
饥饿对圆口铜鱼Coreius guichenoti仔鱼早期发育的影响
2018-01-02 12:07曲焕韬刘勇胡美洪倪勇吕小林鲁雪报
水产学杂志 2017年6期
曲焕韬,刘勇,胡美洪,倪勇,吕小林,鲁雪报
(中国长江三峡集团公司中华鲟研究所,三峡工程鱼类资源保护湖北省重点实验室,湖北 宜昌 443100)
饥饿对圆口铜鱼Coreius guichenoti仔鱼早期发育的影响
曲焕韬,刘勇,胡美洪,倪勇,吕小林,鲁雪报
(中国长江三峡集团公司中华鲟研究所,三峡工程鱼类资源保护湖北省重点实验室,湖北 宜昌 443100)
在水温(22.0±1.0)℃条件下,研究饥饿状态下圆口铜鱼Coreius guichenoti仔鱼生长、初次摄食率、不可逆点(PNR)以及摄食节律。结果表明:圆口铜鱼初孵仔鱼全长(7.50±0.01)mm,卵黄囊体积(3.36±0.11)mm3,摄食组仔鱼全长生长呈线性增加,饥饿组仔鱼生长呈先增长后下降趋势。早期仔鱼(5日龄)和晚期仔鱼(15日龄)的摄食节律相似,高峰在凌晨6:00和黄昏18:00,属于典型的晨昏摄食类型。1~3日龄仔鱼为内源性营养期;4日龄仔鱼开口摄食,进入混合营养期,初次摄食率约为25.1%;5日龄卵黄囊耗尽,进入外源营养期;7日龄仔鱼摄食率达到最高水平为100%,最高摄食率维持6d,PNR期为13~14日龄。在本实验水温下,圆口铜鱼仔鱼最佳投喂时间为4~6日龄。
圆口铜鱼;饥饿;初次摄食率;不可逆点
鱼类早期发育阶段运动、摄食等组织器官发育尚不健全,抵御饥饿的能力较差,极易遭受饥饿威胁。因此,通常认为饥饿是初次摄食期仔鱼死亡的重要原因之一[1]。Blaxter和Hempel[2](1963年)首先从生态学角度提出初次摄食仔鱼饥饿“不可逆点”(point of no return,PNR)的概念:初次摄食仔鱼达到耐受饥饿的时间临界点时,虽然可存活一段时间,但一半个体已体质虚弱,不可能再恢复摄食能力。自“不可逆点”概念提出以来,国内外学者以此作为衡量初次摄食期仔鱼饥饿耐受能力的标准,围绕早期仔鱼饥饿发生过程及其影响机制进行了广泛深入的研究,积累了大量鱼类早期发育生物学基础数据和资料[3-7]。
圆口铜鱼Coreiusguichenoti隶属鲤科Cyprinidae、鮈亚科Gotiongiae、铜鱼属Coreius,俗称水密子、肥沱、麻花等,广泛分布于金沙江中下游、长江上游、雅砻江干流以及乌江下游等水系,为长江上游具有代表性的特有鱼类和重要经济鱼类[8-10]。圆口铜鱼是一种产漂流性卵的河道洄游性鱼类。近年来随着长江上游、金沙江中下游及雅砻江干流下游梯级水电开发的逐步实施,圆口铜鱼栖息生境、繁殖水文条件以及繁殖洄游通道等均遭受不同程度的干扰和破坏,野生圆口铜鱼资源急剧衰退[11-15]。圆口铜鱼物种保护问题引起了广泛关注,已开展一系列物种保护工作。前期研究主要集中在野生种群资源量调查、食性分析、繁殖生物学以及病害学等方面[16-19],有关其仔鱼摄食习性的研究尚未见报道。本实验通过观察中华鲟研究所自主繁殖的圆口铜鱼早期仔鱼发育过程中卵黄囊消耗进程、开口摄食时间、饥饿不可逆点以及摄食节律等,旨在全面了解早期圆口铜鱼的摄食特点,掌握摄食、生长发育的相关参数,为规模化苗种培育中科学合理投喂提供参考。
1 材料与方法
1.1 材料
实验于2017年6月在金沙江溪洛渡向家坝水电站珍稀特有鱼类增殖放流站开展,实验用圆口铜鱼仔鱼为中华鲟研究所繁殖、孵化获得,繁殖用的亲本均为人工驯化的野生鱼。
1.2 方法
1.2.1 饥饿胁迫对仔鱼早期生长发育影响的观测
圆口铜鱼仔鱼孵化后,随机挑选2 000尾体态正常、活力较好的初孵仔鱼,分为饥饿组和正常投喂的对照组,每组1 000尾,分别置于容量约80L的水族缸内。实验用水为向家坝水电站坝下江段水,经沉淀、过滤及曝气处理,水温(22.0±1.0)℃,每天适量换水。饥饿组始终不投喂,直至100%仔鱼死亡,正常投喂组自3日龄卵黄囊基本耗尽开始投喂轮虫,密度 20~30ind./mL。
实验期间,每天取饥饿组和对照组仔鱼各20尾,分别在Olympus SZ61型体视显微镜下用数码显微摄像系统测量仔鱼全长、卵黄囊长径、卵黄囊短径以及肛前长等常规生物学指标,精确度达0.01mm。
卵黄囊体积计算公式:卵黄囊容量=4/3π×(r/2)2×R/2,式中r为卵黄囊短径,R为卵黄囊长径。
1.2.2 初次摄食率和不可逆点(PNR)的测定
3日龄开始每天下午15:00从饥饿组中随机取20尾,转入有效容量1 000mL的烧杯中,微充气,投喂轮虫开口饵料,密度20~30ind./mL,2h后将仔鱼转出。经MS-222麻醉后在体视显微镜下逐一观察仔鱼摄食情况,计算初次摄食率。
初次摄食率=x/z×100%,式中x为肠胃中含有轮虫的仔鱼数,z为实验仔鱼总数。
圆口铜鱼仔鱼的PNR用日龄表示,参照Yin和Blaxter[3]的方法进行测量,即:每天测定饥饿组圆口铜鱼仔鱼的初次摄食率,当测定的仔鱼摄食率降至最高初次摄食率50%时,即为PNR的时间。
1.2.3 摄食节律
依据殷名称[20]的方法,将仔鱼分为早期仔鱼和晚期仔鱼。早期仔鱼定义为初孵仔鱼至卵黄囊完全消失的阶段;晚期仔鱼为卵黄囊耗尽开始至各鳍鳍条发育完整,鳞片开始形成。本实验观察正常投喂组5日龄和15日龄仔鱼的摄食节律。自上午6:00至翌日上午6:00,每4h取样1次,即6:00、10:00、14:00、18:00、22:00、翌日 2:00 以及翌日 6:00,每组设3个平行组,每组选仔鱼15尾,实验仔鱼取自水族缸内正常投喂组。麻醉处理后,在体视显微镜下观测消化道饱满程度,解剖肠道,计数每尾仔鱼摄食轮虫数量,轮虫平均湿重按2.5μg/个计算。
1.3 数据分析
实验数据均用平均值±标准差(Mean±SD)表示,利用SPSS18.0进行单因素方差(ANOVA)分析。
2 结果与分析
2.1 饥饿组和对照组仔鱼早期生长发育
圆口铜鱼初孵仔鱼全长(7.50±0.01)mm,肛前长(5.18±0.02)mm,卵黄囊长径为(4.60±0.10)mm,卵黄囊短径为(1.18±0.03)mm,卵黄囊体积为(3.36±0.11)mm3。对照组不同日龄仔鱼各项生长指标(全长、肛前长、卵黄囊长径、短径以及卵黄囊体积)详见表1。1~3日龄仔鱼为内源性营养期,肛门和口为封闭状态,鱼苗生长发育主要依靠卵黄囊提供营养。1日龄仔鱼卵黄囊消耗速度最快,体积减小为初孵仔鱼卵黄囊的44.05%;2日龄卵黄囊进一步吸收,体积为初始体积的20.54%左右;至4日龄进入混合营养期,消化系统和运动器官快速发育,口窝形成,能够开合,仔鱼已初步具备摄食消化能力;5日龄以上仔鱼,卵黄囊完全耗尽,进入外源性营养期。
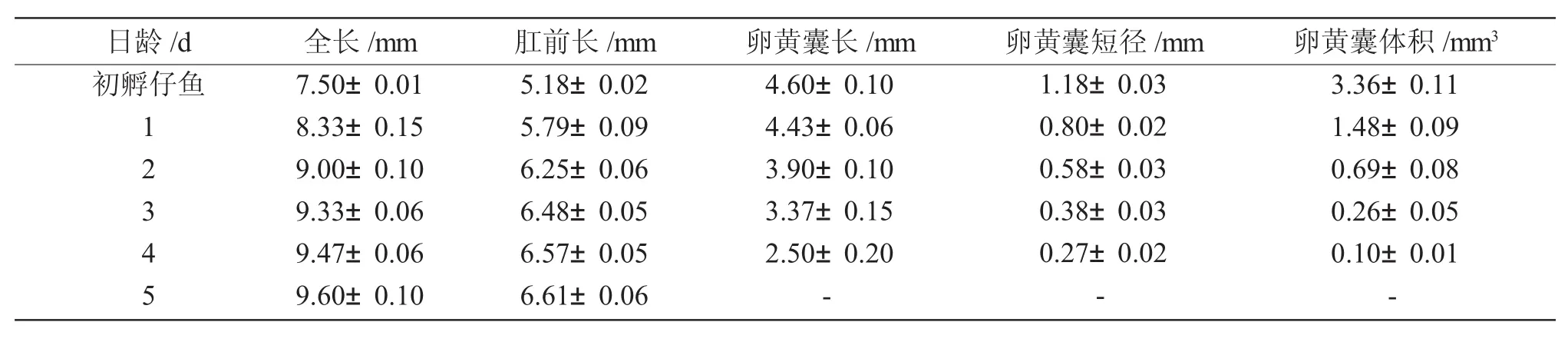
表1 圆口铜鱼对照组仔鱼全长和卵黄囊的变化Tab.1 Changes in total length and volume of yolk sac of Coreius guichenoti larvae in the control group
饥饿组和对照组仔鱼全长变化情况详见图1。由图1可知,饥饿仔鱼全长生长可分为3个阶段:1)初孵仔鱼至3日龄仔鱼为快速生长阶段,全长特定增长率约6.03%;2)4~7日龄仔鱼全长增速明显放缓,全长特定增长率约1.91%,7日龄仔鱼全长达到最大值(10.03±0.15)mm;3)饥饿组 8~15 日龄仔鱼全长为负增长,15日龄仔鱼全长仅为(9.39±0.15)mm。线性拟合分析表明,对照组仔鱼全长始终保持线性增长,体全长(L)与日龄(d)符合线性关系式:L=0.4832d+7.6282(R2=0.9908);饥饿组仔鱼全长呈“升高-趋稳-降低”的变化过程,全长(L)与日龄(d)的关系式为:L=-0.026d2+0.4712d+7.8979(R2=0.925)。
2.2 饥饿仔鱼初次摄食率及PNR
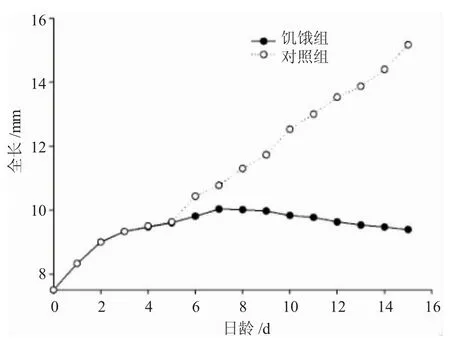
图1 圆口铜鱼仔鱼饥饿组和对照组全长生长比较Fig.1 The comparison of growth in total length of Coreius guichenoti larvae between control and starved groups
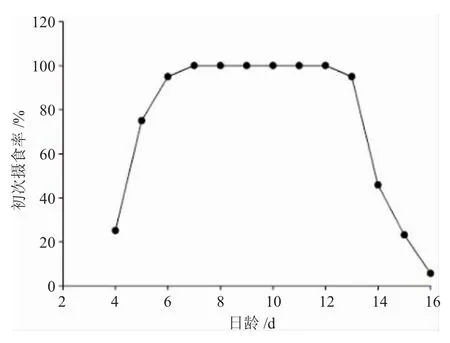
图2 圆口铜鱼饥饿仔鱼的初次摄食率Fig.2 The initial feeding rate of starved Coreius guichenoti larvae
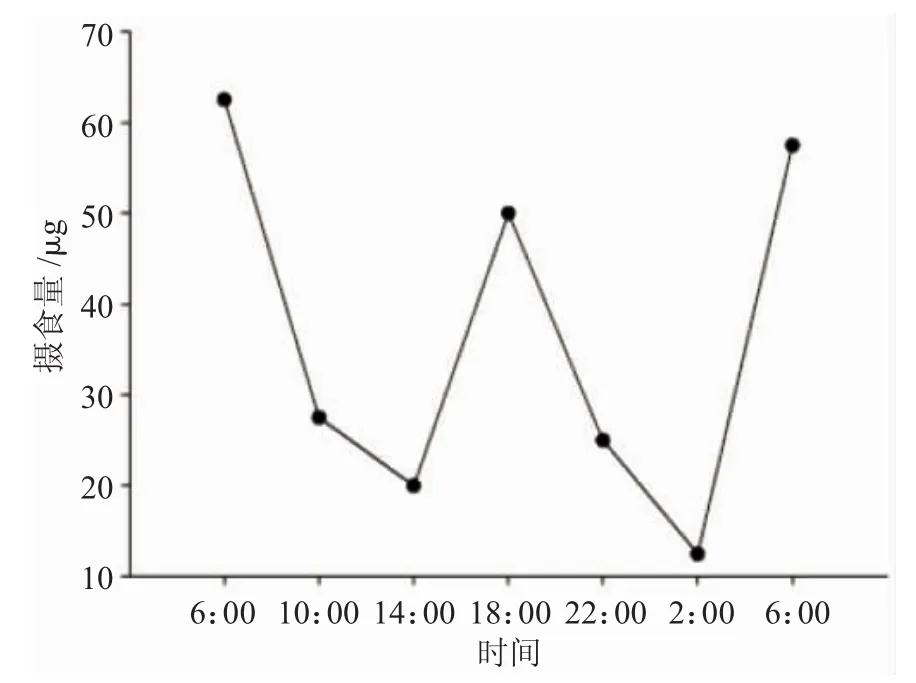
图3 5日龄圆口铜鱼仔鱼摄食节律Fig.3 The feeding rhythm of 5-day-old Coreius guichenoti larvae
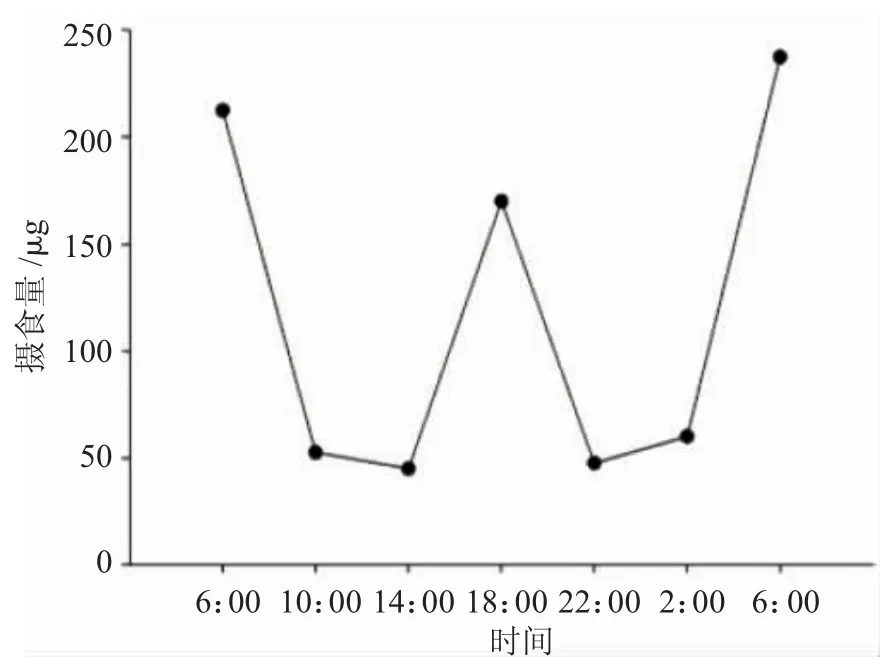
图4 15日龄圆口铜鱼仔鱼摄食节律Fig.4 The feeding rhythm of 15-day-old Coreius guichenoti larvae
饥饿胁迫条件下圆口铜鱼仔鱼初次摄食率见图2。在水温(22.0±1.0)℃下,4日龄仔鱼卵黄囊已大量消耗,残余量约为初始体积的2.98%。此时仔鱼运动和摄食器官已初步发育,肛门与外界相通,具备一定的摄食能力,仔鱼进入混合营养期,初次摄食率约为25.10%;此后,随着日龄增加,初次摄食率逐步上升,至7日龄初次摄食率首次达到最高水平100%,摄食率在最高水平维持6d后开始下降,14日龄仔鱼摄食率约为45.80%,15日龄时已极度衰弱,难以维持平衡,摄食能力显著下降;17日龄鱼苗消瘦弯曲,完全丧失摄食能力,全部死亡。综上所述,在本实验条件下14日龄仔鱼初次摄食率降至最高摄食率的50%以下,因此,13~14日龄仔鱼即进入PNR期。圆口铜鱼仔鱼从初次摄食到PNR期大约经历 10~11d。
2.3 早期仔鱼摄食节律
由图3和图4可知,圆口铜鱼仔鱼昼夜摄食节律明显。5日龄仔鱼在1昼夜内出现3次摄食高峰,分别为6:00、18:00以及翌日6:00,其中,凌晨6:00摄食量最大,每尾仔鱼摄食量约为57~62μg;摄食低谷为夜间2:00左右,摄食量仅为12μg左右。15日龄仔鱼摄食节律与5日龄基本类似,但摄食量明显增加,最高摄食量达到237μg/尾。
3 讨论
3.1 饥饿对仔鱼早期生长发育的影响
Farris根据生长速率差异将卵黄囊仔鱼发育过程划分为3个时期:初孵仔鱼的快速生长期、卵黄囊消失前后的慢速生长期以及不能建立外源营养时的负增长期[21]。本实验中,圆口铜鱼饥饿仔鱼的生长规律基本符合上述生长期的描述。初孵仔鱼至3日龄,仔鱼依靠卵黄囊提供营养,处于快速生长的内源性营养阶段;4~7日龄,卵黄囊基本耗尽,处于内源性营养向外源营养过渡的阶段,增长速率开始放缓;7日龄以上鱼苗因饥饿进展出现负增长现象,这是卵黄囊消耗完毕后,未能及时建立外源性营养的必然结果。关于仔鱼的负增长现象,一般认为是骨骼系统尚未发育完善的仔鱼为保障活动耗能,提高摄食和存活机会的一种生存策略[22,23]。
3.2 圆口铜鱼仔鱼的摄食特性
摄食节律体现摄食量随时间变化的规律,研究鱼类摄食节律是了解鱼摄食行为的重要途径。不同鱼类摄食节律不同,同种类在不同的生活环境和发育阶段,其摄食节律也可能存在一定的差异[24]。Hemifan根据鱼类摄食节律特点,将鱼类的摄食分为白天摄食、晚上摄食、晨昏摄食以及无明显摄食规律4种类型[25]。本实验中,圆口铜鱼早期仔鱼(5日龄)在1昼夜内在凌晨6:00、18:00及次日6:00出现3次摄食高峰,凌晨摄食量略高于黄昏,最高摄食量达到约62μg/尾;晚期仔鱼(15日龄)摄食节律与早期仔鱼基本一致,但摄食强度明显增加,最大摄食量达237μg/尾。根据Hemifan的分类方法可判定圆口铜鱼属于晨昏摄食类型,即白天摄食量相对较小,凌晨和黄昏游动活跃,摄食强度较高。初步分析,圆口铜鱼这种摄食节律的形成可能与其营底栖生活的习性密切相关,在自然界江河深水区,底层光线较弱,水体透明度低,鱼类摄食主要依靠化学感觉、特化夜视觉以及侧线机械感觉。因此,圆口铜鱼仔鱼的摄食特性也可视为其与环境协同进化的生存策略的重要体现。
3.3 初次摄食率、不可逆点及饥饿耐受力
鱼类的初次摄食时间不仅由种属遗传特性、器官发育、卵黄囊大小以及吸收速度等内源性因素决定,也受培育水温、饵料种类及密度等外界环境因子的影响[26-28]。在(22.0±1.0)℃条件下,圆口铜鱼仔鱼4日龄开始初次摄食,这与(21±1)℃下花鲈Lateolabrax maculatus[29]4日龄和(21±1)℃下斑鳜Siniperca scherzeri[1]2日龄开口摄食时间基本接近,但较10~12℃条件下哲罗鱼Hucho taimen[30]21日龄开口摄食明显提前。上述开口摄食时间的差异主要是不同鱼类种群遗传属性和培育水温差异综合作用的结果。
鱼类的摄食能力可用摄食率与最高初次摄食率持续时间来判断,而PNR主要用于判断鱼类耐受饥饿的能力。抵达PNR的时间越长,鱼类耐受饥饿能力越强,反之则耐受饥饿的能力越弱[31]。本实验研究表明:圆口铜鱼仔鱼初次摄食率约为25.10%,明显高于斑鳜[1]的5%、花鲈[29]的10%以及大泷六线鱼Hexagrammos otakii 15%;7日龄仔鱼摄食率达到100%,最高摄食率维持时间约6d,与瓦氏黄颡鱼Pelteobagrus vachelli[32]摄食能力基本一致,但明显高于黄鲷Taius tumifrons[33]最高摄食率75%,持续1d。这可能与仔鱼摄食效率、器官发育进程以及饵料密度等因素相关。
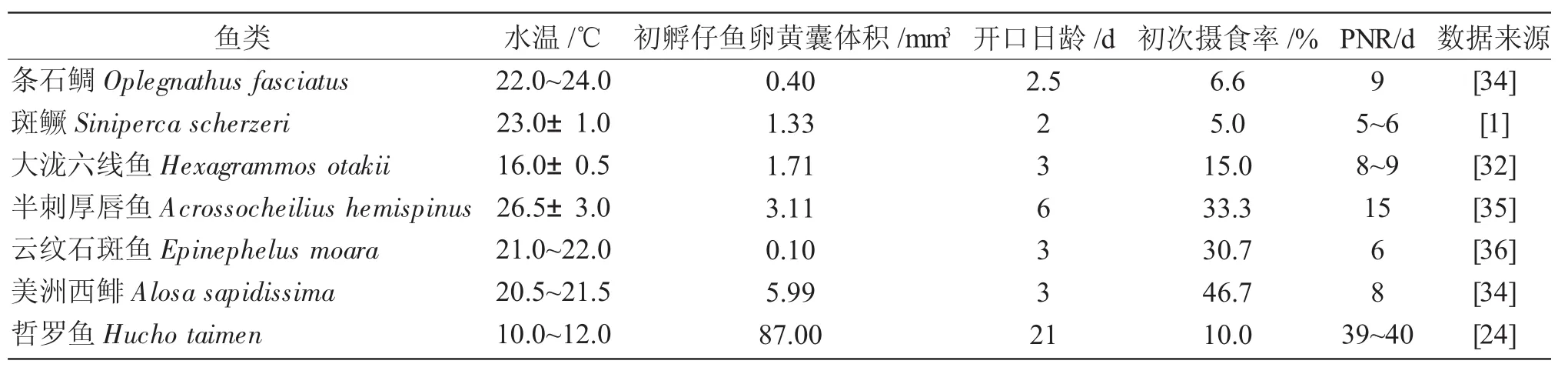
表2 不同鱼类卵黄囊体积、初次摄食率以及PNRTab.2 The volume of yolk sac,initial feeding rate and PNR in different fishes
从初次摄食到PNR是鱼类构建外源营养的关键时期,若仔鱼在混合营养期内没有及时建立外源性营养,则将遭受进展性饥饿而大量死亡。与大部分鱼类仔鱼相比(表2),圆口铜鱼仔鱼的PNR期明显偏长,约为13~14d,具有较强的摄食和耐受饥饿的能力,这可提高自然状态下仔鱼存活的概率,保障其种群数量稳定和下一世代种质资源补充,为人工驯化摄食创造有利条件,显著提高开口阶段仔鱼的存活率。
综上所述,在水温(22.0±1.0)℃培育条件下,4日龄圆口铜鱼仔鱼开始摄食,7日龄摄食率达到最高水平,13~14日龄仔鱼进入不可逆时期。在养殖生产中,应在仔鱼达到最高摄食水平前即开始投喂,以便仔鱼积累捕食经验,建立巡游模式。本研究表明,在本实验温度条件下,圆口铜鱼仔鱼最适初次投喂时间为孵化后的4~6d。
[1]王茂元.斑鳜仔鱼饥饿实验及不可逆点的确定[J].广东海洋大学学报,2015,35(4):99-103.
[2]Blaxter J H S and Hempel G.The influence of egg size on herring larvae (Clupea harengus L.)[J].ICES Journal of Marine Science,1963,28(2):211-240.
[3]Yin MC and Blaxter J H S.Morphological changes during growth and starvation of larval cod(Gadus morhua L.)and flounder(Platichthys flesus L.)[J].Journal of Experimental Marine Biologyand Ecology,1986,104(1-3):215-228.
[4]万瑞景,李显森,庄志猛,等.鳀鱼仔鱼饥饿试验及不可逆点的确定[J].水产学报,2004,28(1):79-83.
[5]黄良敏,谢仰杰,张光后,等.延迟投饵对浅色黄姑鱼仔鱼摄食、生长和存活的影响[J].大连水产学院学报,2005,20(4):300-303.
[6]殷名称,哈维S M,凯克J C A.江鲽在卵和卵黄囊期仔鱼发育阶段生化成分的变化[J].动物学报,1993,39(3):272-279.
[7]高露姣,陈立侨,宋兵,等.饥饿对杂交鲟消化系统发育的影响[J].上海水产大学学报,2006,15(4):442-447.
[8]吴江,吴明森.金沙江的鱼类区系[J].四川动物,1990,9(3):23-26.
[9]施白南,熊天寿.三十年来四川省江河鱼类资源调查概况(1950-1980)[J].四川水产,1980(3):1-22.
[10]曹发君,余先元,周键,等.攀枝花市雅砻江鱼类资源调查[J].四川动物,1996,15(3):113-115.
[11]黄木桂,邱林顺,陈大庆,等.长江三峡水利枢纽与库区渔业资源[J].淡水渔业,1998,28(5):7-9.
[12]邱林顺,陈大庆,黄木桂,等.三峡工程截流前长江渔业资源状况初析[J].淡水渔业,1998,28(2):3-6.
[13]唐会元,杨志,高少波,等.金沙江中游圆口铜鱼早期资源现状[J].四川动物,2012,31(3):416-421.
[14]张志英,袁野.溪洛渡水利工程对长江上游珍稀特有鱼类的影响探讨[J].淡水渔业,2001,31(1):62-63.
[15]高少波,唐会元,陈胜,等.金沙江一期工程对保护区圆口铜鱼早期资源补充的影响[J].水生态学杂志,2015,36(2):6-10.
[16]刘飞,但胜国,王剑伟,等.长江上游圆口铜鱼的食性分析[J].水生生物学报,2012,36(6):1081-1086.
[17]陈春娜,黄颖颖.圆口铜鱼的生物学及病害学等研究现状[J].水产科学,2009,28(11):706-709.
[18]张其中,陈达丽.患小瓜虫病圆口铜鱼的组织病理学观察[J].西南师范大学学报:自然科学版,2005,30(6):1112-1115.
[19]杨志,乔晔,张轶超,等.长江中上游圆口铜鱼的种群死亡特征及其物种保护[J].水生态学杂志,2009,2(2):50-55.
[20]殷名称.鱼类早期生活史研究与其进展[J].水产学报,1991,15(4):348-358.
[21]Farris D A.Change in the early rates of four larval marine fishes[J].Limnologyand Oceangraphy,1959,4(1):29-36.
[22]邹记兴,向文州,胡超群,等.点带石斑鱼仔鱼营养转换期的摄食与生长[J].高技术通讯,2003,13(5):87-91.
[23]殷名称.北海鯡卵黄囊期仔鱼的摄食能力和生长[J].海洋与湖沼,1991,22(6):554-559.
[24]高小强,洪磊,刘志峰,等.美洲西鲱仔鱼不可逆点及仔、稚鱼摄食特性研究[J].水产学报,2015,39(3):392-400.
[25]Helfman G S.Fish behavior by day,night and twilight[M]//Pitchered T J.The Behavior of Teleost Fishes.Baltimore Maryland:The Johns Hopkins University Press,1986:366-387.
[26]马旭洲,王武,甘炼,等.延迟投饵对瓦氏黄颡鱼仔鱼存活、摄食和生长的影响[J].水产学报,2006,30(3):323-328.
[27]Houde E D.Fish earlylife dynamics and recruitment variability[J].Amer Fish Symp,1987,2:17-29.
[28]殷名称.鱼类仔鱼期的摄食和生长[J].水产学报,1995,19(4):335-342.
[29]王晓龙,温海深,张美昭,等.花鲈初孵仔鱼饥饿不可逆点的确定及摄食节律研究[J].中国海洋大学学报,2017,47(5):57-64.
[30]张永泉,尹家胜,杜佳,等.哲罗鱼仔鱼饥饿实验及不可逆生长点的确定[J].水生生物学报,2009,33(5):945-950.
[31]殷名称.鱼类早期生活史阶段的自然死亡[J].水生生物学报,1996,20(4):363-372.
[32]菅玉霞,房慧,张少春,等.大泷六线鱼仔鱼饥饿试验及不可逆点研究[J].海洋科学,2014,38(3):111-115.
[33]夏连军,施兆鸿,陆建学.黄鲷仔鱼饥饿试验及不可逆点的确定[J].海洋渔业,2004,26(4):286-290.
[34]彭志兰,柳敏海,罗海忠,等.条石鲷仔鱼饥饿试验及不可逆点的确定[J].水产科学,2010,29(3):152-155.
[35]秦志清.饥饿对半刺厚唇鱼(Acrossocheilius hemispinus)仔鱼早期发育的主要影响[J].集美大学学报:自然科学版,2015,20(4):241-248.
[36]于欢欢,陈超,张廷廷,等.饥饿对云纹石斑鱼卵黄囊期仔鱼摄食和生长的影响[J].渔业科学进展,2015,36(6):37-42.
Effects of Starvation on Feeding,Growth and Development of Coreius guichenoti Larvae
QU Huan-tao,LIU Yong,HU Mei-hong,NI Yong,LV Xiao-lin,LU Xue-bao
(Hubei Key Laboratory of Three Gorges Projects for Conservation of Fishes,Institute of Chinese Sturgeon,China Three Gorges Corporation,Yichang 443100,China)
The effects of starvation on point of no return (PNR),growth,initial feeding rate and feeding rhythm were studied in Coreius guichenoti larvae at water temperature of(22.0 ± 1.0)℃.The results showed that newly-hatched larvae had total length of(7.50± 0.01)mm,with yolk sac of(3.36 ± 0.11)mm3in volume.The larvae in control group showed continuous increase in total length,while the starving larvae had first increase and then decrease in total length.A comparable trend of feeding rhythm was observed in early and late stage larvae,with feeding peaks at 6:00 and 18:00,indicating that Coreius guichenoti is typical morning-dusk feeder.The 1~3 day-old larvae showed endogenous nutrition,and the 4 day-old larvae had initial feeding,showing mixed nutrition,with initial feeding rate of 25.1%.The yolk sac was completely exhausted and the exogenous nutrients were found in 5 day old larvae.The maximum initial feeding rate(100%)appeared at the age of 7 days,and lasted for 6 days,and the PNR was observed in 13~14 day-old larvae.The optimal initial feeding time is suggested to be carried out from 4 d to 6 d after hatching.
Coreius guichenoti;starvation;initial feeding rate;point of no return
S965.199
A
1005-3832(2017)06-0024-06
2017-07-10
曲焕韬(1984-),男,硕士研究生,工程师,从事长江珍稀特有鱼类保护生物学研究.Email:quhuantao35@163.com
猜你喜欢
农村科学实验(2022年16期)2022-11-19
空间科学学报(2021年1期)2021-05-22
当代水产(2021年1期)2021-03-19
四川蚕业(2021年1期)2021-02-12
水产科学(2020年4期)2020-07-28
当代水产(2019年9期)2019-10-08
中国学校体育(2014年4期)2014-12-06
中国海洋大学学报(自然科学版)(2014年8期)2014-02-28
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28
人生与伴侣·共同关注(2009年36期)2009-01-15
