
稻草覆盖和香根草篱对红壤坡耕地土壤酶活性和微生物群落结构的影响
2017-12-18 03:49成艳红黄欠如武琳黄尚书钟义军孙永明张昆章新亮
中国农业科学 2017年23期
成艳红,黄欠如,武琳,黄尚书,钟义军,孙永明,张昆,章新亮
稻草覆盖和香根草篱对红壤坡耕地土壤酶活性和微生物群落结构的影响
成艳红,黄欠如,武琳,黄尚书,钟义军,孙永明,张昆,章新亮
(江西省红壤研究所/江西省红壤耕地保育重点实验室/国家红壤改良工程研究中心/农业部江西耕地保育科学观测试验站,江西进贤331717)
明确不同生物水土保护措施下土壤有机碳与土壤微生物种类及功能间的关系。依托始于2009水土保护年的红壤坡地不同生物水土保护措施的水土保护效果野外试验,利用磷脂脂肪酸方法(PLFA),研究花生等高种植(CK)、香根草篱+花生(H)、稻草覆盖+花生(M)和稻草覆盖+香根草篱+花生(HM)对土壤微生物量和组成以及参与碳循环土壤酶活性、土壤有机碳、活性有机碳的影响。与CK相比,H、M和HM 3种水土保护措施土壤总有机碳(TOC)、可溶性有机碳(DOC)、土壤微生物量碳(MBC)和微生物熵含量均有增加,香根草篱是增加土壤MBC和微生物熵的主要影响因素。参与碳循环的蔗糖酶(INV)、纤维素酶(CEL)、β-葡糖苷酶(β-Glu)以及微生物量碳酶强度均受到香根草篱的显著影响。PLFA分析表明,与CK相比,H、M和HM显著增加了土壤PLFAs 总量(TB)、细菌(B)和放线菌(ACT)的数量,降低了革兰氏阴性菌(GN)/革兰氏阳性菌(GP)比例,尤以HM处理减少幅度最大。主成分分析表明,H和M处理土壤微生物群落结构相近,与CK差别较大。冗余分析(RDA)表明,土壤TOC、MBC与参与碳循环CEL、INV、β-Glu酶的活性均是影响土壤微生物数量和结构的主要因子。稻草覆盖和香根草篱对红壤坡耕地土壤参与碳循环的酶活性、微生物数量和群落结构有不同影响。
红壤坡耕地;稻草覆盖;香根草篱;土壤酶活性;微生物群落结构
0 引言
【研究意义】红壤坡耕地是中国耕地资源的重要组成部分,该区水热资源丰富,是中国重要粮油生产基地[1]。近年来在降雨、地形等自然因素和耕作、施肥等人为因素的作用下,该区水土流失加剧,土地生产力下降,农业生态系统脆弱,严重制约了区域农业的可持续发展和生态环境的改善[2-3]。因此,探索、研究和实施有效的控制坡耕地水土流失技术,对实现我国资源、环境和人口的协调发展意义重大。【前人研究进展】等高植物篱和秸秆地面覆盖因其成本低、简单实用,生态、经济、社会效益显著,业已成为阻控南方红壤坡地水土流失和恢复重建侵蚀型红壤的有效措施[4-5]。近年来,植物篱和秸秆覆盖等水土保护措施在有效减流减沙,提高土壤养分,改善土壤团粒结构,增强土壤抗侵蚀能力等方面效果显著[6-11]。本研究团队之前研究发现,香根草篱种植5年后,土壤中>0.25 mm粒级机械稳定性和水稳性团聚体含量均显著增加,香根草篱结合稻草覆盖增加了红壤坡耕地土壤及团聚体内有机碳含量[12]。有机碳对土壤团聚体的形成和稳定的重要作用已得到公认[13-14]。土壤微生物及其分泌的酶与土壤养分的生物转化过程紧密相关,是土壤有机碳转化过程的重要调控者,其在形成土壤肥力和稳定土壤团聚结构上有积极作用[15]。土壤团聚结构是制约土壤抗蚀性和抗冲性的重要因子[16]。因此,从水土保持的角度来讲,土壤微生物量及其结构变化一定程度上反映了水土保护措施阻控水土流失的机理和作用方式。【本研究切入点】目前,有关稻草覆盖、植物篱的研究多集中在其对土壤肥力、土壤结构状况和水土保持效益研究,对土壤微生物区系及多样性的研究较少,且大多集中在植物篱、覆盖等单一措施下[17-19],对两种措施联合作用下土壤微生物组成特征及其与土壤碳及碳转化相关土壤酶活性的关系研究鲜有报道。【拟解决的关键问题】本文采用始于2009年的红壤坡地不同生物水土保护措施的水土保护效果野外试验,利用磷脂脂肪酸方法(PLFA)研究香根草篱和稻草覆盖对红壤坡耕地土壤微生物数量及群落组成的影响,结合微生物生物量碳及土壤酶活性变化,探讨不同生物水土保护措施下土壤有机碳与土壤微生物种类及功能间的关系。旨在为探讨侵蚀红壤结构重建的途径,筛选更利于红壤坡地土壤结构改良和碳累积的合理管理模式提供理论依据。
1 材料与方法
1.1 试验区概况
试验区位于江西省进贤县江西省红壤研究所水土保持试验站(116°20′24″E, 28°15′30″N)。该区属中亚热带季风气候,年均降雨量1 537 mm,年蒸发量1 100—1 200 mm,年均气温17.7—18.5℃。土壤为第四纪黏土母质发育的红壤旱地,供试土壤的基本理化性质如下:pH(H2O)5.0,有机质 16.2 g·kg-1,全氮 1.03 g·kg-1,全磷 0.66 g·kg-1,有效磷(Olsen-P)6.8 mg·kg-1,全钾 1.39 g·kg-1。土壤容重1.27 g·cm-3,土壤总孔隙度58.4%,土壤砂粒、粉粒和黏粒的组成分别是16.31%、42.35%、41.35%。
1.2 试验设计
该试验始于2009年,设置等高花生常规耕作(CK)、花生+香根草篱(H)、花生+稻草覆盖(M)、花生+香根草篱+稻草覆盖(HM)4个处理,3次重复,完全随机排列。各处理基础肥力及花生种植方式和农事操作相同。样地坡度10°,小区面积120 m2(24 m×5 m)。供试花生品种为粤油991,种植密度为32 cm×20 cm,花生等高种植,对照小区和秸秆覆盖小区种植72行,草篱小区和草篱+秸秆覆盖小区种植66行(其余6行为草篱)。香根草()篱每隔8 m双行种植,株行距为50 cm×50 cm,试验期间确保香根草定期刈割至30—50 cm。每隔3年施一次石灰,于土壤翻耕后均匀播撒石灰1 875 kg·hm-2,播种前施三元复合肥(N-P2O5-K2O: 15-15-15)416.7 kg·hm-2,钙镁磷肥(CaO≥18%,MgO≥3%,P2O5≥12%)525 kg·hm-2。覆盖稻草处理花生播种后每个小区均匀覆盖干稻草4 500 kg·hm-2。种植制度为一季花生。
1.3 样品采集及测定方法
于2014年9月(花生收获后一个月)在试验小区的上坡位采集距离香根草篱带上部边缘2 m内耕层0—15 cm土样,无篱小区沿等高线采集相同坡位原状土样,每个土样为采样区内10个采样点的混合样。采集的土样剔除可见的动、植物残体和石块等过2 mm筛,在4℃下保存用于土壤微生物PLFA和土壤酶活性、微生物生物量碳和可溶性有机碳测定。取出部分样品风干后磨碎过100目筛,用于测定土壤有机碳含量。
1.3.1 土壤酶活性和有机碳 选取了与土壤碳代谢有密切关系的β-葡(萄)糖苷酶(β-Glucosidase,β-Glu)、多酚氧化酶(polyphenol oxidase,PPO)、纤维素酶(cellulase,CEL)和蔗糖酶(invertase,INV),其酶活性的测定参考关松荫等[20]的方法,其中β-葡糖苷酶采用对硝基酚比色法,以1 g土壤在30℃下培养1 h生成对硝基酚的mg数表示;多酚氧化酶活性采用邻苯三酚显色法,以1 g土壤在30℃下培养2 h分解邻苯三酚的mg数来表示;纤维素酶采用3,5-二硝基水杨酸比色法,以10 g土壤在37℃下培养72 h后生成葡萄糖的mg数表示;蔗糖酶活性采用3,5-二硝基水杨酸比色法,以1 g土壤在37℃下培养24 h后生成葡萄糖的mg数表示。
土壤有机质测定采用浓硫酸-重铬酸钾外加热法;土壤可溶性有机碳采用蒸馏水25℃恒温振荡浸提30 min(水土比为4﹕1)后,以4 000 r/min离心10 min,上清液用0.45 µm 滤膜抽滤, 滤液直接在岛津TOC- VCPH有机碳分析仪上测定;土壤微生物生物量碳采用氯仿熏蒸-0.5 mol·L-1硫酸钾浸提法[21]测定。
1.3.2 土壤微生物中的磷脂脂肪酸(PLFA) 本试验采用修正的Bligh-Dyer[22]方法。(1)提取:称取8 g冷冻烘干土壤,用色谱纯CHCl3、CH3OH、磷酸缓冲液按2﹕1﹕0.8的比例混合浸提液在<25℃下振荡2 h(285 r/min)后离心10 min(3 500 r/min),收集上层离心液,如此反复一次提取后加入磷缓冲液和氯仿各12 mL,于黑暗中静置12 h,吸取下层溶液,在30—32℃水浴锅中用氮气吹干。(2)分离:取1 000 μL CHCl3转移浓缩的磷脂到萃取小柱,向小柱加5mL CHCl3用于洗去中性脂,加2次5 mL丙酮用于洗去糖性脂,用1 mL甲醇清洗萃取小柱底部,收集5 mL甲醇淋洗硅胶小柱溶液,加入4 μL C19:0 内标,在32℃水浴锅中用氮气吹干。(3)甲脂化:用1 mL甲醇﹕甲苯混合液(体积比为1﹕1)和1 mL 0.2 mol·L-1KOH溶解干燥的磷脂样品,在37℃水浴中加热15 min,加入0.3 mL 0.2 mol·L-1的醋酸溶液,2 mL正己烷,2 mL超纯水,低速振荡(120 r/min)10 min,吸取上层正己烷溶液,氮气吹干,加入200μL正己烷冲洗试管,将溶液吸入小柱保存。用Agilent 6850气相色谱仪(FID检测器)测定,仪器条件:色谱柱型号为Agilent 19091B-102E Ultra 25% Phenyl Methyl Siloxane 25.0 m×200 μm×0.33 μm;进样量为2 μL;进样口温度为250℃;载气气体为氦气;恒压模式为22 psi;分流比为100﹕1。
1.4 数据处理与分析
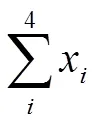
试验结果通过SPSS 17.0 进行单因素方差分析(one-way ANOVA)和相关分析。多重比较选择Duncan极值法,显著性水平为5%。绘图采用Origin8.0软件。为保证结果可靠性,减少误差,仅分析含量高于0.1%的脂肪酸。
2 结果
2.1 土壤有机质和活性有机碳含量变化
土壤有机质含量研究结果表明,与试验前相比,香根草篱(H)和稻草覆盖(M)措施下土壤有机质含量维持在相对稳定水平,且有上升趋势,在二者联合时具有叠加效应,但CK处理降低了土壤总有机碳含量。与CK相比, H、M和HM 3种水土保护措施土壤总有机碳含量分别显著增加7.1%、6.5%和9.9%。但3种水土保护措施间土壤有机质含量无明显差异(图1)。
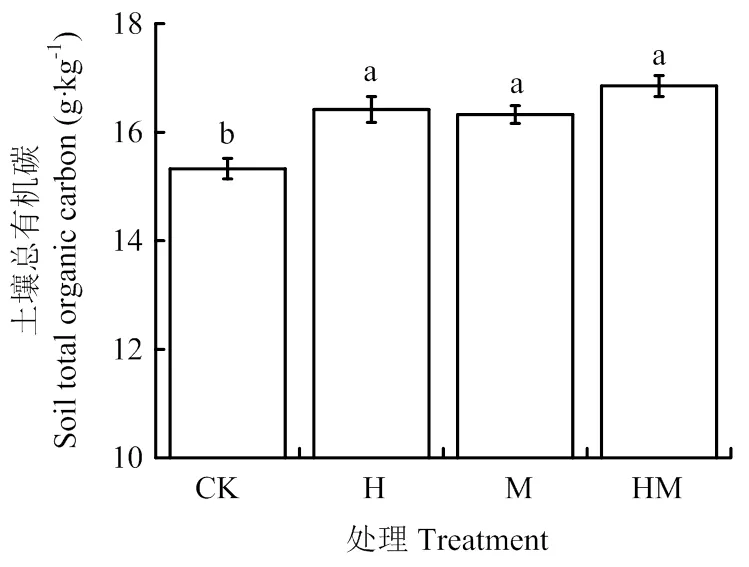
土壤可溶性有机碳(DOC)和土壤微生物生物量碳(MBC)是土壤活性有机碳库的两个重要表征指标,对植物养分供应有最直接作用;微生物熵(微生物量碳与土壤总有机碳的比值,qMB)是评价土壤有机碳动态和质量的有效指标。从图2中可以看出,与CK相比,H、M和HM处理土壤中DOC含量分别提高15.1%、19.6%和13.6%, MBC含量分别提高100.7%、23.3%和160.0%,qMB分别提高87.4%、15.3%和135.9%,其提高幅度远远大于有机碳含量的提高幅度。双因素方差分析结果显示(表1),香根草篱和稻草等外源碳的投入均是增加土壤总有机碳、微生物量碳和微生物熵的主要影响因素。稻草覆盖对DOC含量的影响程度依赖于有无香根草篱种植。
图柱上不同小写字母代表处理间达5%差异显著,平均值±标准误差(n=3)。CK、H、M、HM分别表示花生常规耕作、花生+重根草篱、花生+稻草覆盖、花生+香根草篱+稻草覆盖。下同
2.2 土壤酶活性变化
由表2可知,除了多酚氧化酶(PPO)外,参与碳循环的蔗糖酶(INV)、纤维素酶(CEL)、β-葡糖苷酶(β-Glu)以及微生物生物量碳酶强度均受到香根草篱(H)的显著影响。INV和CEL的趋势一致,H和HM处理均能促进酶活性,分别是CK处理INV活性的1.51倍和1.53倍,CEL活性的1.61倍和1.44倍。与CK相比,H和HM处理均降低了土壤β-Glu活性和微生物生物量碳酶(MBCE)强度,单独香根草篱处理H对β-Glu的降低程度显著高于单独覆盖(M)处理。与CK相比,H和HM处理土壤β-Glu活性分别降低41.9%和73.1%。
图2 香根草篱和稻草覆盖下的土壤可溶性有机碳(A)、微生物生物量碳(B)含量和微生物熵(C)
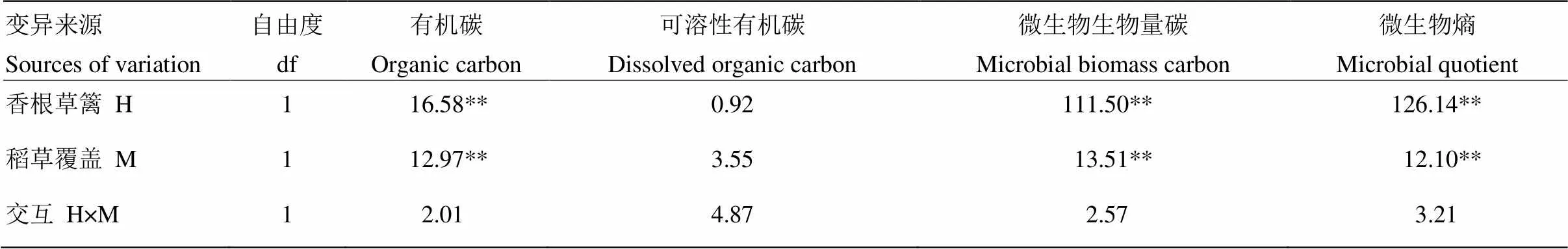
表1 香根草篱和稻草覆盖对土壤有机碳、可溶性有机碳、微生物量碳和微生物熵影响的方差分析
表中数据和星号为F值和显著水平,*0.05,**<0.01。下同
Data and asterisk in table represent F values and probability level, *<0.05, **<0.01. The same as below
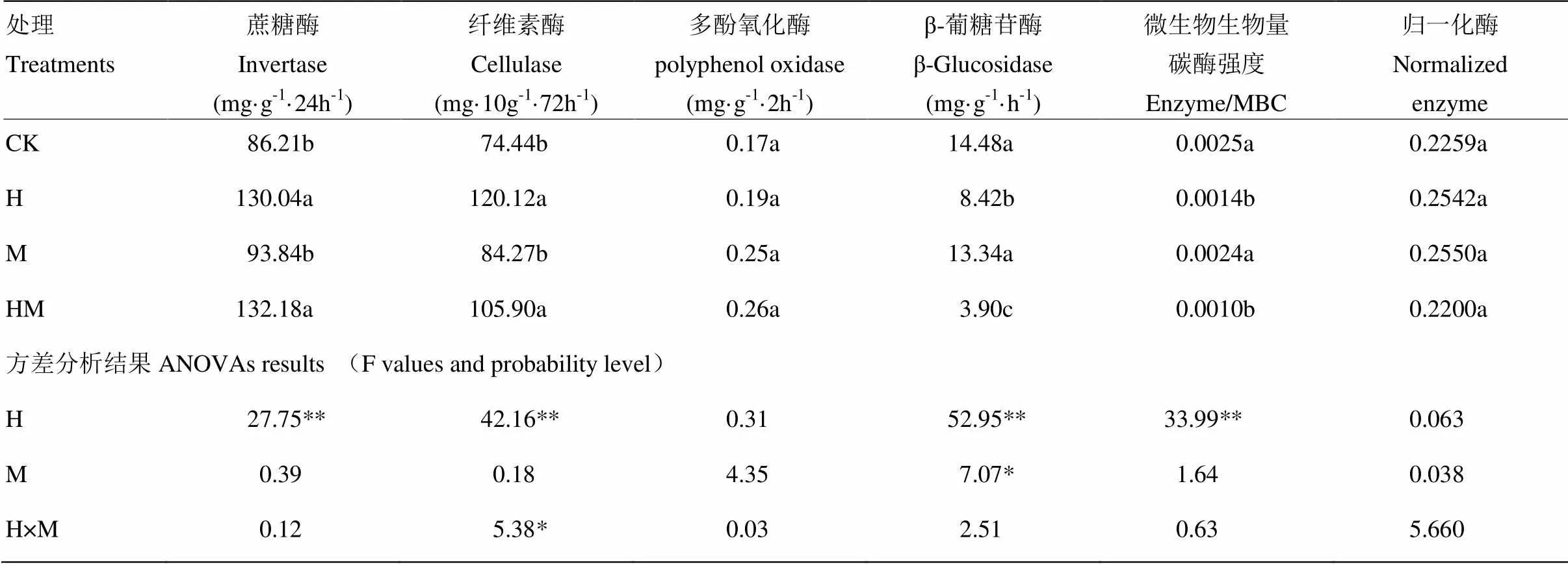
表2 香根草篱和稻草覆盖对土壤酶活性的影响及方差分析
表中处理间不同小写字母代表5%差异显著性。下同
Different letters among the treatments mean significantly difference at 0.05 level. The same as below
2.3 土壤微生物群落结构变化
2.3.1 土壤微生物磷脂脂肪酸(PLFA)含量变化及组成 PLFA 是活体微生物细胞膜的组分,特定的菌落PLFA数量变化可反映出原位土壤真菌(F)、细菌(B)活体生物量与菌落结构[23-24]。由表3可以看出,与CK相比,H、M和HM处理均增加了土壤微生物总PLFA(TB)的数量,不同水土保护措施下土壤各PLFAs含量具有明显差异,并且影响趋势和程度因微生物种类而不同(表3)。H处理的PLFAs 总量及细菌(B)、真菌(F)、革兰氏阳性菌(GP)、革兰氏阴性菌(GN)、放线菌(ACT)及AM真菌(AMF)含量均较高,且除了F和GN数量外,其他均显著高于CK处理;M处理主要增加了总PLFAs、B和ACT的PLFA量,较CK处理分别显著提高11.8%、13.4%和18.6%。
真菌/细菌(F/B)及革兰氏阴性菌/革兰氏阳性菌(GN/GP)与微生物群落结构紧密相关,其中F/B比值可以表征农田土壤生态系统的稳定性[25]。由表3可以看出,不同水土保护措施间F/B无明显差异,说明稻草覆盖和香根草篱短期内对红壤坡耕地土壤生态系统稳定性无显著影响。稻草覆盖和香根草篱主要增加了土壤中GP菌的数量,进而减少了土壤中GN/GP比例,尤以HM处理减少幅度最大,较CK显著降低13.4%。
2.3.2 主成分分析 由PLFAs主成分分析可知(图3),第一主成分(PC1 )贡献率为86.75%,第二主成分(PC2)贡献率为6.49%,两者累积解释了微生物群落结构变化的93.24%。CK处理投射点分布在第四象限,H和HM处理投射点分布在第二象限,M处理投射点分布在第三象限,说明红壤坡耕地在稻草覆盖、香根草篱等水土保护措施下土壤磷脂脂肪酸组成结构与CK处理间产生较大的差异。不同水土保持措施处理中,H和M处理土壤微生物群落结构相近,与HM处理的磷脂脂肪酸的结构差别较大。由土壤PLFAs在主成分上的因子载荷分析表明(图3),对PC1贡献较大的为土壤总PLFAs生物量(TB)、B、AMF、ACT、GP和GN;对PC2贡献较大的为GN/GP和F/B。

表3 不同水土保护措施下土壤微生物磷脂脂肪酸组成及含量(nmol·g-1)
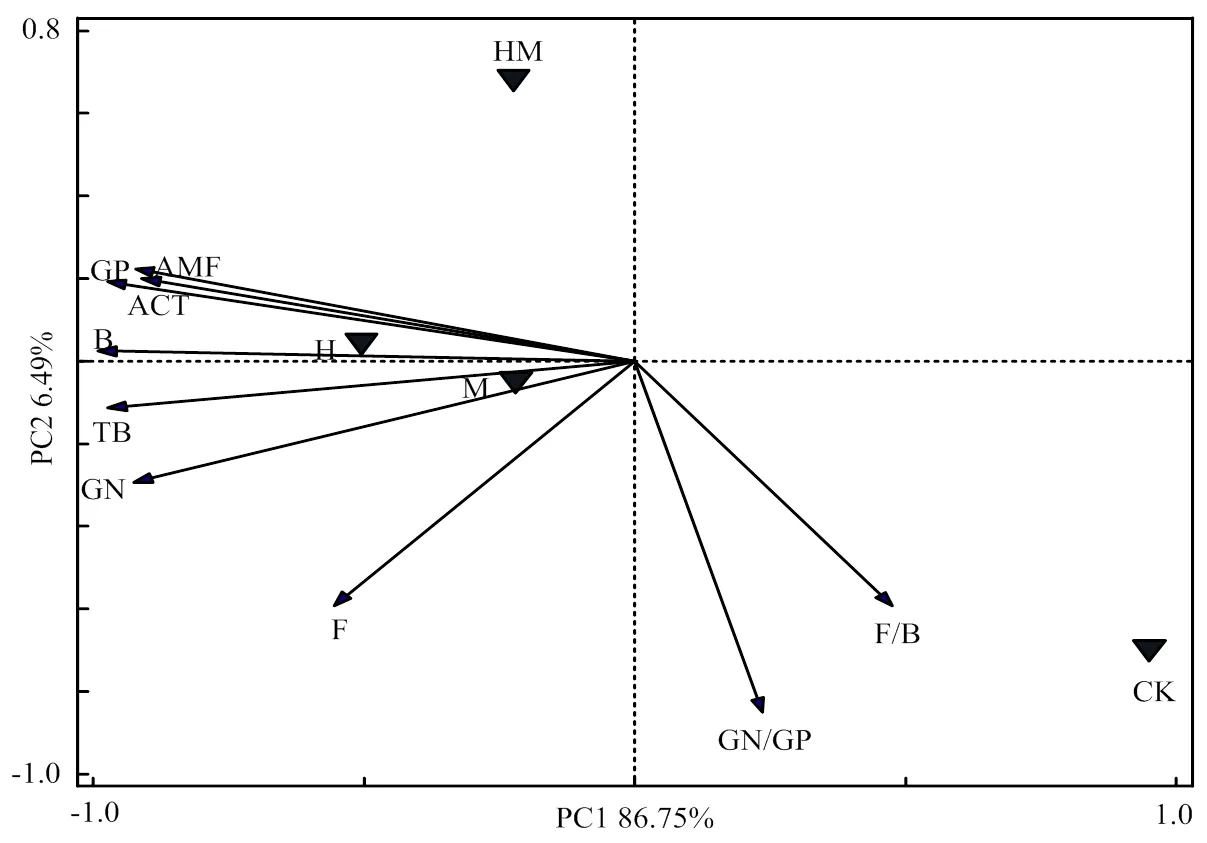
图3 不同水土保护措施下土壤PLFA主成分分析
2.4 各菌群PLFAs与土壤碳及参与碳循环的土壤酶活性的关系
冗余分析(RDA)表明,轴1和轴2解释了土壤菌落变异程度的95.37%。除了GN/GP和F/B外,土壤磷脂脂肪酸组成各指标之间及其和土壤MBC、TOC和DOC均呈正相关。在土壤酶活性指标中,土壤磷脂脂肪酸组成各指标与CEL和INV活性呈正相关,而与β-Glu活性及和碳循环酶有关的比例指标OCE和MBCE呈负相关。不同水土保护措施的影响差异明显,4个处理分别各占据1个象限,且H处理与除了β-Glu、OCE、MBCE、F/B和GN/GP外的所有指标呈正相关性(图4)。
3 讨论
3.1 稻草覆盖和香根草篱对土壤碳及参与碳循环酶活性的影响
土壤有机碳组分及其含量是衡量土壤肥力的重要指标[26],农田系统中作物根系及其分泌物、根系残茬、有机肥投入和秸秆还田等均是增加土壤有机碳输入量的直接因素[12,27-28]。本研究中,与CK相比,H、M和HM外源碳投入均分别显著增加土壤总有机碳1.09、1.00和1.52 g·kg-1,另一方面,新鲜有机物的输入增加了土壤微生物活性,促进有机物的固定,进而增加有机碳的积累[29]。微生物生物量碳和可溶性有机碳是土壤有机碳中最活跃的组分之一,易被微生物分解利用、对植物养分供应有直接作用,是土壤潜在生产力的重要评价指标[30]。香根草篱和稻草覆盖通过提高土壤养分含量、改善土壤环境状况[31],为土壤微生物繁殖和生长提供了养料和水热条件,加速了土壤有机碳分解和转化,进而增加了土壤可溶性碳和微生物量碳的含量。相比于草篱根系的拦截作用,稻草覆盖在减弱雨滴对地表溅蚀的同时延缓了雨水在坡面地表的集聚速度和强度,进而减少了可溶性有机碳随雨水的淋失[32],以及稻草分解过程中产生大量水溶性有机碳均是其对增加土壤可溶性碳含量效果优于草篱的原因。相比稻草覆盖处理,草篱根际微生物群落结构的改变及其复杂的根系分泌物可能是其影响土壤可溶性有机碳含量改变的原因[33];同时,草篱处理因根系作用致使可溶性有机碳含量随雨水淋溶[34]以及本试验中为保证香根草篱长势,每年不定期刈割并移走,造成凋落物等可溶性有机碳的主要来源物减少是造成香根草篱有降低土壤可溶性有机碳含量趋势的又一原因。土壤酶活性是土壤生物活性和土壤肥力的重要指标,对土壤养分溶解、生物化学过程反应速率等都具有显著促进作用[35]。本试验中除了β-葡糖苷酶,其他3种参与碳循环的酶活性在不同水土保护措施下都有一定增加,主要是因为草篱和稻草覆盖直接或间接增加了土壤碳源,促进了微生物的繁殖,刺激了酶活性的增加,这与黄尚书等[36]对同一试验地的旱季土壤研究结果相似,但不同的是,本研究中香根草篱对4种参与碳循环的酶活性的影响显著,是因为本试验采集的是距离草篱2 m范围内的土壤样品。距离植物篱越近,植物篱对土壤理化性质及作物生长的影响越明显[12],同时,草篱发达的根系代谢也会释放大量的酶类[37]。H和HM处理降低了β-葡糖苷酶活性,这可能与各类酶的功能及来源有关,具体原因还有待进一步研究,但这也说明用单一酶活性评价土壤质量的局限性,土壤微生物数量的多少与土壤酶活性强弱不一定一致[38]。稻草覆盖对单一酶活性的影响不明显,但显著增加了单位微生物量酶活性(MBCE),说明了稻草覆盖条件下微生物对酶活性的贡献。
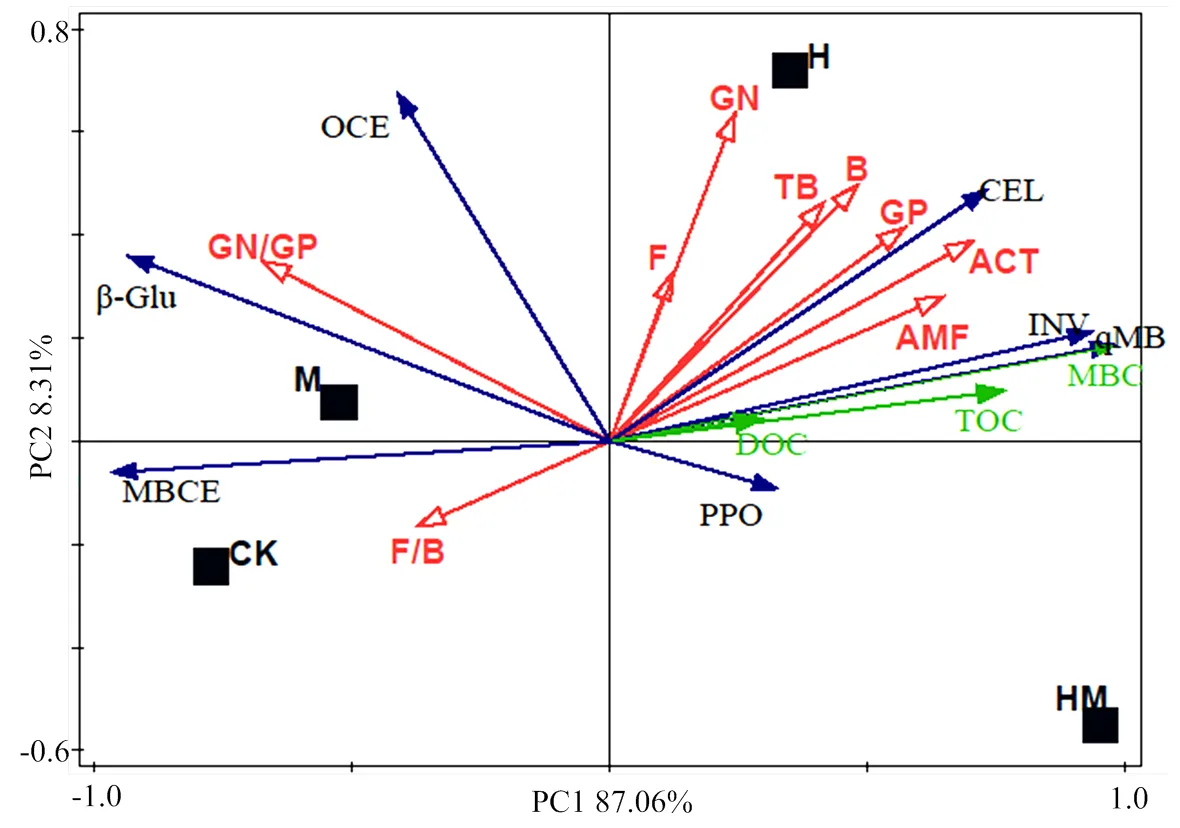
β-Glu:β-葡糖苷酶;INV:蔗糖酶;CEL:纤维素酶;PPO:多酚氧化酶;TOC:总有机碳;DOC:可溶性碳;MBC:微生物量碳;MBCE:单位微生物量碳酶活性;OCE:单位有机碳酶活性;GN:革兰氏阴性菌;GP:革兰氏阳性菌;TB:总生物量;B:细菌;F:真菌;ACT:放线菌;AMF:AM真菌
3.2 稻草覆盖和香根草篱对土壤微生物各群落PLFA 含量的影响
本研究中,稻草覆盖、香根草篱和二者结合都增加了土壤中PLFAs总量,细菌、真菌和放线菌数量,此研究结果与前人研究结果一致[18]。土壤PLFAs主成分分析表明,H和M处理土壤微生物群落结构相似,而与HM处理的磷脂脂肪酸的结构差别较大,一方面说明香根草篱和稻草覆盖通过凋落物和根系带入了大量可被微生物分解利用的碳源和氮源,为微生物的繁殖提供了物质基础[16],而输入土壤的有机物抗分解能力较强,是造成土壤中真菌和放线菌丰富度较高的原因[39];另一方面也说明香根草篱和稻草覆盖相结合相比单一措施土壤理化性质的变异程度不同,影响了土壤微生物的数量及组成结构。AMF是土壤与植物根系关系最密切的微生物之一[40],广泛分布在根际土壤中,因此本研究中有草篱处理(H和HM)增加了AMF的数量。值得提出的是,主成分分析结果(图3)与各处理间单项指标单因素分析结果(表3)不一致的原因是因为主成分分析是将多项指标转化为少数几项综合指标,用综合指标来解释多变量的方差-协方差结构,是对原有变量的综合。不同种类微生物磷脂脂肪酸(PLFA)组成和含量间的差异,可用来直接评估其微生物的生物量及群落结构。真菌/细菌的比例是土壤生态系统缓冲能力的重要指标,比例越高则生态缓冲能力越高[25]。有研究表明秸秆还田显著提高了真菌/细菌比例[41],但本研究中真菌/细菌的比例并未有显著变化,可能是由于稻草覆盖和草篱等对细菌和真菌群落间丰度增加量相互抵消所致[41]。革兰氏阳性菌(GP)和革兰氏阴性菌(GN)是土壤细菌中重要组成部分,尤其对凋落物的分解起重要作用[15]。GN/GP值可以指示土壤营养状况,GN/GP越小,表示营养胁迫越强[42-43]。H、M及HM虽然都增加了GP和GN的数量,但GN/GP与CK相比呈下降趋势,且以HM处理GN/GP最小。H和M处理向土壤提供了丰富的有机碳源,但高碳氮比有机物的分解加剧了微生物和植物之间的养分竞争,因此H和M处理土壤微生物群落结构更倾向于向擅长竞争资源的革兰氏阳性细菌方向转变[44]。
土壤理化性质的改变是影响微生物群落结构的主要原因,有机碳的种类和数量决定了微生物的种类和数量,并对微生物的生长繁殖及代谢过程产生影响。土壤酶作为土壤环境的重要组分,其与土壤微生物以及有机碳等理化性质之间存在密切的关系。而土壤微生物群落的动态变化反过来也影响着土壤系统的物质循环和能量流动[45]。冗余分析结果显示,土壤MBC、TOC,CEL、 INV、β-Glu活性及碳循环酶有关的比例指标OCE和MBCE均是影响土壤微生物数量和结构的主要因子。H处理中土壤MBC、TOC和CEL、 INV活性主要影响了B、ACT、AMF、GN、GP等微生物数量和总PLFAs量;M处理中β-Glu活性及OCE和MBCE主要影响了微生物结构指标GN/GP 和F/B,说明本实验中香根草篱和稻草覆盖由于不同密度、不同种类植物的凋落物和根系分泌物中输入碳的种类和数量不同[46-47],以及两种措施不同水土保护机制造成土壤养分及结构各异[12],进而影响土壤微生物的数量及组成结构,而香根草篱和稻草覆盖相结合加剧了与单一措施下土壤理化性质的变异程度,这也可能是导致HM处理土壤微生物结构不同于H和M处理的原因。同时,尽管PLFA技术常被用于研究土壤微生物群落结构,但该技术本身也有局限性。PLFA能定量描述环境样品中的微生物群体,而不能在种的水平精确的描述微生物的种类,且从土壤中提取出的各类脂肪酸种类仍十分有限[47],因此,要全面解析土壤微生物群落结构还需结合其他的研究手段。
4 结论
稻草覆盖和香根草篱通过秸秆、根系残茬和根系分泌物等直接或间接外源碳的投入均能显著增加土壤总有机碳含量及可溶性碳和微生物量碳等活性有机碳的含量。磷脂脂肪酸分析结果表明,香根草篱和稻草覆盖因不同密度、不同种类植物的凋落物和根系分泌物输入碳的种类和数量不同,以及两种措施不同水土保护机制造成土壤养分及结构各异,磷脂脂肪酸的种类及数量也会产生差异。同时,香根草篱和稻草覆盖降低了革兰氏阴性菌磷脂脂肪酸与革兰氏阳性菌磷脂脂肪酸的比值,且尤以二者结合处理显著。由此说明,香根草篱和稻草覆盖等生物水土保护措施在短期内是控制坡耕地水土流失,维持和提升耕地质量的有效措施,但长期应用要关注土壤碳氮养分均衡状态,以实现南方红壤坡耕地的可持续发展。
[1] 赵其国. 我国红壤的退化问题. 土壤, 1995, 27(6): 281-285.
ZHAO Q G. The red soil degradation of China., 1995, 27(6): 281-285. (in Chinese)
[2] 于东升, 史学正. 低丘红壤区旱地土壤渗透性与可蚀性定量关系的研究. 土壤学报, 2000, 37(3): 316-322.
YU D S, SHI X Z. Quantification relationship between soil permeability of upland and soil erodibility in hilly red soil region., 2000, 37(3): 316-322. (in Chinese)
[3] 梁音, 张斌, 潘贤章, 史德明. 南方红壤丘陵区水土流失现状与综合治理对策. 中国水土保持科学, 2008, 6(1): 22-27.
LIANG Y, ZHANG B, PAN X Z, SHI D M. Current status and comprehensive control strategies of soil erosion for hilly region in the Southern China., 2008, 6(1): 22-27. (in Chinese)
[4] 蒲玉琳, 谢德体, 林超文, 丁恩俊, 张坤, 魏朝富. 植物篱-农作坡耕地土壤微团聚体组成及分形特征. 土壤学报, 2012, 49(6): 1069-1077.
PU Y L, XIE D T, LIN C W, DING E J, ZHANG K, WEI Z F. Composition and fractal features of soil icro-aggregates in sloping farmland with hedgerow., 2012, 49(6): 1069-1077. (in Chinese)
[5] 裴峥, 段喜明. 翻耕和覆盖对坡耕地产流产沙的影响. 山西农业大学学报(自然科学版), 2016, 36(6): 412-416.
PEI Z, DUAN X M. Effect of tilling and mulching on runoff yield and sediment yield in slope farmland.(), 2016, 36(6): 412-416. (in Chinese)
[6] 钟义军, 叶川, 黄欠如, 章新亮, 武琳, 孙永明, 秦江涛. 红壤缓坡花生地不同水土保持措施效果分析. 中国水土保持科学, 2011, 9(3): 71-74.
ZHONG Y J, YE C, HUANG Q R, ZHANG X L, WU L, SUN Y M, QIN J T. Benefit analysis of different soil and water conservation measures at slopingland with red soil., 2011, 9(3): 71-74. (in Chinese)
[7] 李新平, 王兆骞, 陈欣, 方志国. 红壤坡耕地人工模拟降雨条件下植物篱笆水土保持效应及机理研究. 水土保持学报, 2002, 16(2): 36-40.
LI X P, WANG Z Q, CHEN X, FANG Z G. Research on soil and water conservation effect and mechanism of hedges under rainfall simulation in red soil slope field., 2002, 16(2): 36-40. (in Chinese)
[8] 袁久芹, 梁音, 曹龙熹, 卢慧中, 祝亚云, 陈玲. 红壤坡耕地不同植物篱配置模式减流减沙效益对比. 土壤, 2015, 47(2): 400-407.
YUAN J Q, LIANG Y, CAO L X, LU H Z, ZHU Y Y, CHEN L. Comparison of benefits of runoff and sediment reduction of different hedgerow configuration modes in red-soil hilly area, 2015, 47(2): 400-407. (in Chinese)
[9] 郑海金, 杨洁, 黄鹏飞, 万佳蕾, 王凌云, 赵佳鼎. 覆盖和草篱对红壤坡耕地花生生长和土壤特性的影响. 农业机械学报, 2016, 47(4): 119-126.
ZHENG H J, YANG J, HUANG P F, WAN J L, WANG L Y, ZHAO J D. Effects of straw mulching and vetiver grass hedgerows on peanut growth and soil property in red soil sloping field., 2016, 47(4): 119-126. (in Chinese)
[10] 蒲玉琳, 林超文, 谢德体, 魏朝富, 倪九派. 植物篱-农作坡地土壤团聚体组成和稳定性特征.应用生态学报, 2013, 24(1): 122-128.
PU Y L, LIN C W, XIE D T, WEI Z F, NI J P. Composition and stability of soil aggregates in hedgerow-crop slope land., 2013, 24(1): 122-128. (in Chinese)
[11] HUANG D, HAN J G, WU J Y, WANG K, WU W L, TENG W J, SARDO V. Grass hedges for the protection of sloping lands from runoff and soil loss: An example from Northern China., 2010, 110: 251-256.
[12] 成艳红, 武琳, 孙慧娟, 钟义军, 孙永明, 章新亮, 黄尚书, 黄欠如. 稻草覆盖和香根草篱对红壤水稳性团聚体组成及有机碳含量的影响.生态学报, 2016, 36(12): 3518-3524..
CHENG Y H, WU L, SUN H J, ZHONG Y J, SUN Y M, ZHANG X L, HUANG S S, HUANG Q R. Effects of straw mulching and Vetiver grass hedgerows on the size distribution of the soil water stable aggregates and aggregate-associated organic carbon in red soil., 2016, 36(12):3518-3524. (in Chinese)
[13] 彭新华, 张斌, 赵其国. 土壤有机碳库与土壤结构稳定性关系的研究进展. 土壤学报, 2004, 41(4): 618-623.
PENG X H, ZHANG B, ZHAO Q G. A review on relationship between soil otganic carbon pools and soil structure stability., 2004, 41(4): 618-623. (in Chinese)
[14] 彭新华, 张斌, 赵其国. 红壤侵蚀裸地植被恢复及土壤有机碳对团聚体稳定性的影响. 生态学报, 2003, 23(10): 2176-2183.
PENG X H, ZHANG B, ZHAO Q G. Effects of soil organic carbon on aggregate stability after vegetative restoration on severely eroded red soil., 2003, 23(10): 2176-2183. (in Chinese)
[15] 毕明丽, 宇万太, 姜子绍, 马强, 张璐, 徐永刚. 利用PLFA 方法研究不同土地利用方式对潮棕壤微生物群落结构的影响. 中国农业科学, 2010, 43(9): 1834-1842.
BI M L, YU W T, JIANG Z S, MA Q, ZHANG L, XU Y G. Study on the effects of different land use patterns on microbial community structure in aquic brown soil by utilizing PLFA method., 2010, 43(9): 1834-1842. (in Chinese)
[16] VALMIS S, DIMOYIANNIS D, DANALATOS N G. Assessing interrill erosion rate from soil aggregate instability index, rainfall intensity and slope angle on cultivated soils in central Greece., 2005, 80(1/2): 139-147.
[17] 郭甜, 何丙辉, 蒋先军, 马云, 吴咏, 向明辉, 谌芸, 唐春霞. 紫色土区植物篱对坡面土壤微生物特性的影响. 水土保持学报, 2011, 25(5): 97-98.
GUO T, HE B H, JIANG X J, MA Y, WU Y, XIANG M H, CHEN Y, TANG C X. Effects of hedgerows on soil microbial characteristics of slope in purple soil area., 2011, 25(5): 97-98. (in Chinese)
[18] 王玉川, 陈强, 林超文, 李浩. 不同植物篱对石灰性紫色土坡耕地土壤微生物学特性影响研究. 土壤, 2012, 44(1): 73-77.
WANG Y C, CHEN Q, LIN C W, LI H. Microbial features of sloping farmland soil in calcareous purple area affected by different hedgerows treatments in Sichuan Basin., 2012, 44(1): 73-77. (in Chinese)
[19] 王芸, 韩宾, 史忠强, 邵国庆, 江晓东, 宁堂原, 焦念元, 李增嘉. 保护性耕作对土壤微生物特性及酶活性的影响. 水土保持学报, 2006, 20(4): 120-122, 142.
WANG Y, HAN B, SHI Z Q, SHAO G Q, JIANG X D, NING T Y, JIAO N Y, LI Z J. Effects of conservation tillage on soil microbial characters and soil enzyme activities., 2006, 20(4): 120-122, 142. (in Chinese)
[20] 关松荫. 土壤酶及其研究方法. 北京: 农业出版社, 1986.
GUAN S Y.Beijing: Agriculture Press, 1986. (in Chinese)
[21] 吴金水, 林启美, 黄巧云, 肖和艾. 土壤微生物生物量测定方法及其应用. 北京: 气象出版社, 2006.
WU J S, LIN Q M, HUANG Q Y, XIAO H A.. Beijing: Meteorological Press, 2006. (in Chinese)
[22] BLIGH E G, DYER W J. A rapid method of total lipid extraction and purification., 1959, 37: 911-917.
[23] TUNLID A, BAIRD B H, TREXLER M B. Determination of phospholipid ester-linked fatty acids and poly β-hydroxybutyrate for the estimation of bacterial biomass and activity in the rhizosphere of the rape plant(L.)., 1985, 31(12):1113-1119.
[24] ZELLES L, BAI Q Y, BECK T, BEESE F. Signature fatty acids in phospholipids and lipopolysaccharides as indicators of microbial biomass and community structure in agricultural soils., 1992, 24: 317-323.
[25] DE VRIES F T, HOFFLAND E, VAN EEKEREN N, BRUSSAARD L, BLOEM J. Fungal/bacterial ratios in grasslands with contrasting nitrogen management., 2006, 38: 2092-2103.
[26] 黄丽, 蔡崇法, 丁树文, 张光远. 几种绿篱梯田中紫色土有机质组分及其性质的研究. 华中农业大学学报, 2000, 19(6): 559-562.
HUANG L, CAI C F, DING S W, ZHANG G Y. Study on the components of organic matter and its properties of purple soil in several hedgerow terraces., 2000, 19(6): 559-562. (in Chinese)
[27] 张敬业, 张文菊, 徐明岗, 黄庆海, 骆坤. 长期施肥下红壤有机碳及其颗粒组分对不同施肥模式的响应. 植物营养与肥料学报, 2012, 18(4): 868-875.
ZHANG J Y, ZHANG W J, XU M G, HUANG Q H, LUO K. Response of soil organic carbon and its particle-size fractions to different long-term fertilizations in red soil of China., 2012, 18(4): 868-875. (in Chinese)
[28] 刘春增, 常单娜, 李本银, 曹卫东, 吕玉虎, 潘兹亮. 种植翻压紫云英配施化肥对稻田土壤活性有机碳氮的影响. 土壤学报, 2017, 54(3) :656-668.
LIU C Z, CHANG D N, LI B Y, CAO W D, LV Y H, PAN Z L. Effects of planting and incorporation of Chinese milk vetch coupled with application of chemical fertilizer on active organic carbon and nitrogen in paddy soil., 2017, 54(3): 656-668. (in Chinese)
[29] GREGORICH E G, CARTER M R, ANGERS D A, MONREAL C M, ELLERT B H. Towards a minimum data set to assess soil organic matter quality in agricultural soils., 1994, 74: 376-385.
[30] 王保君, 王伟, 胡乃娟, 顾泽海, 陈兵林, 张政文, 徐蒋来, 朱利群. 麦秸还田下水氮管理对稻田土壤养分、酶活性及碳库的短期影响. 核农学报, 2016, 30(5): 957-964.
WANG B J, WANG W, HU N J, GU Z H, CHEN B L, ZHANG Z W, XU J L, ZHU L Q. Short-term effect of different water and nitrogen managements on paddy soil nutrient,enzyme activity and carbon pool under wheat straw-returning fields., 2016, 30(5): 957-964. (in Chinese)
[31] 柳开楼, 李亚贞, 秦江涛, 胡惠文, 周利军, 黄欠如, 章新亮, 刘满强, 范红杰. 香根草篱和稻草覆盖对红壤坡耕地土壤肥力的影响. 土壤, 2015, 47(2): 305-309.
LIU K L, L I Y Z, QIN J T, HU H W, ZHOU L J, HUANG Q R, ZHANG X L, LIU M Q, FAN H J. Effects of straw mulching and vetiver hedge on soil fertility in sloping land of red soil., 2015, 47(2): 305-309. (in Chinese)
[32] SCHIMEL J P, WEINTRAUB M N. The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: a theoretical model., 2003, 35(4): 549-563.
[33] 张金波, 宋长春, 杨文燕. 小叶章湿地表土水溶性有机碳季节动态变化及影响因素分析. 环境科学学报, 2005, 25(10): 1397-1402.
ZHANG J B, SONG C C, YANG W Y. Seasonal dynamics of dissolved organic carbon and its impact factors in themarsh soil., 2005, 25(10): 1397-1402. (in Chinese)
[34] KALBITZ K, SOLINGER S, PARK J H, MICHALZIK B, MATZNER E. Controls on the dynamics of dissolved organic matter in soils: a review.2000, 165: 277-304.
[35] TRASAR-CEPEDA C, LEIRÓS M C, GIL-SOTRES F. Hydrolytic enzyme activities in agricultural and forest soils. Some implications for their use as indicators of soil quality., 2008, 40(9): 2146-2155.
[36] 黄尚书, 成艳红, 钟义军, 孙永明, 黄欠如, 武琳, 章新亮, 许彦. 水土保持措施对红壤缓坡地土壤活性有机碳及酶活性的影响. 土壤学报, 2016, 53(2): 468-476.
HUANG S S, CHENG Y H, ZHONG Y J, SUN Y M, HUANG Q R, WU L, ZHANG X L, XU Y. Effects of different soil and water conservation measures on soil labile organic and soil enzyme activity in slope land of red soil.2016, 53(2): 468-476. (in Chinese)
[37] 杨宁, 邹冬生, 杨满元, 陈璟, 陈志阳, 林仲桂, 宋光桃. 衡阳紫色土丘陵坡地不同植被恢复阶段土壤酶活性特征研究. 植物营养与肥料学报, 2013, 19(6): 1516-1524.
YANG N, ZOU D S, YANG M Y, CHEN J, CHEN Z Y, LIN Z G, SONG G T. Soil enzyme activities in different re-vegetation stages on sloping-land with purple soils in Hengyang of Hunan Province, China., 2013, 19(6): 1516-1524. (in Chinese)
[38] 马东云, 郭天财, 宋晓, 王晨阳, 朱云集, 王永华, 岳艳军, 査菲娜. 尿素施用量对小麦根际土壤微生物数量及土壤酶活性的影响. 生态学报, 2007, 7(12): 5222-5228.
MA D Y, GUO T C, SONG X, WANG C Y, ZHU Y J, WANG Y H, YUE Y J, CHA F N. Effects of urea application rate on the quantity of microorganisms and activity of enzymes in wheat rhizosphere., 2007, 7(12): 5222-5228. (in Chinese)
[39] 樊芳玲, 谢德体, 魏朝富, 倪九派, 唐振亚. 坡耕地⁃桑树系统土壤微生物群落结构的PLFA 分析. 生态学报, 2016, 36(12): 3812-3823.
FAN F L, XIE D T, WEI Z F, NI J P, TANG Z Y. PLFA analysis showed that the configuration modes of sloping crop-mulberry systems affect the soil microbial community structure., 2016, 36(12): 3812-3823. (in Chinese)
[40] 叶佳舒, 李涛, 胡亚军, 郝志鹏, 高彦征, 王幼珊, 陈保东. 干旱条件下AM 真菌对植物生长和土壤水稳定性团聚体的影响. 生态学报, 2013, 33(4): 1080-1090.
YE J S, LI T, HU Y J, HAO Z P, GAO Y Z, WANG Y S, CHEN B D. Influence of AM fungi on plant growth and water-stable soil aggregates under drought stresses.2013, 33(4): 1080-1090. (in Chinese)
[41] 强学彩, 袁红莉, 高旺盛. 秸秆还田量对土壤CO2释放和土壤微生物量的影响. 应用生态学报, 2004, 15(3): 469-472.
QIANG X C, LI T, HU Y J, HAO Z P, GAO Y Z, WANG Y S, CHEN B D. Effect of crop-residue incorporation on soil CO2emission and soil microbial biomass., 2004, 15(3): 469-472. (in Chinese)
[42] 郭梨锦, 曹凑贵, 张枝盛, 刘天奇, 李成芳. 耕作方式和秸秆还田对稻田表层土壤微生物群落的短期影响. 农业环境科学学报, 2013, 32(8): 577-1584.
GUO L J, CAO Z G, ZHANG Z S, LIU T Q, LI C F. Short-term effects of tillage practices and wheat-straw returned to rice fields on topsoil microbial community structure and microbial diversity in central China., 2013, 32(8): 1577-1584. (in Chinese)
[43] THIET R K, FREY S D, SIX J. Do growth yield efficiencies differ between soil microbial communities differing in fungal: bacterial ratios? Reality check and methodological issues., 2006, 38(4): 837-844.
[44] DE VRIES F T, HOFFLAND E, VAN EEKEREN N, BRUSSAARD L, BLOEM J. Fungal/bacterial ratios in grasslands with contrasting nitrogen management., 2006, 38(8): 2092-2103.
[45] 刘满强, 陈小云, 郭菊花, 李辉信, 胡锋. 土壤生物对土壤有机碳稳定性的影响. 地球科学进展, 2007, 22(2): 152-158.
LIU M Q, CHEN X Y, GUO J H, LI H X, HU F. Soil biota on soil organic carbon stabilization., 2007, 2(2): 152-158.
[46] 雷海迪, 尹云锋, 刘岩, 万晓华, 马红亮, 高人, 杨玉盛. 杉木凋落物及其生物炭对土壤微生物群落结构的影响. 土壤学报, 2016(3): 790-799.
LEI H D, YIN Y F, LIU Y, WAN X H, MA H L, GAO R, YANG Y S. Effects of fir () litter and its biochar on soil microbial community structure., 2016(3): 790-799. (in Chinese)
[47] 卜洪震, 王丽宏, 尤金成, 肖小平, 杨光立, 胡跃高, 曾昭海. 长期施肥管理对红壤稻田土壤微生物量碳和微生物多样性的影响. 中国农业科学, 2010, 43(16): 3340-3347.
BU H Z, WANG L H, YOU J C, XIAO X P, YANG G L, HU Y G, ZENG Z H. Impact of long-term fertilization on the microbial biomass carbon and soil microbial communities in Paddy Red Soil., 2010, 43(16): 3340-3347. (in Chinese)
(责任编辑 李云霞)
Effects of Straw Mulching andHedgerows on Soil Enzyme Activities and Soil Microbial Community Structure in Red Soil Sloping Land
CHENG YanHong, HUANG QianRu, WU Lin, HUANG ShangShu, ZHONG YiJun, SUN YongMing, ZHANG Kun, ZHANG XinLiang
(Jiangxi Institute of Red Soil/Jiangxi Key Laboratory of Red Soil Arable Land Conservation/National Engineering and Technology Research Center for Red soil Improvement/Jiangxi Province Scientific Observation and Experimental Station of Arable Land Conservation, Ministry of Agriculture, Jinxian 331717, Jiangxi)
The objective of this paper is to clarify the relationship between soil organic carbon and microorganism under different biological water protection measures.Soil samples were collected from a field soil and water conservation measures experiment, which was started since 2009 with 4 treatments, including peanuts grown in high order (CK), hedgerows+ peanuts (H), mulching+ peanuts (M) and hedgerows+ mulching+ peanuts (HM). Phospholipid fatty acid (PLFA) profiles for each treatment were used to characterize the microbial community structure. Soil organic carbon, microbial biomass carbon, dissolved organic carbon and soil enzyme activities were also determined.Compared with the CK, all treatments showed total organic carbon, dissolved organic carbon, total microbial biomass carbon and qMB increased. H is the main influence factor on MBC and Qmb. The results showed that H significantly affected the activities of soil invertase, cellulase, β-Glu and intensity of MBC/TOC. Compared with the CK, H, M and HM, PLFA increased the total PLFAs, bacterial and actinomycetes PLFAs, while decreased G-PLFA/G+PLFA, and that was the lowest under HM treatment. Principal components analysis showed that soil microbial community structure was similar to that of H and M, which was larger difference with CK. Redundancy analysis showed that TOC, MBC and enzymes activities of soil CEL, INV, β-Glu were all the main factors affecting the quantity and structure of soil microorganisms.Microbial biomass, community structure and soil normalized C-acquiring enzyme activity were influenced significantly by straw mulching and vetiver grass hedgerows.
red soil sloping land; straw mulching;Hedgerows; soil enzyme activity; microbial community structure
2017-05-16;
2017-08-06
国家自然科学基金(41301235)、江西省科技支撑项目(20151BBF60060)
联系方式:成艳红,E-mail:yanhongch007@163.com。通信作者黄欠如,E-mail:qianruhuang@163.com
猜你喜欢
环球时报(2023-01-16)2023-01-16
中国水土保持(2022年2期)2022-04-07
中国土壤与肥料(2021年5期)2021-12-02
昆钢科技(2021年6期)2021-03-09
中国水土保持(2018年5期)2018-05-10
海峡姐妹(2018年1期)2018-04-12
中国水利(2015年24期)2015-04-25
小说月刊(2014年10期)2014-11-18
中国水土保持(2014年3期)2014-04-02
短篇小说(2014年11期)2014-02-27
