”,整个过程持续时间均为先“缩短”后“延长”。
3.1.2典型雨天树干液流特征
按照降雨量的大小(中雨:10.0—24.9mm、大雨:25.0—49.9mm),选取连续降雨2—3d的数据作为典型雨天树干液流的日变化(图2)。
雨天树干液流的日变化多呈“多峰”或不规则上下波动,平均值为64.27g/h,且中雨天(70.52g/h)要高于大雨天(58.03g/h);与当年同月晴天和阴天树干液流的日变化相比,雨天的要显著小于晴天和阴天(P<0.05)。
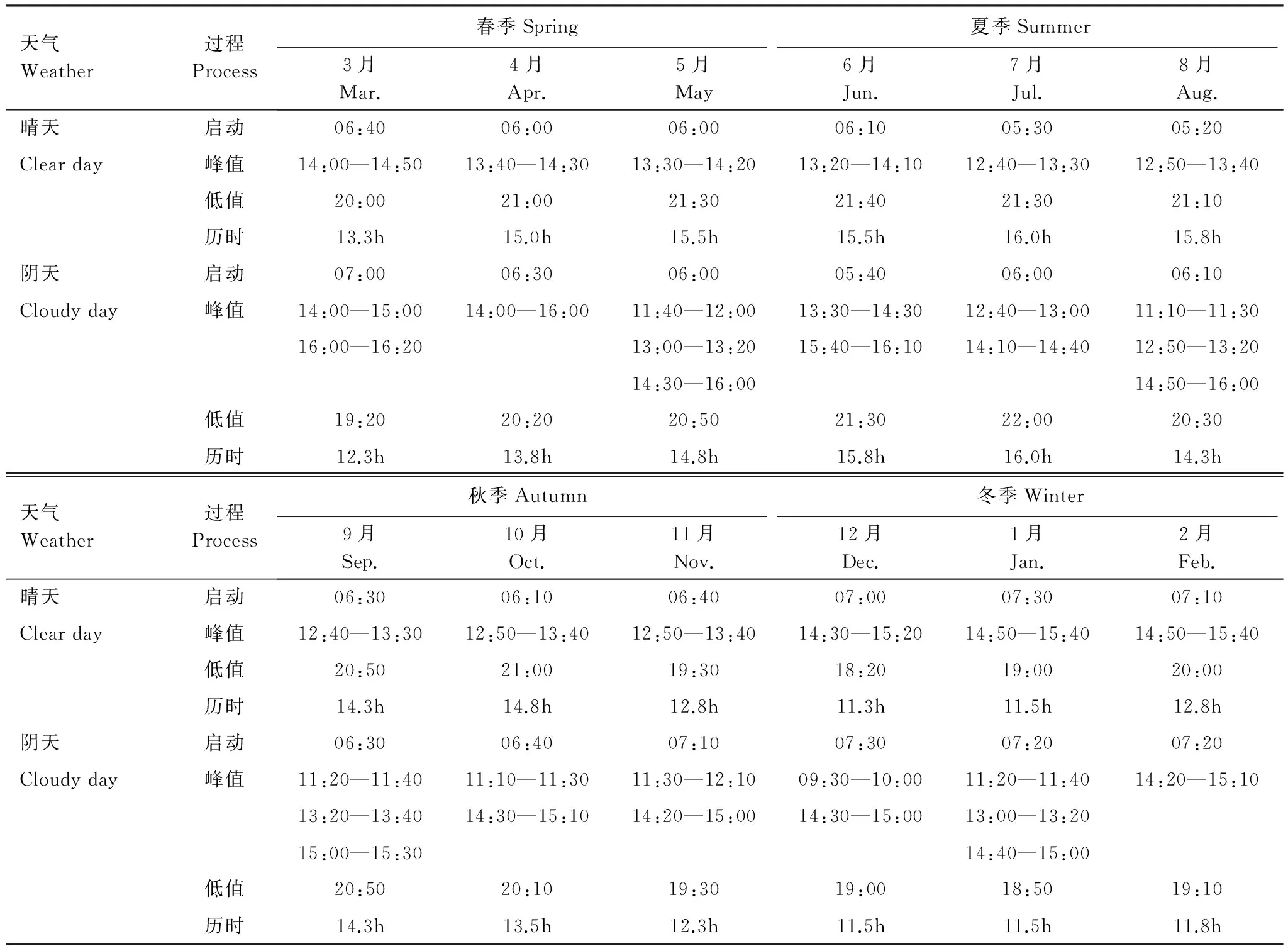
表3 不同季节、不同天气条件下树干液流日变化持续过程时间表
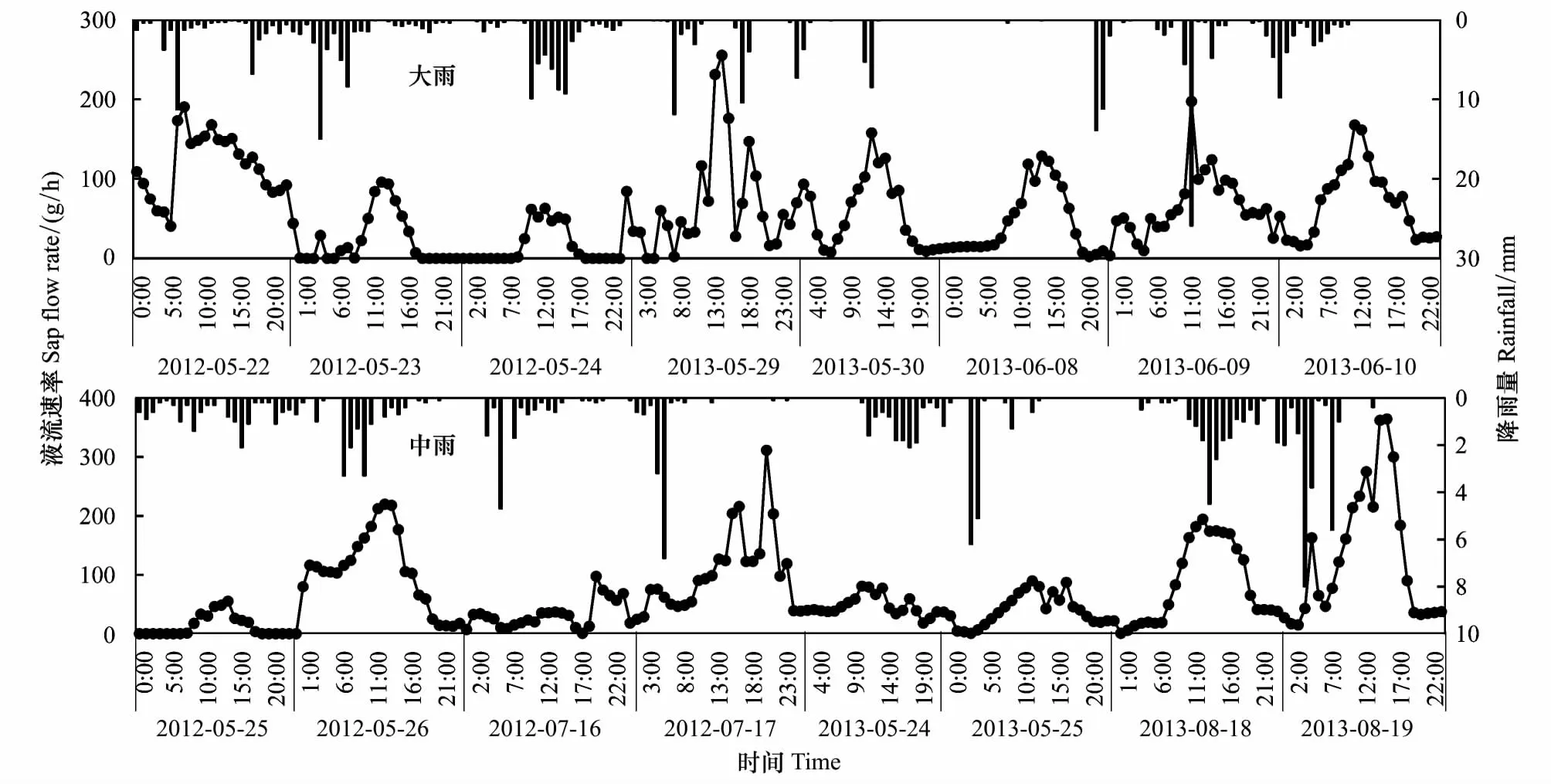
图2 典型雨天天峨槭树干液流和降雨量的日变化Fig.2 Diurnal variation of rainfall and stems sap flow in Acer wangchiiof rainy day
3.1.3小时尺度下树干液流的昼夜差异
不同天气条件下的树干液流均呈明显的“昼高夜低”变化规律,且差异均显著(P<0.05);晴天、阴天、雨天,日间树干液流的平均值分别是夜间的12.37倍、6.59倍和2.49倍。另外,不同天气条件下日间和夜间树干液流对日总液流量的贡献率也不同;晴天,昼、夜贡献率的平均值分别为90.53%和9.47%,阴天分别为84.08%和15.92%,雨天分别为77.26%和22.74%,夜间贡献率以雨天为最高,阴天次之,晴天最低。晴天和阴天不同季节夜间的贡献率则均以冬季为最高,春季次之,夏季最低(表4)。

表4 小时尺度下树干液流的昼夜差异/(g/h)
同列之间凡标有不同小写字母的表示差异显著(P<0.05);x为对日总液流量的贡献率
3.1.4小时尺度下树干液流与环境因子的相关性
综合图1、图2可知,不同天气条件下树干液流均受环境因子的影响,但其影响方式、作用大小并不清楚。为此,我们进一步分析了各环境因子(Solar、Ta、RH、WS、SWC10cm、SWC20cm、SWC40cm和Rainfall)与树干液流的相关系数、直接及间接通径系数(表5)。
(1)树干液流昼夜变化与环境因子的相关性
由表5可知(根据相关系数、直接和间接通径系数绝对值的大小进行判断),不同天气条件下,Solar都是影响全天树干液流的主要环境因子;Solar、Ta是影响晴天和阴天日间的主要环境因子,而Solar、SWC10cm是影响雨天日间的主要环境因子;Ta、RH是影响晴天和阴天夜间的主要环境因子,而SWC10cm是影响雨天夜间的主要环境因子。另外,不同天气条件下树干液流对土壤水分的利用和响应也存在差异,晴天和阴天受日间较强的Solar和Ta等环境因子的影响,致使土壤表层含水量减小,较深层的土壤水分就成了夜间树干液流主要的水分来源,所以SWC20cm对晴天和阴天夜间的树干液流有较明显的促进作用;雨天全天及昼夜,由于日间太阳辐射小、温度较低,树木蒸腾潜力小,且水分供应充足,极高的土壤表层含水量(SWC10cm)反而对树干液流产生了抑制作用。

表5 小时尺度下树干液流昼夜变化与环境因子的通径分析结果
**P<0.01,*P<0.05;Ta、RH、WS、Solar、SWC分别代表空气温度Air temperature、相对湿度Relative humidity、风速Wind speed、太阳辐射Solar radiation和土壤含水量Soil water content
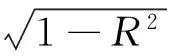

表6 小时尺度下树干液流昼夜变化与环境因子的逐步回归分析结果
(2)树干液流季节变化与环境因子的相关性
不同天气条件下各环境因子对树干液流的响应及与其拟合的回归方程因季节的不同也存在差异(表7,因篇幅所限,此表省略了间接通径系数的计算)。晴天,影响春季、秋季和冬季树干液流的环境因子主要是Solar和Ta,直接通径系数都在0.5左右;影响夏季的主要是Solar(0.602),Ta对其直接影响较小(0.149)。阴天,不同季节树干液流受环境因子的影响差异较大,春季主要受Solar、Ta和RH的影响,夏季的是RH,秋季的是Solar,冬季则受Solar和Ta的影响较大。大雨天,主要受Solar、SWC10cm和Ta的影响,相较于Ta对晴天、阴天树干液流明显的促进作用,Ta对大雨天树干液流抑制作用明显;中雨天,影响树干液流的主要是Solar。不同天气条件,RH对树干液流多是抑制作用;而WS对树干液流的作用因天气和季节的不同而存在差异。逐步回归分析结果表明(表7):入选方程的各环境因子与树干液流的相关系数以及各拟合方程的回归系数(R2)均达到极显著水平(P<0.01);晴天,不同季节各环境因子对树干液流的解释程度在82.00%—93.40%之间;阴天,各环境因子对树干液流的解释程度在81.60%—93.90%之间;雨天,各环境因子对树干液流的解释程度较低,大雨天和中雨天分别为39.80%和47.70%。
3.2 日尺度下树干液流特征及其与环境因子的相关性
由图3可知,天峨槭的日液流量在0.01—24.00 kg/d之间,平均值为5.08 kg/d。不同季节日平均液流量分别为:春季((4.86±3.77) kg/d)、夏季((8.38±5.32) kg/d)、秋季((5.16±3.99) kg/d)和冬季的((1.94 ±1.40) kg/d),除春季和秋季间差异不显著以外(P>0.05),其余各季节间均达到极显著差异(P<0.01);冬季的日液流量最低,低温(7.56℃)和少的降雨量(104.75 mm)可能是其主要限制因子;夏季的日液流量最高,主要得益于高温(23.33℃)和强太阳辐射(134.15 W/s)致使其蒸腾潜力很大,且水分供应充足(降雨量1121.00 mm),致使其液流量很高;秋季的日液流量显著低于夏季,可能是由于该季节降雨量偏少(412.70 mm)、土壤含水量过低等原因造成的;春季的降雨量虽然充足(1002.40 mm),但其空气温度(16.48℃)和太阳辐射(110.70 W/s)较夏季要低,致使其蒸腾潜力不高,使得其日液流量也显著低于夏季。
逐步回归分析结果表明(表8):入选方程的各环境因子对树干日液流量的直接作用以及方程的回归系数均达到显著(P<0.05)或极显著水平(P<0.01),说明所拟合方程均能很好的揭示不同季节及全年树干日液流量与环境因子的变化规律;除冬季各环境因子对日液流量的解释程度(33.90%)较低以外,其余各季节的解释程度均超过了70.00%。从全年来看,各环境因子对其解释程度也高达68.50%,而Solar和Ta是影响全年日液流量的主要环境因子,可解释日液流量的65.80%,占总解释程度的96.06%。
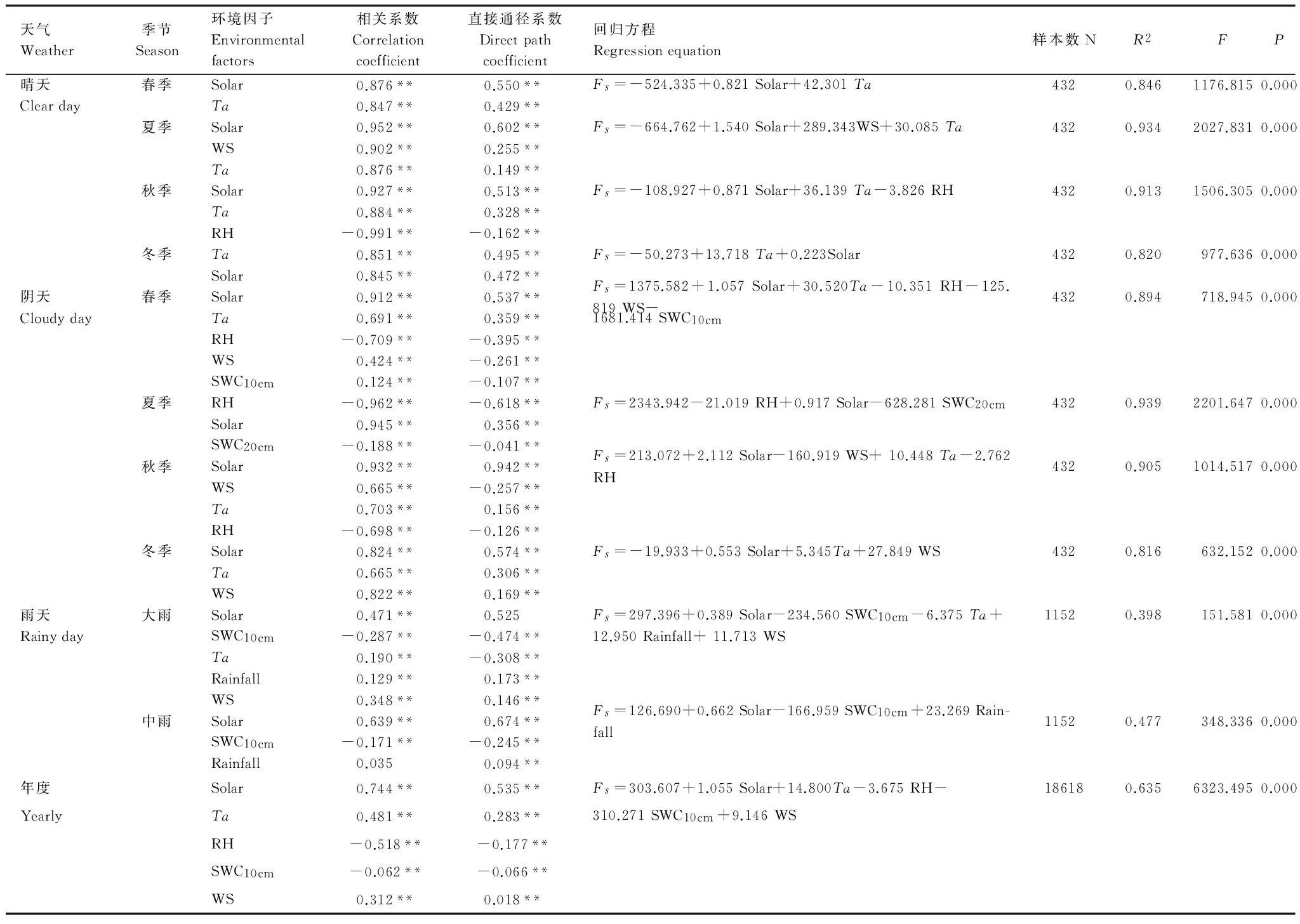
表7 小时尺度下树干液流季节变化与环境因子的逐步回归分析结果
注:**P<0.01,*P<0.05
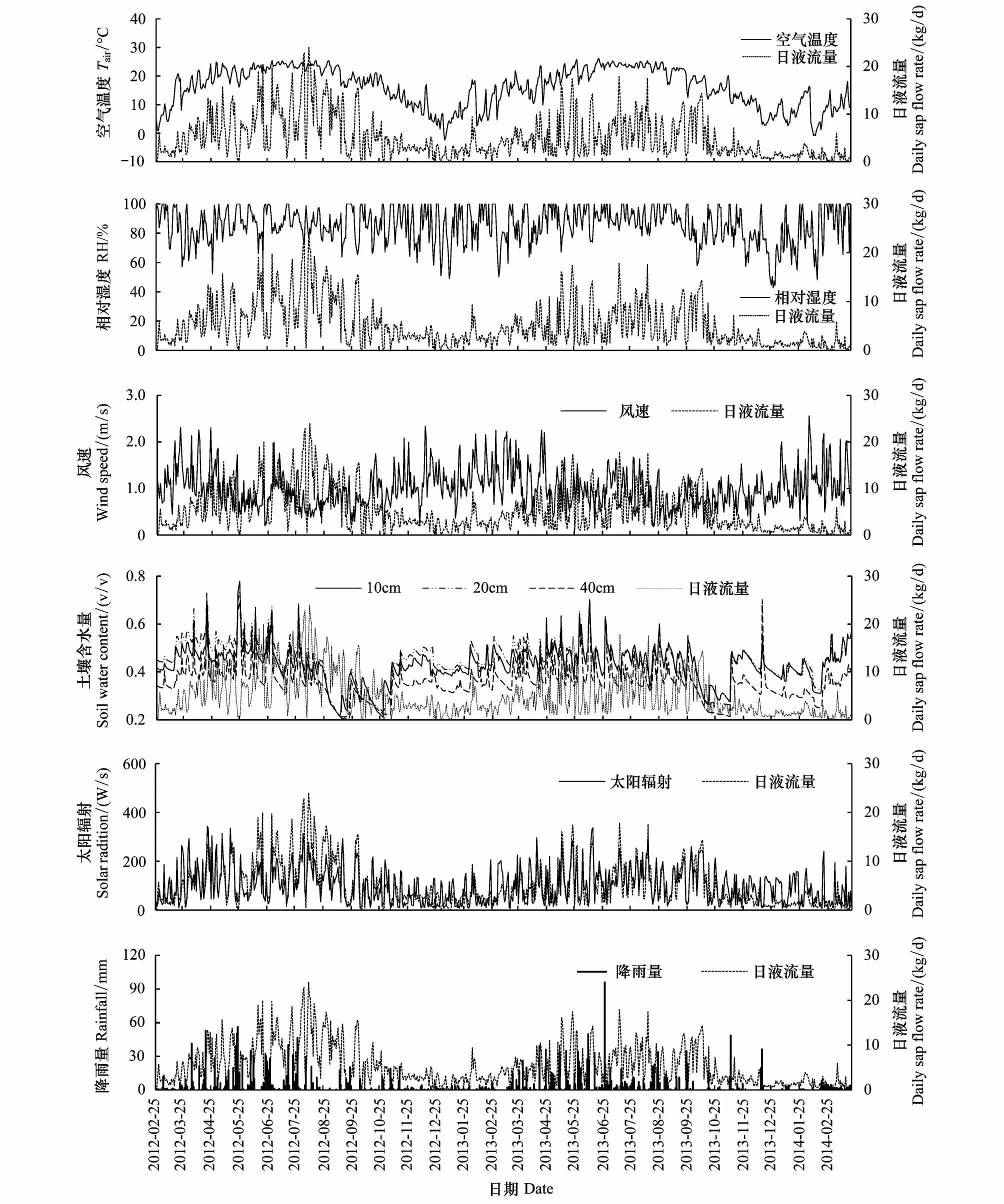
图3 天峨槭树干液流与空气温度、相对湿度、太阳辐射、土壤含水量、风速及降雨量的日均值变化Fig.3 Changes of daily of stems sap flow in Acer wangchii Tair, RH, Solar radiation, SWC, WS and Rainfall

尺度Scale季节Season环境因子Environmentalfactors相关系数Correlationcoefficient直接通径系数Directpathcoefficient回归方程Regressionequation样本数NR2FP日尺度春季Solar0.768**0.620**Fd=3.043+0.029Solar+0.273Ta-2060.723175.8240.000DailyscaleTa0.566**0.363**12.291SWC10cmSWC10cm-0.217**-0.192**夏季RH-0.818**-0.533**Fd=25.188-0.366RH+0.029So-lar+1840.794172.0820.000Solar0.810**0.359**0.572Ta-1.605WSTa0.368**0.159**WS-0.081-0.105**秋季Solar0.816**0.742**Fd=0.457+0.043Solar+0.176Ta-1820.724155.9020.000Ta0.477**0.172**6.298SWC10cmSWC10cm-0.272**-0.136**冬季SWC20cm0.452**0.400**Fd=-6.573+12.852SWC20cm-1850.33923.0390.000Rainfall-0.105-0.220**0.093Rainfall+0.009Solar+0.031RHSolar0.175**0.335**RH0.225**0.339**年度Solar0.753**0.581**Fd=1.119+0.036Solar+0.246Ta-7570.685407.9760.000Ta0.599**0.374**18.133SWC10cm-0.025RainfalSWC10cm-0.114**-0.141**Rainfall-0.139**-0.049*月尺度Monthlyscale年度Ta0.865**0.865**Fm=-58.540+13.207Ta250.74868.1330.000
3.3 月尺度下树干液流特征及其与环境因子的相关性
由图4可知,在月尺度下(2012年2月份只有5天数据,不能代表该月的情况,故舍去;2014年3月份有22天的数据,基本上可以代表该月的情况,故对其保留,为便于比较,该月的月液流量=该月日液流量的平均值×该月的天数),天峨槭的月平均液流量为153.64 kg,其中以2012年8月份的417.61 kg最高,2014年2月份的37.48 kg最低。不同年际间同一月份或同一季节的液流量因环境因子的变化也存在差异;以3月份为例,2012年、2013年和2014年的月液流量分别为86.68、131.19 kg和37.68 kg,2013年分别是2012年和2014年的1.51倍和3.48倍;2012年春、夏、秋、冬各季的树干液流总量分别为:502.36、959.59、490.56、218.57 kg,2013年的则分别为472.36、582.46、448.64、128.78 kg,2012年夏季和冬季的树干液流总量分别为2013年同季的1.65倍和1.70倍。2012年3月1日—2014年3月22日间,天峨槭树干液流总量为3830.05 kg,年平均液流量可达1838.40 kg。逐步回归分析结果显示(表8),在月尺度下,Ta是影响其液流量的主要环境因子,与液流量呈极显著的正相关关系(P<0.01),可解释液流量的74.80%,Ta每升高1℃,液流量可增加13.21 kg。
4 结论与讨论
小时尺度下,天峨槭树干液流的日变化表现为晴天>阴天>雨天(图1、图2),这主要是由于晴天Solar强、Ta高、RH小,植物蒸腾量大,致使其液流也大,而阴雨天Solar弱、RH大,植物蒸腾量小,致使其液流也较小,另外,雨天极高的RH使叶片内外的蒸汽压梯度降低,促使叶片气孔关闭,也制约了树干液流的上升[27]。晴天树干液流主要受Solar和Ta的影响,而这两个环境因子在晴天多为“单峰”变化,且波动较小,使得其树干液流也多呈平滑“单峰”曲线变化;阴天树干液流主要受Solar的影响,而Solar在阴天上下波动较大,致使其树干液流出现了“单峰”“双峰”和“多峰”并存的现象;雨天除了各环境因子对其影响以外,还有“根压”[27]以及降雨和光照的间断性等对其的影响,致使其日变化呈“多峰”或不规则上下波动;此结果与张璇[12]、张中峰[16]、刘华[28]、凡超[29]等人的研究结果基本一致。有研究认为,在中午前后,植物为了保存体内水分,会短暂关闭或减小叶片气孔开合程度,使其蒸腾量降低,导致树干液流出现明显或较明显的“午休”现象[30- 31];而在本研究中,除了天气原因导致树干液流呈多个峰值或上下波动以外,并未出现明显的“午休”现象;此结果与张中峰等人[16]的研究结果一致;其原因可能是植物为了适应喀斯特区高温高湿的环境以及生境的高度异质性而做出的水分调控策略,也可能是由于该区域太阳辐射的峰值不足以使植物的叶片气孔关闭而形成“光合午休”现象[12]。
与大多数研究结果一致[1,10,12- 13,29,31- 32],不同天气条件下天峨槭树干液流的日变化均呈明显的“昼高夜低”变化规律,昼、夜树干液流因季节的不同及降雨量的大小而存在差异。有学者认为,树干的夜间液流是为了弥补日间蒸腾导致的水分亏缺而在树体内储存水分[30,33],白天强烈的蒸腾作用,使树体与土壤之间形成了较大水势差,夜间在蒸腾停止后,在根压作用下根系吸水方式由日间的被动变为夜间的主动,持续保持吸水能力,导致了夜间液流活动的存在[30- 31];本研究中,晴天和阴天树干夜间液流的平均值分别为53.32 g/h和56.02 g/h,均要高于雨天的35.79 g/h,其原因主要是因为雨天植物的蒸腾量小,且由于降雨的存在,相对湿度高,水分供应充足,植物体内的水分亏缺不明显,致使雨天的夜间液流较晴天和晴天要低;但是从夜间液流对日总液流量的贡献率来看,由于雨天夜间液流速率与日间的差距较晴天、阴天夜间和日间的差距要小的多,导致其夜间液流的贡献率反而最高。
图4 天峨槭树干液流与空气温度、相对湿度、太阳辐射、土壤含水量、风速及降雨量的月均值变化Fig.4 Changes of monthly total of stems sap flow in Acer wangchii, Tair, RH, Solar radiation, SWC, WS and Rainfall
天峨槭树干的日、月平均液流量分别为5.08 kg/d和153.64 kg,年液流量达1838.40 kg,是湖南会同杉木(Cunninghamialanceolata)树干液流的1.5倍以上[34],比华北落叶松(Larixprincipis-rupprechtii)生长季的日液流量(3.17 kg/d)还要高[35];这可能与本研究所选样树边材面积较大有关,但相较其生长的喀斯特生境(土壤稀少,土层浅薄,地表水缺乏),也可能是得益于其植物根系能从更深的岩溶内部含水层中吸取水分的能力[17];但要低于程静等[32]对鼎湖山针阔混交林主要优势树种树干液流的研究结果;这可能是因为树干液流的大小受多种因子的影响,立地条件、气象因子、土壤温度、水分以及土壤肥力等因子的不同,都能导致植物对水分的利用产生很大差异[32]。还有研究表明,同一树种即使胸径相同或相近,其液流差异也很大[34,36],这可能是由于树种之间生物学结构的不同而导致的。
小时尺度下,影响晴天、阴天、雨天全天树干液流的主要环境因子都是Solar,但各环境因子对树干液流的影响又因昼夜差异、季节不同以及降雨量的大小而存在差异(表5、表7)。有研究认为[37],晴天日间WS有利于植物的蒸腾作用,而夜间或阴雨天WS对液流影响不大,WS过大还会降低液流水平;在本研究中,WS对夜间的树干液流都有明显的促进作用,反而对晴天日间的作用不明显,对阴雨天日间的树干液流还存在抑制作用,这个可能与不同的生境条件有关。
土壤含水量(SWC)对树干液流影响较为复杂。一般来说,在小尺度上,SWC对树干液流无显著影响,适宜在月、季节等大尺度上对其变化进行分析[38];有学者认为[12],SWC对雨天夜间的树干液流影响显著,它的变化是导致雨天夜间液流大于晴、阴天的主要因素;而王艳兵等人[33]通过对华北落叶松树干液流的研究得出:SWC0—40cm是限制夜间树干液流的主要因子,且月均土壤含水量对月蒸腾贡献率影响显著,但对雨天夜间树干液流影响不显著(P>0.05);还有研究认为,土壤水分亏缺在一定程度上能限制生长盛期刺槐(Robiniapseudoacacia)的蒸腾作用[10],但对展叶期刺槐树干液流的影响不大[14];曹文强等[27]则认为,SWC是解释雨天辽东栎(Quercusliaotungensis)树干液流的主要环境因子,但对晴天和阴天树干液流的影响不大;但张晓艳等[39]通过对民勤梭梭(Haloxylonammodendron(C. A. Mey)Bunge)树干液流的研究,得出了与其正好相反的结论:土壤含水量与晴天树干液流的相关性显著,但与阴雨天相关性则不显著;其原因可能与树种的不同以及树种根系对当地土壤水分的适应机制有关。本研究中,小时尺度下,SWC10cm是影响雨天全天和日间、夜间的主要环境因子,且对树干液流有较强的抑制作用;SWC20cm对晴天和阴天夜间的树干液流有明显的促进作用,但对晴天日间和全天的树干液流存在抑制作用。在日尺度下,SWC10cm对春季、秋季和全年的树干液流抑制作用较明显;而SWC20cm是影响冬季树干液流最主要的环境因子(表5、表7、表8);此结果与刘延惠等[18]对该地区短萼海桐树干液流的研究亦存在一定差异,其原因可能是由于两树种在同一群落中所处的层次不同,造成了对水分利用机制的差异。
从年度范围来看,不同时间尺度下,各环境因子对树干液流的影响也存在一定差异(表7、表8);小时尺度下,Solar、Ta、RH、SWC10cm和WS可以共同解释树干液流的63.50%,其中以Solar的直接影响最大,而RH和SWC10cm对其则有明显的抑制作用;日尺度下,Solar、Ta、SWC10cm和Rainfall可以共同解释树干液流的68.50%,其中以Solar的直接影响最大,Ta次之,Rainfall最小;月尺度下,单个因子Ta就能解释树干液流74.80%;我们发现,随着时间尺度增大,与树干液流显著相关的环境因子个数有减少的趋势,而对其解释程度则有增加的趋势,此结果与王文杰等[4]对兴安落叶松(Larixgmelinii)树干液流的研究结果一致,他们还认为,影响树干液流的主要环境因子随着时间尺度的增大,有从地上气象因子向地下土壤环境因子(土壤温度、湿度等)转变的趋势,本研究因没有对土壤温度的监测,故无法论证此结论。
对比其他地区研究结果:王文杰等[4]认为,在小时尺度上,光合有效辐射(PAR)、5 cm土壤温度(Ts5 cm)、RH和Ta是影响兴安落叶松树干液流的主要环境因子;在日尺度上,Ts5cm、PAR和Ta是其主要影响因子;在月尺度上,Ts5cm和SWC5cm对其影响显著。赵仲辉等[34]的研究则得出,在小时尺度上,Solar、Ta和Ts20cm是影响杉木树干液流的主要环境因子;在日尺度上,Ta、SWC30cm、Solar和SWC20cm是其主要影响因子;在月尺度上,Ta和RH对其影响显著。而通过对晋西黄土区辽东栎树干液流的研究,张建国等[40]认为空气水汽压亏缺(VPD)对树干液流能进行很好的拟合,隋旭红等[30]则认为Ta和Solar是影响其树干液流的主要环境因子,而陈宝强等[41]的研究则得出:不同月份影响树干液流的环境因子也不同,5、6月份主要是VPD和PAR对其产生影响,而7、8月份则主要是VPD和Ta对其影响显著;曹文强等[27]通过对太岳山辽东栎树干液流的研究则得出:晴天影响其树干液流的主要环境因子是Solar、Ta和RH,阴天主要是Solar和RH,而雨天则是SWC5—10cm、Rainfall和Ta。对比同处喀斯特地区的树干液流与环境因子的研究结果,其差异也很大[16- 18]。
总的来说,由于地域、物种、生境、所监测的环境因子、时间尺度、天气条件以及季节等的不同,还有数据分析方法的差异等等,即使处于同一生境中的同一物种,各环境因子对其树干液流的影响差异也很大。但是,我们也发现,无论在何种时间尺度上,Solar(或PAR)和Ta基本上都是影响各树种树干液流的主要环境因子[1,2,10,13- 14,16,18,24,27- 31,34- 35,39],究其原因,Solar(或PAR)作为光合作用的主要驱动力,决定了叶片气孔的开张程度,从而主要影响着植物蒸腾的强弱[34]。另外,我们还发现,雨天各环境因子对树干液流的解释程度较晴天和阴天要低的多[13,27,42],本研究亦是如此;这可能是由于雨天Solar弱,RH高,再加上降雨事件的发生(降雨的历时、强度、雨量的大小、间断性等因素)又对Solar、PAR、Ta、SWC、WS、Ts等环境因子产生影响,使得影响树干液流的环境因子众多,另外还有“根压”的存在[27],使其雨天树干液流的影响因子较晴天和阴天要复杂的多;当然,这也可能与不同天气条件下物种自身的生理特性有关[36],具体原因还有待于进一步深入研究;所以,以环境因子对雨天的树干液流进行预测的时候可能会存在误差,尤其应注意不同土层深度的土壤含水量对其树干液流的影响。
[1] 黄德卫, 张德强, 周国逸, 刘世忠, Dennis O, 李跃林. 鼎湖山针阔叶混交林优势种树干液流特征及其与环境因子的关系. 应用生态学报, 2012, 23(5): 1159- 1166.
[2] 徐世琴, 吉喜斌, 金博文. 典型荒漠植物沙拐枣茎干液流密度动态及其对环境因子的响应. 应用生态学报, 2016, 27(2): 345- 353.
[3] Hansen S. Daisy, a flexible Soil-Plant-Atmosphere system Model. 2002. [2017-02- 13].http://www.dina.kvl.dk/daisy/ftp/DaisyDescription.pdf.
[4] 王文杰, 孙伟, 邱岭, 祖元刚, 刘伟. 不同时间尺度下兴安落叶松树干液流密度与环境因子的关系. 林业科学, 2012, 48(1): 77- 85.
[5] Ford C R, Hubbard R M, Kloeppel B D, Vose J M. A comparison of sap flux-based evapotranspiration estimates with catchment-scale water balance. Agricultural and Forest Meteorology, 2007, 145(3/4): 176- 185.
[6] 张雷, 孙鹏森, 刘世荣. 树干液流对环境变化响应研究进展. 生态学报, 2009, 29(10): 5600- 5610.
[7] Bleby T M, Mcelrone A J, Jackson R B. Water uptake and hydraulic redistribution across large woody root systems to 20m depth. Plant, Cell and Environment, 2010, 33(12): 2132- 2148.
[8] 于贵瑞, 王秋凤. 植物光合、蒸腾与水分利用的生理生态学. 北京: 科学出版社, 2010.
[9] Braun P, Schmid J. Sap flow measurements in grapevines (VitisviniferaL.) 1. stem morphology and use of the heat balance method. Plant and Soil, 1999, 215(1): 39- 45.
[10] 吴芳, 陈云明, 于占辉. 黄土高原半干旱区刺槐生长盛期树干液流动态. 植物生态学报, 2010, 34(4): 469- 476.
[11] Ford C R, Goranson C E, Mitchell R J, Will R E, Teskey R O. Diurnal and seasonal variability in the radial distribution of sap flow: predicting total stem flow in Pinus taeda trees. Tree Physiology, 2004, 24(9): 941- 950.
[12] 张璇, 张会兰, 王玉杰, 王云琦, 刘春霞, 杨坪坪, 潘声雷. 缙云山典型树种树干液流日际变化特征及与气象因子关系. 北京林业大学学报, 2016, 38(3): 11- 20.
[13] 王小菲, 孙永玉, 李昆, 张春华, 李彬. 山合欢树干液流的季节变化. 生态学杂志, 2013, 32(3): 597- 603.
[14] 于占辉, 陈云明, 杜盛. 黄土高原半干旱区人工林刺槐展叶期树干液流动态分析. 林业科学, 2009, 45(4): 53- 59.
[15] 孙慧珍, 周晓峰, 康绍忠. 应用热技术研究树干液流进展. 应用生态学报, 2004, 15(6): 1074- 1078.
[16] 张中峰, 黄玉清, 李先琨, 莫凌, 焦继飞, 尤业明. 岩溶区青冈栎树干液流特征及其与环境因子关系. 中国岩溶, 2008, 27(3): 228- 234.
[17] 黄玉清, 张中峰, 何成新, 赵平, 袁维园, 焦继飞, 尤业明. 岩溶区青冈栎整树蒸腾的季节变化. 应用生态学报, 2009, 20(2): 256- 264.
[18] 刘延惠, 舒德远, 崔迎春, 侯贻菊, 赵文君, 丁访军. 茂兰喀斯特森林亚优势种短萼海桐树干液流特征及其环境因子响应. 水土保持学报, 2016, 30(5): 205- 211.
[19] 朱守谦. 喀斯特森林生态研究(Ⅱ). 贵阳: 贵州科技出版社, 1997.
[20] 俞国松, 王世杰, 容丽, 冉景丞. 茂兰喀斯特森林主要演替群落的凋落物动态. 植物生态学报, 2011, 35(10): 1019- 1028.
[21] 胡蕖, 魏鲁明, 杨成华, 崔迎春, 丁访军. 茂兰喀斯特峰丛中部常绿落叶阔叶混交林群落特征及多样性研究. 福建林业科技, 2009, 36(1): 43- 48.
[22] Granier A. A new method of sap flow measurement in tree stems. Annales Des Sciences Forestieres, 1985, 42(2): 193- 200.
[23] 陈立欣, 张志强, 李湛东, 张文娟, 张晓放, 董克宇, 王国玉. 大连4种城市绿化乔木树种夜间液流活动特征. 植物生态学报, 2010, 34(5):535- 546.
[24] 李海涛, 向乐, 夏军, 林耀明, 梁涛. 应用热扩散技术对亚热带红壤区湿地松人工林树干边材液流的研究. 林业科学, 2006, 42(10): 31- 38.
[25] 王华, 欧阳志云, 郑华, 王效科, 倪永明, 任玉芬. 北京城区常见树种生长季树干液流的时滞特征. 应用生态学报, 2009, 20(9): 2111- 2117.
[26] 杜家菊, 陈志伟. 使用SPSS线性回归实现通径分析的方法. 生物学通报, 2010, 45(2): 4- 6.
[27] 曹文强, 韩海荣, 马钦彦, 康峰峰, 蔺琛. 山西太岳山辽东栎夏季树干液流通量研究. 林业科学, 2004, 40(2): 174- 177.
[28] 刘华, 佘春燕, 白志强, 李倩, 刘端, 韩燕梁. 不同径级的西伯利亚红松树干液流及蒸腾耗水特征的差异. 西北植物学报, 2016, 36(2): 390- 397.
[29] 凡超, 邱燕萍, 李志强, 李建光, 张邦跃, 袁沛元. 荔枝树干液流速率与气象因子的关系. 生态学报, 2014, 34(9): 2401- 2410.
[30] 隋旭红, 张建军, 文万荣. 晋西黄土区辽东栎、山杨树干液流比较研究. 生态学报, 2011, 31(16): 4791- 4798.
[31] 徐先英, 孙保平, 丁国栋, 郭树江, 柴成武. 干旱荒漠区典型固沙灌木液流动态变化及其对环境因子的响应. 生态学报, 2008, 28(3): 895- 905.
[32] 程静, 欧阳旭, 黄德卫, 刘世忠, 张德强, 李跃林. 鼎湖山针阔叶混交林4种优势树种树干液流特征. 生态学报, 2015, 35(12): 4097- 4104.
[33] 王艳兵, 德永军, 熊伟, 王彦辉, 李振华, 刘千. 华北落叶松夜间树干液流特征及生长季补水格局. 生态学报, 2013, 33(5): 1375- 1385.
[34] 赵仲辉, 康文星, 田大伦, 项文化, 闫文德. 湖南会同杉木液流变化及其与环境因子的关系. 林业科学, 2009, 45(7): 127- 132.
[35] 熊伟, 王彦辉, 徐德应. 宁南山区华北落叶松人工林蒸腾耗水规律及其对环境因子的响应. 林业科学, 2003, 29(2): 1- 7.
[36] 孙鹏森, 马履一, 王小平, 翟明普. 油松树干液流的时空变异性研究. 北京林业大学学报, 2000, 22(5): 1- 6.
[37] 涂洁, 刘琪璟, 李海涛, 林耀明. 江西千烟洲湿地松生长旺季树干液流动态及影响因素分析. 林业科学, 2008, 44(1): 46- 51.
[38] 马玲, 赵平, 饶兴权, 蔡锡安, 曾小平, 陆平. 马占相思树干液流特征及其与环境因子的关系. 生态学报, 2005, 25(9): 2145- 2151.
[39] 张晓艳, 褚建民, 孟平, 姚增旺, 王鹤松, 李得禄, 姜生秀. 民勤绿洲荒漠过渡带梭梭(Haloxylonammodendron(C. A. Mey) Bunge)树干液流特征及其对环境因子的响应. 生态学报, 2017, 37(5): 1525-1536
[40] 张建国, 久米朋宣, 大规恭一, 山中典和, 杜盛. 黄土高原半干旱区辽东栎的树干液流动态. 林业科学, 2011, 47(4): 63- 69.
[41] 陈宝强, 张建军, 张艳婷, 田宁宁. 晋西黄土区辽东栎和山杨树干液流对环境因子的响应. 应用生态学报, 2016, 27(3): 746- 754.
[42] 刘春鹏, 朱美秋, 薛帅, 马长明, 支恩波, 李向军. 干旱条件下荆条树干液流对不同天气的响应. 福建林业科技, 2014, 41(1): 34- 39.
CharacteristicsofsapflowandcorrelationanalysiswithenvironmentalfactorsofAcerwangchiiinthekarstarea
WU Peng1,2, YANG Wenbin1, CUI Yingchun2, ZHAO Wenjun2, SHU Deyuan2, HOU Yiju2, DING Fangjun2,*
1InstituteofDesertificationStudies,ChineseAcademyofForestry,Beijing100091,China2GuizhouAcademyofForestry,Guiyang550005,China
The Thermal Dissipation Probe (TDP) method was used to conduct long-term continuous observations from February 2012 to March 2014 on the sap flow velocity ofAcerwangchiiin karst regions. In addition, environmental factors including air temperature (Ta), relative humidity (RH), solar radiation (Solar), wind speed (WS), rainfall, soil water content (SWC), and the characteristics ofA.wangchiisap flow were investigated, and correlations between environmental factors and different temporal scales were determined. The findings revealed the following: 1) sap flow velocity in relation to different weather conditions, in descending order, was sunny, cloudy, and rainy days, and the sap flow was generally high in the daytime and low at night; 2) the average amount of sap flow was 5.08 kg/d, with the sap flow quantity varying in different seasons, in descending order of (8.38 ± 5.32) kg/d in summer, (5.16 ± 3.99) kg/d in autumn, (4.86 ± 3.77) kg/d in spring, and (1.94 ± 1.40) kg/d in winter; 3) the monthly sap flow was 153.64 kg and the annual sap flow was 1838.40 kg, on average; 4) on the hourly scale, solar radiation over 1 day was the main environmental factor influencing sap flow regardless of the weather conditions, while the influence of the other environmental factors on sap flow varied in the daytime, at night, and in different seasons and rainfalls; and 5) over 1 year: on the hourly scale, 63.50% of the sap flow could be interpreted jointly by the environmental factors including solar radiation,Ta, RH, SWC10cmand WS. On the daily scale, 68.50% of the sap flow could be interpreted jointly by the environmental factors including solar radiation,Ta, SWC10cmand rainfall. On the monthly scale, the sap flow quantity could be interpreted with the single environmental factor,Ta, which contributed 74.80% to the total. With the time scale reducing, the more environmental factors were involved in the regression equation, the lesser interpretation of sap flow quantity (R2) would be. In comparison to similar findings from other regions, the impact of environment factors on sap flow diverged greatly. However, on temporal scales, solar radiation or PAR andTawere the two major factors that affected sap flow, and no environmental factor contributed in the interpretation of sap flow on rainy days, which is consistent with the present study. Therefore, estimates of the impact of environmental factors on sap flow on rainy days deviate, and particular attention should be paid to the role of soil water content.
karst;Acerwangchii; sap flow; environmental factors; different temporal scale
黔科合人字(2011)12号;黔科合人才团队[2014]4004号;黔林科合J字〔2015〕14号;黔科合SY字〔2014〕3039号;黔科合院士站〔2014〕4006;黔林科合J字〔2014〕09号
2016- 09- 25; < class="emphasis_bold">网络出版日期
日期:2017- 07- 12
*通讯作者Corresponding author.E-mail: ding3920034@163.com
10.5846/stxb201609251934
吴鹏, 杨文斌, 崔迎春, 赵文君, 舒德远, 侯贻菊, 丁访军.喀斯特区天峨槭(Acerwangchii)树干液流特征及其与环境因子的相关分析.生态学报,2017,37(22):7552- 7567.
Wu P, Yang W B, Cui Y C, Zhao W J, Shu D Y, Hou Y J, Ding F J.Characteristics of sap flow and correlation analysis with environmental factors ofAcerwangchiiin the karst area.Acta Ecologica Sinica,2017,37(22):7552- 7567.
猜你喜欢
疯狂英语·新读写(2021年8期)2021-11-05 08:44:28
阅读(低年级)(2021年2期)2021-04-08 02:16:27
启蒙(3-7岁)(2020年9期)2020-09-30 05:45:24
小学生导刊(2018年22期)2018-08-21 01:47:48
作文小学中年级(2018年6期)2018-06-28 05:51:22
好孩子画报(2017年1期)2017-04-06 17:44:51
电源技术(2017年1期)2017-03-20 13:38:00
电源技术(2015年7期)2015-08-22 08:48:36
健康必读(2014年1期)2014-06-23 14:58:22
中国质量与标准导报(2014年5期)2014-02-28 22:23:14
