
湿涝对花生干物质积累与分配的影响
2017-12-14 18:36:04刘登望王建国万书波
花生学报 2017年3期
易 静,刘登望,2,王建国,张 昊,2,郭 峰,万书波*,李 林,2*
(1. 湖南农业大学农学院,湖南 长沙 410128; 2. 湖南农业大学旱地作物研究所,湖南 长沙 410128;3.山东省作物遗传改良与生态生理重点实验室,山东 济南 250100)
湿涝对花生干物质积累与分配的影响
易 静1,刘登望1,2,王建国1,张 昊1,2,郭 峰3,万书波3*,李 林1,2*
(1. 湖南农业大学农学院,湖南 长沙 410128; 2. 湖南农业大学旱地作物研究所,湖南 长沙 410128;3.山东省作物遗传改良与生态生理重点实验室,山东 济南 250100)
为确定花生耐涝品种的干物质积累与分配特征及洪涝灾害的防控对策,选用18个耐涝性差异稳定的花生品种,在适合水旱轮作(土壤渗水性强)的大田条件下于营养生长末期进行短期湿涝10d(W10)、长期湿涝88d(W88)和正常灌溉(NI)处理,研究湿涝对不同花生品种各器官干物质积累和分配的影响。结果表明,短期湿涝对整个花生品种生物量的影响,以叶片最甚,其他依次是茎秆、荚果,而根系略增重;长期湿涝会降低叶片生物量,而促进根系、茎秆、荚果的增加,说明光合器官叶片对湿涝最敏感,其次是茎秆,荚果受短期湿涝影响小且因长期湿涝而增产,而根系具有稳定耐性。短、长涝期时实际产量较高的品种一般表现出矮秆、叶多、根系发达、根冠比较高,而耐性系数多偏低,短、长涝时叶片与根系增重可实现耐渍涝与高产的统一,根系、叶片生物量比重高是维持长涝耐性的基础。筛选出豫花15、花119、彩珠、桂红花、金花1012等耐渍涝品种。
花生;湿涝;干物质生产;产量;耐涝系数
花生是我国主要油料和经济作物[1]。花生的生长发育受很多因素影响,水分是重要因素之一[2]。渍涝是制约我国花生乃至整个农业生产发展的重要因素[3]。在我国南方大部分地区,春天梅雨季节导致花生萌芽出苗期渍涝;夏季高温多雨也会引起渍涝。研究表明[4-8],土壤水分过多会造成花生等豆科作物生长缓慢、干物质积累减少、降低植物根冠比、影响花生品质,但对不同花生品种的影响有差异。豆科作物承受湿涝的时间一般为2~4 d,随作物品种、生育时期、土壤类型和年份等有差异。花生幼苗期湿涝处理1~2d,对其根系干重影响较小,但对地上部干重影响较大;若湿涝处理延长至4~6d时影响甚大[9]。湿涝同样导致小麦[10-11]、玉米[12-14]减产,但湿涝对红薯有增产作用[15]。本研究采用7大类18个花生品种[16],比较湿涝与正常水分条件下光合产物在不同器官的积累与分配规律,以及物质积累分配特性与耐湿涝指标的关系,进一步为基于产量的耐湿涝性差异评价提供依据。
1 材料与方法
1.1 试验材料
2002年在长沙基地进行本研究的预备试验,参试材料为来自国内外的117份花生栽培种品种,从中筛选出耐湿涝性差异稳定的品种18份[16](表1),其中多份已经历耐湿涝改良。2003年对筛选出的品种进行耐涝性鉴定试验。试验土壤为第四纪红土发育成的水稻土,四周环境较一致,土壤渗水性强,适合水旱轮作,土壤肥力中等。
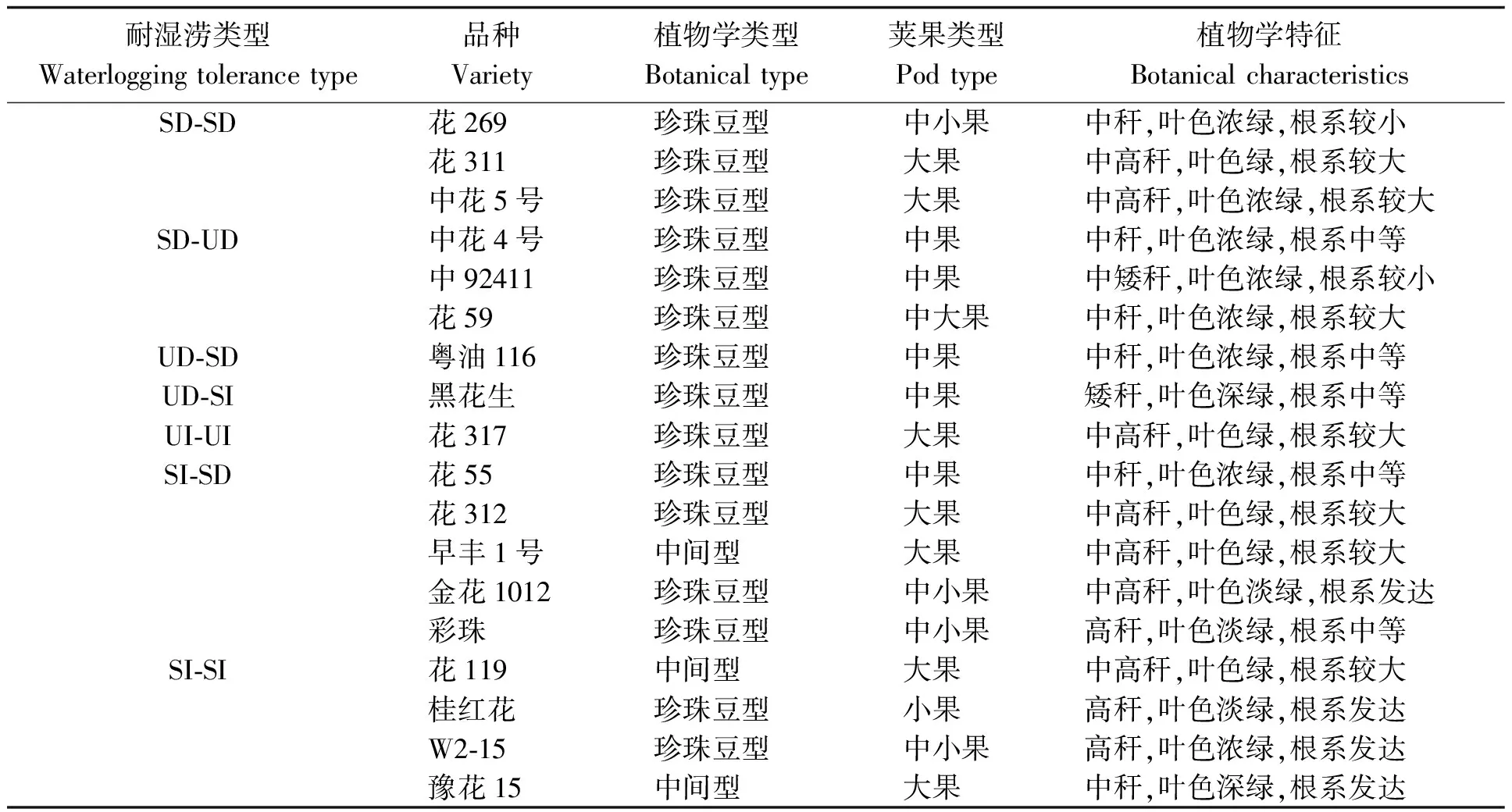
表1 供试花生品种及其耐湿涝分类
注:SI:比正常灌溉增产9.5%以上即耐湿涝系数大于1.095的品种,增产达显著或极显著水平;SD:减产8.0%即耐湿涝系数小于0.920的品种,减产达显著或极显著水平;UI:增产5.5%~8.7%即耐湿涝系数介于1.055~1.087的品种,增产不显著;UD:减产2.5%~6.0%即耐湿涝系数介于0.940~0.975的品种,减产不显著。前为长涝的反应,后为短涝的反应。下同。
Note: SI: the variety that the yield increased 9.5% more than normal irrigation and the coefficient of waterlogging tolerance was greater than 1.095, the increased yield of which was significant or very significant. SD: the variety that the yield decreased by 8% and the coefficient of waterlogging tolerance was less than 0.920, the decreased yield of which was significantly or extremely significantly. UI: the variety that the yield increased was 5.5% to 8.7% and the coefficient of waterlogging tolerance ranged from 1.055 to 1.087, the increased yield of which was not significant. UD: the variety that the yield decreased ranged from 2.5% to 6% and the coefficient of waterlogging tolerance ranged from 0.940 to 0.975, the decreased yield of which was not significant. The former is the reaction of long waterlogged response, the later is the reaction of short waterlogged response. The same as below.
1.2 处理与方法
试验采用裂区设计,主区为3个水分处理(短期湿涝、长期湿涝、正常灌溉),副区为18个花生品种,共计54个处理组合,重复3次,随机排列。正常灌溉(NI)时,根据花生的需水特性,土壤水分管理采取生育前期干爽、中期湿润、后期干爽的方式。2个湿涝处理均在花生苗期(大约播种后25d)进行人工输水。由于土壤渗水性强,湿涝期间一直保持流水灌溉,以维持土面持水1~2cm,其中短期湿涝持续10d,记作W10,简称短涝,此后正常灌溉;长期湿涝一直保持到成熟期的前第5d,共处理88d,记作W88,简称长涝。各主区间筑土埂,在土埂上下再覆盖3层农膜相隔,田沟深度70cm,宽度40cm,以防串水影响。主区面积60m2,副区面积3.33m2。每个品种种植5行,每行8穴,每穴播种4粒,定苗2株。按照一般大田生产水平进行肥水和病虫害管理。
1.3 测试项目
1.3.1 生物量与分配的测定
在花生成熟期,每个处理的各重复挖取10株完整植株,冲洗干净,擦干水分后,分成茎、叶、根、果4个部分,在105℃杀青20min,然后80℃烘干直至恒重。用天平称量各器官的干物质量。再计算单株干重、根冠比和经济系数。
1.3.2 数据统计方法
本试验采用单株籽仁烘干产量来分析处理间产量差异。其方法是:花生晒干后,各处理均称取100g荚果剥壳取籽仁,之后放入恒温80℃干燥箱中烘48h左右,直至恒重后,称重,计算烘干率,以此推算小区烘干籽仁总产量,该总产量除以实收株数即得单株籽仁产量,称实际产量(AY),湿涝时称湿涝产量。参照有关文献[17]计算指标:耐湿涝系数(WTC),即某品种湿涝条件下的产量与该品种在正常灌溉条件下产量之比,用以表征稳产性。采用SPSS统计软件中的方差、显著性差异分析。
2 结果与分析
2.1 各器官干物质产量对湿涝的反应以及与耐湿涝指标的关系
2.1.1 茎秆生物量
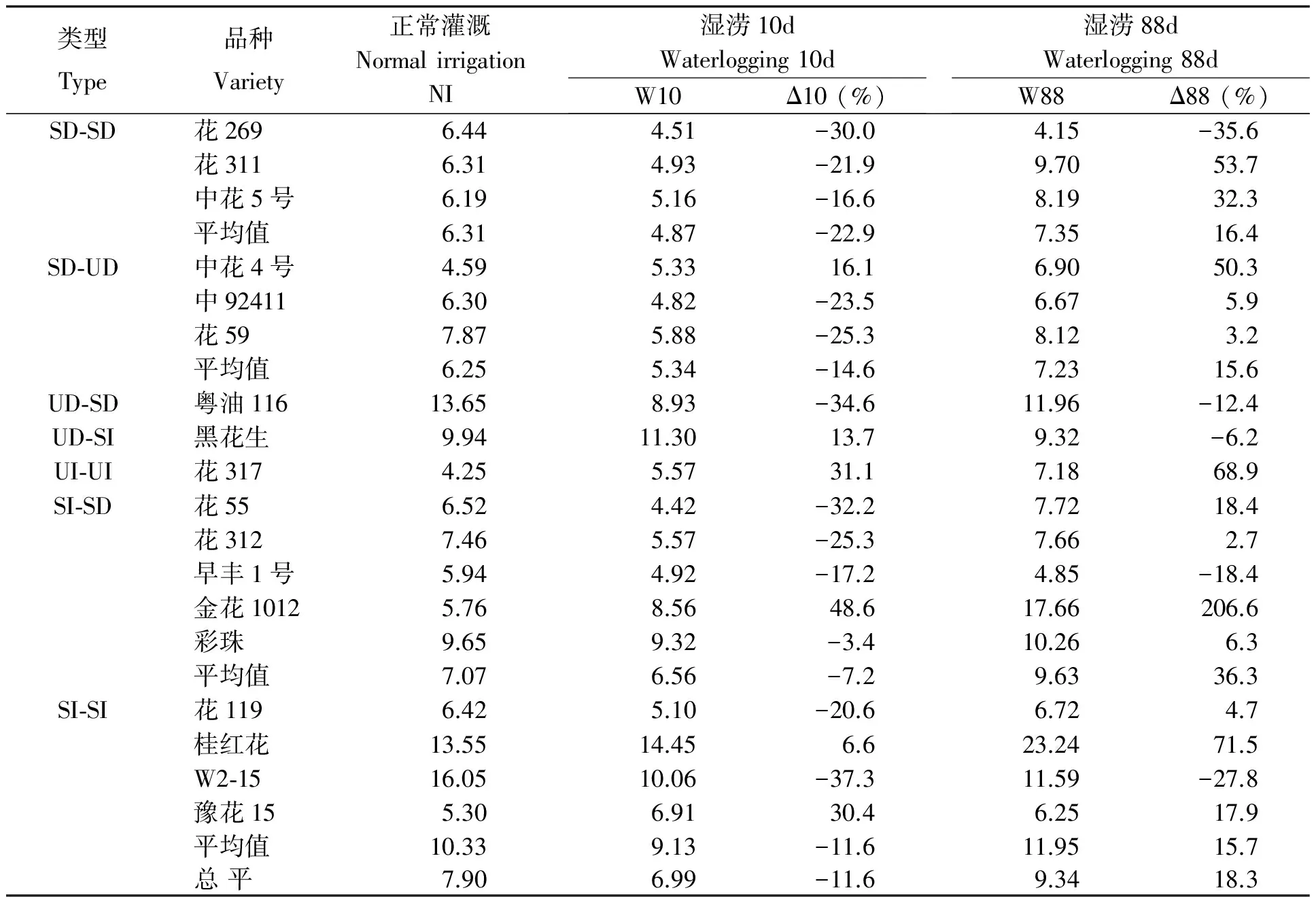
表2 湿涝对花生品种茎秆生物量的影响

表3 湿涝条件下茎秆生物量与耐湿涝系数的相关性
注:WTC:耐湿涝系数;AY:实际产量。 下同。
Note: WTC: waterlogging tolerance coefficient; AY: actual yield. The same as below.
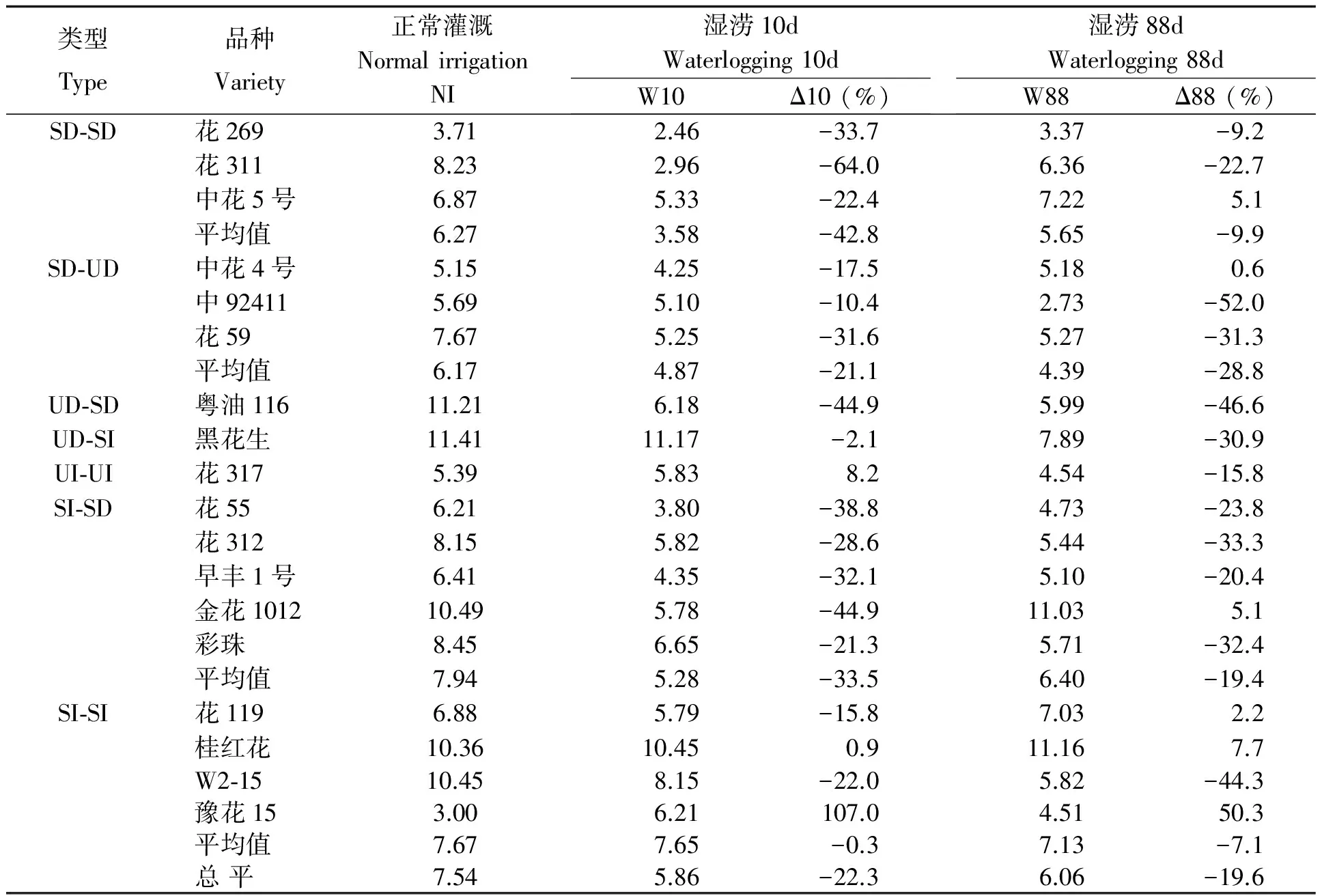
表4 湿涝对花生品种叶片生物量的影响
表2看出,短涝导致多熟花生品种茎秆生物量下降,降幅超过30%的品种有W2-15、粤油116、花55、花269(后三者有亲缘关系),而金花1012、花317、豫花15增幅在30%以上。SI-SI、SI-SD类型的多数品种在正常和湿涝时的茎秆生物量均高于SD-SD、SD-UD类型,且受湿涝影响较小。长涝茎秆生物量却普遍高于正常水分处理。其中金花1012增幅超过2倍,桂红花、花317、花311、中花4号增幅在50%以上。而花269、W2-15、早丰1号、粤油116减幅较大。类型间差异无明显规律性。相关分析表明(表3),短涝茎秆生物量高的品种一般表现出耐性较强而实际产量较低;长涝时各项指标相关程度均较低。
2.1.2 叶片生物量
短涝造成绝大部分品种叶片生物量下降(见表4),花311、粤油116、金花1012降幅逾40%,而豫花15增长1倍多,花317、桂红花略有增长。SI-SI、SI-SD类型叶片生物量较大,尤其SI-SI类型受湿涝的负效应小、正效应大。叶片生物量受长涝的影响反而小于短涝,中92411、粤油116的叶片生物量降幅逾40%,桂红花、中花5号、金花1012、花119、中花4号略增,而豫花15大幅上升。类型间差异无明显规律性。相关分析显示(表5),短涝后叶片生物量大特别是增幅大的品种,往往耐涝性强、实际产量较高;长涝叶片生物量及其变率与耐湿系数有较低正相关。
2.1.3 根系生物量
花生根系的湿涝适应性因品种而异(表6)。根系生物量表现明显不耐短涝的品种有花55、花312、金花1012、粤油116和花269、中花5号;耐的有花119、豫花15、花317、黑花生、中花4号和花59。长涝时多数品种根系生物量增加,其中花119和中花4号超过一倍。SI-SI、SI-SD类型多数品种湿涝时根系生物量大于SD-SD、SD-UD类型。相关分析显示(表7),短、长涝期时根系生长量高且增幅大对短期湿涝耐性和产量的贡献大,而与长期湿涝耐性和产量关系不大。
2.1.4 荚果生物量
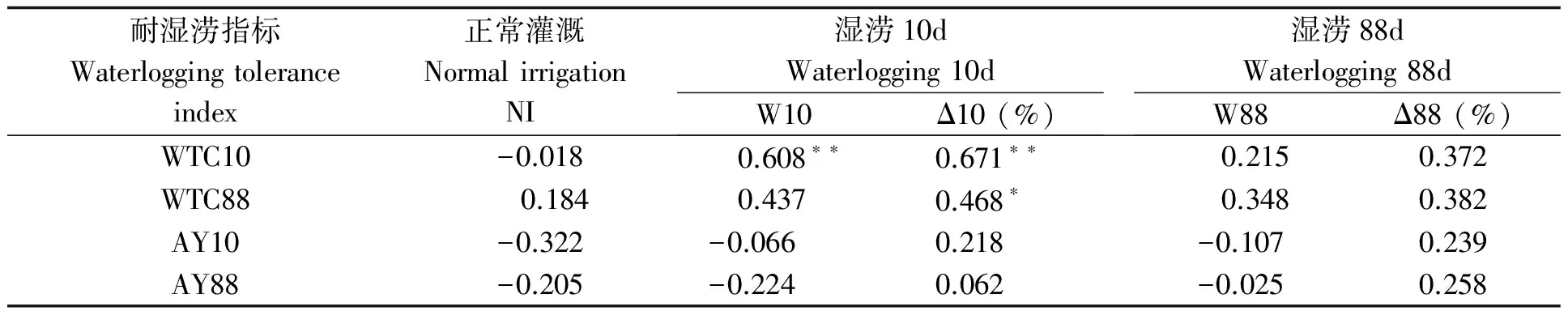
表5 湿涝条件下叶片生物量与耐湿涝系数的相关性

表6 湿涝对花生品种根系生物量的影响
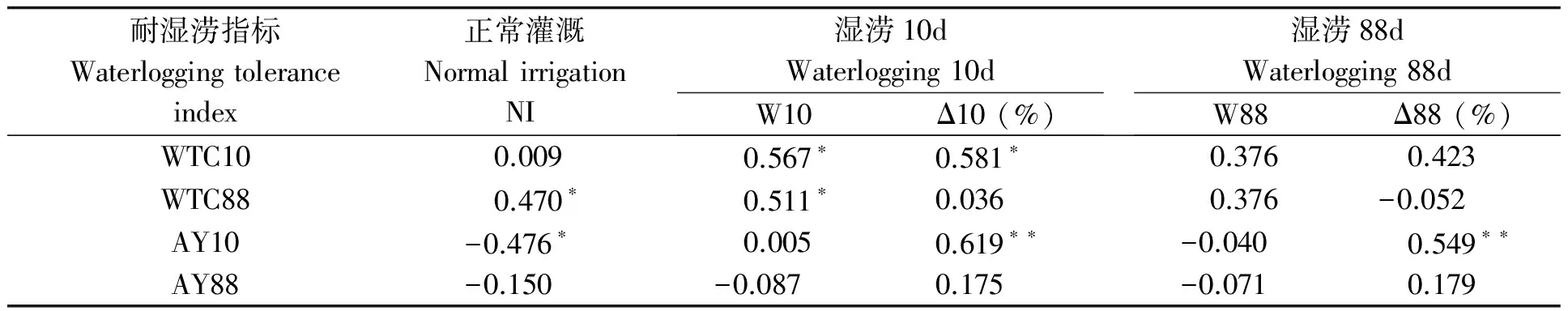
表7 湿涝条件下根系生物量与耐湿涝系数的相关性
短涝导致品种间荚果生物量差异较大(表8),其中花311、金花1012和花269降幅最大,而豫花15、彩珠增幅最大。SI-SI、SI-SD类型多熟品种荚果生物量低于SD-SD、SD-UD类型,SI-SI类型所有品种的荚果生物量因湿涝而大幅增加, SD-SD类型所有品种因湿涝而显著下降。在长涝时大多数品种的荚果生物量增加,豫花15增加近2倍,桂红花、彩珠和金花1012增长63.3%~92.2%,但花269减少27.4%。SI-SI、SI-SD类型湿涝荚果生物量增幅明显高于SD-SD、SD-UD类型。相关分析表明(表9),湿涝后荚果生物量大的品种多显示出实际产量高而耐性系数较低。

表8 湿涝对花生品种荚果生物量的影响
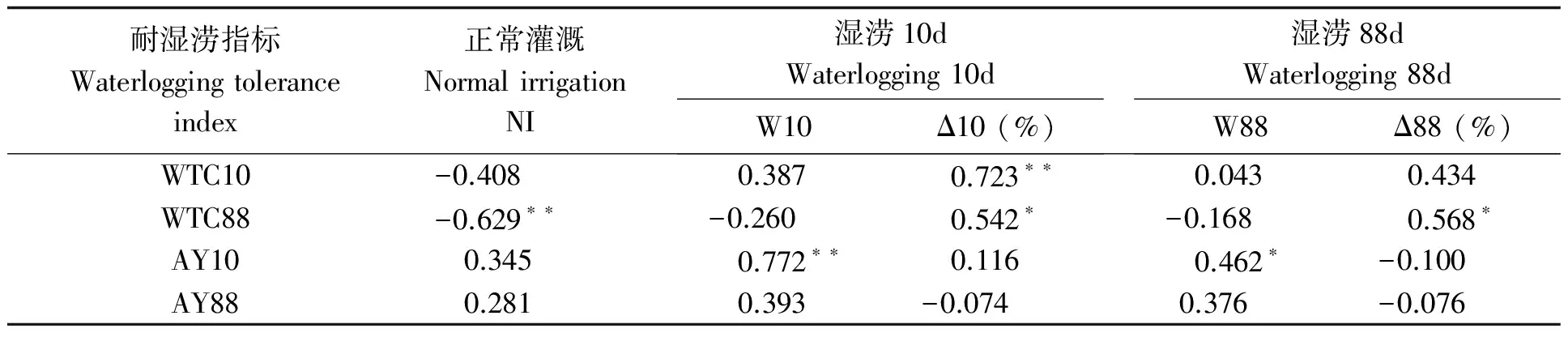
表9 湿涝条件下荚果生物量与耐湿涝系数的相关性
2.2 干物质分配比例对湿涝响应以及与耐性系数关系
2.2.1 各器官的干物质分配比例
总体而言(表10),正常水分时所有品种不同器官生物量比例(%)平均为茎∶叶∶根∶果为27.7∶26.7∶3.5∶42.1,在短、长涝时调整为28.0∶23.2∶4.1∶44.8和29.4∶19.4∶3.5∶47.7,说明随着湿涝期延长,茎秆和荚果(荚果比重实为经济系数)所占整株生物量的比重加大,叶片比重明显下降,而根系所占比重因短涝而上升,又因长涝而下降。相关分析表明(表11),茎秆、叶片、根系比重大的品种短、长涝耐性系数均较高,而产量偏低,荚果比重大的品种耐性系数较低,而产量较高,因此矮壮应成为高产品种的主体株型;叶片增重对短涝耐性尤为重要,根系、叶片生物量比重高是维持长涝耐性的基础。

表10 湿涝对花生品种各器官干物质分配比例的影响 (%)
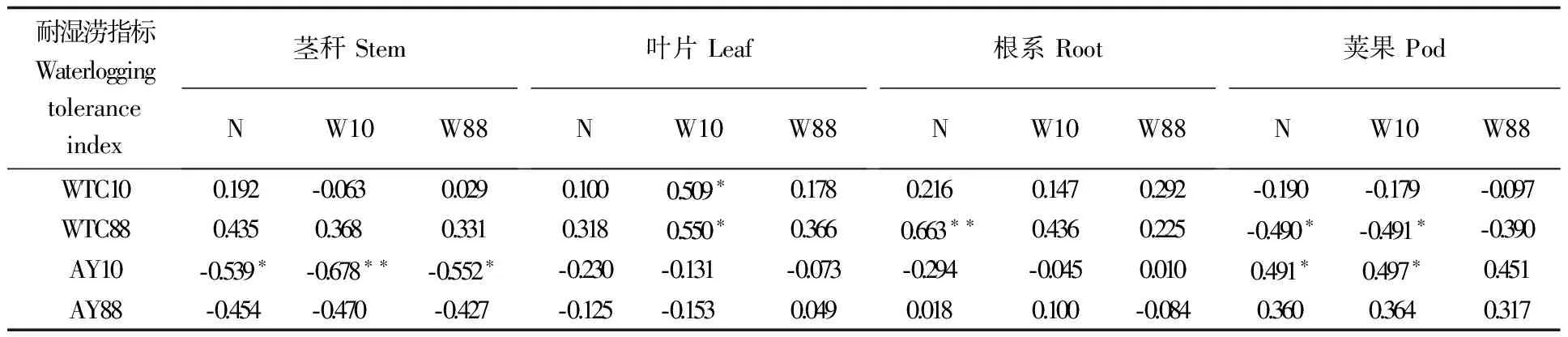
表11 不同器官干物质比例与耐湿涝系数的相关性 (%)
2.2.2 根冠比
湿涝时多数品种根冠比增加(表12),短涝下更明显,花311的根冠比增长近1倍,花119、中花4号、花59增幅超过75%,而花55、豫花15、金花1012有所下降。长涝后,花59、花119、黑花生的根冠比增长明显。SD-SD、SD-UD类型在湿涝时根冠比及其增幅均高于SI-SI和SI-SD类型。相关分析显示(表13),湿涝时根冠比较高的品种实际产量较高,而与耐性系数相关度低。
3 讨 论
3.1 耐湿涝品种的干物质积累与分配特征
土壤水分过多对植物发育和干物质积累分配有深刻影响[18-19]。本研究表明,短涝导致所有花生品种叶片生物量平均降低22.3%,茎秆降低11.6%,荚果略降低6.5%,而根系增重2.7%;长涝导致叶片生物量降低19.6%,而根系、茎秆、荚果分别增重9.4%、18.3%和26.3%。总干物质在不同器官中的分配比率也相应变化,在短、长涝条件下叶片比率分别下降3.5%和7.3%,茎秆上升0.3%和1.7%,根系上升0.6%和持平,荚果上升2.7%和5.6%,根冠比上升23.0%和11.1%。说明光合器官叶片对湿涝最敏感,其次是茎秆,荚果受短涝影响小且因长涝而增产,而根系具有稳定耐性。一些品种长涝时反而增产,可能包括几个方面缘由:一是虽然叶片生物量下降,但净光合速率可能提高;二是湿涝环境下光合作用对产量建成的贡献下降了,而地下器官(包括根系和荚果)直接参与物质吸收合成的比重可能提高了;三是叶片中的光合产物向荚果转运的速率和总量增加了。综合来看,短、长涝期时实际产量较高的品种一般表现出矮秆、叶多、根系发达、根冠比较高,而耐性系数多偏低,短、长涝时叶片与根系增重可实现耐湿与高产的统一,根系、叶片生物量比重高是维持长涝耐性的基础。
3.2 花生不同渍涝类型的防控对策
本试验表明,不同品种之间耐涝性差异很大,有的甚至会增产。因此,在涝害严重的地区可选用豫花15、花119、彩珠、桂红花、金花1012等耐渍涝品种;从类型间,可选用SI-SI类型。
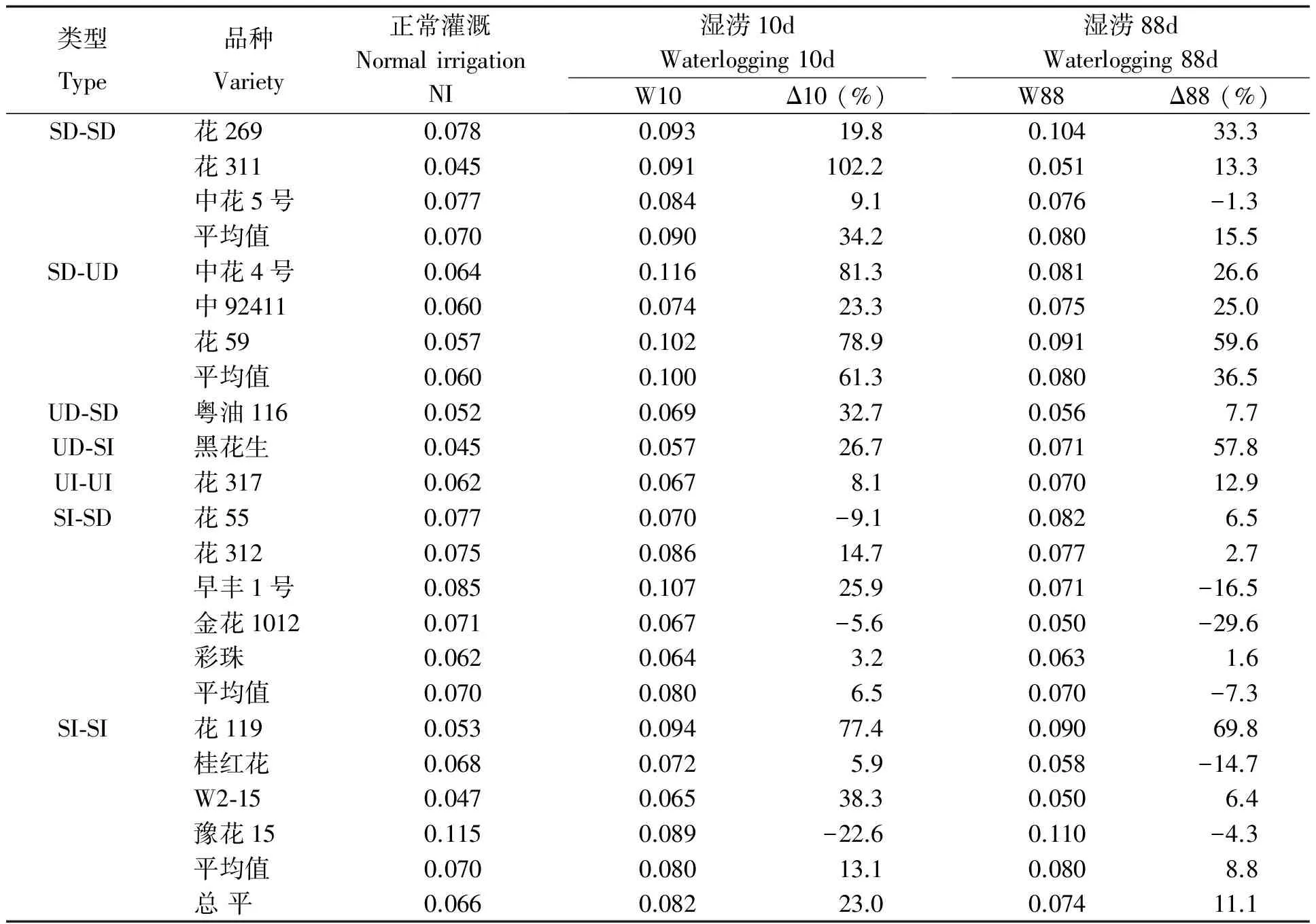
表12 湿涝对花生品种根冠比的影响
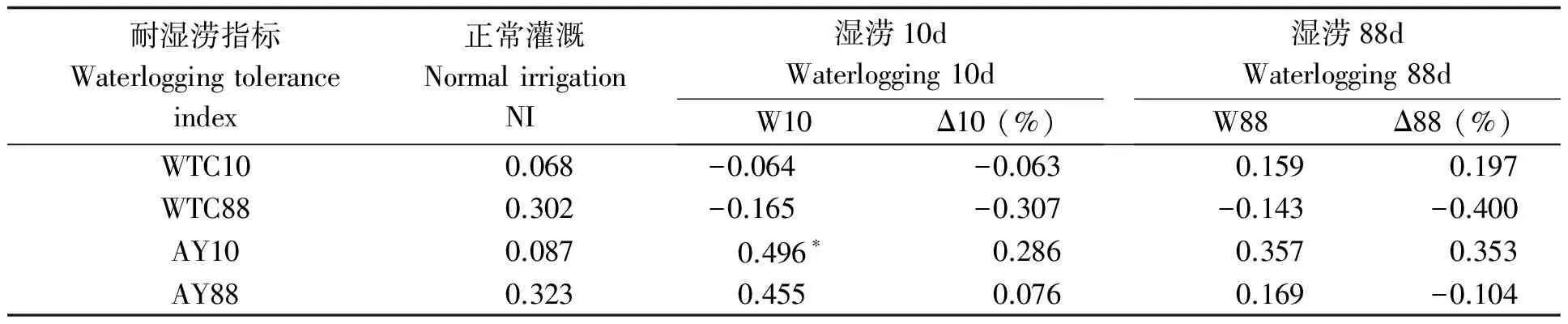
表13 湿涝条件下根冠比与耐湿涝系数的相关性
邱柳等[20]研究结果表明,在无流水的培养钵中,随着渍涝时间的延长,花生受伤害越大,尤其是根系,地上部次之。值得注意的是,本试验发现长涝时多数品种的根系、荚果生物量增加。其原因可能是采用活水进行渍涝处理,溶解氧含量较高,加之土壤渗水性强,加快了氧气的运输,可以支持花生根系的有氧呼吸,不至于大量无氧呼吸产生酒精毒害,还可能与长涝环境下耐性机制的巨大改变有关。因此,在花生实际生产中遭受洪涝灾害时,应根据水分状况采取相应对策:若是流动、清亮的洪水,短期淹涝对花生危害不大,而死水且浑浊则危害甚大,应设法尽快排涝。由此推测,未来花生采取无土水培时,若满足活水、暗环境、营养全面等必要条件,应该是高效的。
[1] 秦利, 韩锁义, 刘华. 我国食用花生研究现状[J]. 江苏农业科学, 2015(11):4-7.
[2] 石明权, 王晓林, 陈宏,等. 缺乏水分对花生生长发育、栽培的影响[J]. 陕西农业科学, 2008, 54(6):79-80.
[3] 曾红远. 耐渍花生生育生理对不同耕种模式的响应[D]. 长沙:湖南农业大学, 2013.
[4] 张翔, 张新友, 张玉亭,等. 施氮量对不同花生品种生长及干物质积累的影响[J]. 花生学报, 2011, 40(1):23-29.
[5] 刘登望, 王建国, 李林,等. 不同花生品种对旱涝胁迫的响应及生理机制[J]. 生态学报, 2015, 35(11):3817-3824.
[6] 刘吉利, 王铭伦, 吴娜,等. 苗期水分胁迫对花生产量、品质和水分利用效率的影响[J]. 中国农业科技导报,2009,11(2):114-118.
[7] 杨伟强, 宋文武, 鞠倩,等. 不同类型花生品种干物质积累特性研究[J]. 山东农业科学,2009(1):47-49.
[8] 褚田芬, 朱金庆, 徐明时,等. 土壤湿涝对春玉米前期生长的影响[J]. 浙江农业学报, 1995(4):39-41.
[9] 李林, 刘登望, 熊璟,等. 花生生育早期耐涝性室内鉴定对大田期的意义[J]. 作物学报, 2008, 34(3):477-485.
[10] 张岁岐, 山仑, 薛青武. 氮磷营养对小麦水分关系的影响[J]. 植物营养与肥料学报, 2000, 6(2):147-151.
[11] 骆兰平, 于振文, 王东,等. 土壤水分和种植密度对小麦旗叶光合性能和干物质积累与分配的影响[J]. 作物研究,2011,37(6):104-105.
[12] 于舜章, 陈雨海, 周勋波,等. 冬小麦期覆盖秸秆对夏玉米土壤水分动态变化及产量的影响[J]. 水土保持学报, 2004, 18(6):175-178.
[13] 杨京平, 陈杰. 受渍条件下稻田玉米产量与渍害指标的关系[J]. 浙江大学学报,2001,27(3):349-351.
[14] 杨璐, 曹卫东, 白金顺,等. 翻压二月兰对玉米干物质积累和养分吸收及土壤养分的影响[J]. 华北农学报,2014,29(1):183-189.
[15] 丁成伟, 华希新, 钮福祥,等.甘薯品种资源耐涝性鉴定研究[J]. 作物品种资源,1996(2):32-34.
[16] 李林, 邹冬生, 刘登望,等. 基于产量的花生基因型耐湿涝性综合评价[J]. 中国油料作物学报, 2004, 26(4):27-33.
[17] 李林. 花生品种间耐湿涝性差异及其机理研究[D]. 长沙:湖南农业大学, 2004.
[18] 向鹏华, 罗建新, 周万春,等. 土壤水分管理对烤烟生长发育及干物质与养分积累分配的影响[J]. 作物研究,2007,21(5):733-735.
[19] 骆兰平,于振文,王东,等.土壤水分和种植密度对小麦旗叶光合性能和干物质积累与分配的影响[J]. 作物研究,2011,37(6)103-105.
[20] 邱柳. 花生种质资源耐渍性鉴定研究[D]. 长沙:湖南农业大学, 2012.
EffectsofWaterloggingonDryMatterAccumulationandDistributioninPeanut
YI Jing1, LIU Deng-wang1,2, WANG Jian-guo1, ZHANG Hao1,2, GUO Feng3, WAN Shu-bo3*, LI Lin1,2*
(1.CollegeofAgriculture,HunanAgriculturalUniversity,Changsha410128,China; 2.UplandCropResearchInstitute,HunanAgriculturalUniversity,Changsha410128,China; 3.KeyLabofCropGeneticImprovementandEcologicalPhysiologyofShandong,Jinan2501100,China)
In order to determine the characteristics of dry matter accumulation and distribution characteristics of waterlogging-tolerant varieties and countermeasures for prevention and control waterlogging and flooding in peanut crops. 18 peanut varieties with different waterlogging tolerance were selected and treated with short-term waterlogging for 10 days (W10), long-term waterlogging for 88 days (W88) and normal irrigation (NI) during the end of vegetative growth stage in the field adapted to paddy and dry rotation (with strong soil water permeability). The effects of waterlogging on dry matter accumulation and distribution in different peanut varieties were studied. The results showed that the effects of short-term waterlogging on biomass of whole peanut varieties were obvious in leaf weight, and stem, legume, and root was also increased slightly. Long submergence reduced leaf biomass while promoted the development of root system, stem and pod, which showed that leaf as photosynthetic organ was most sensitive to waterlogging, followed by stem and pod which increased yield due to long-term waterlogging rather than short waterlogging and the root system had stably tolerant. The high yield varieties in short and long term waterlogging had the characteristics of generally dwarf with many leaves, well-developed roots and relatively high root-shoot ratio, however, the tolerant coefficient was lower. The increase in weight of leaves and roots during short and long waterlogging could achieve via the unity of waterlogging resistance and high yield. The high proportion of root and leaf biomass was the basis of maintaining long waterlogging tolerance. The varieties of Yuhua15, Hua119, Caizhu, Guihuahong, and Jinhua1012 were screened out which had high resistant of waterlogging in our study.
peanut(ArachishypogeaeL.); waterlogging; dry matter biomass; yield; tolerance coefficient
10.14001/j.issn.1002-4093.2017.03.007
S565.2; S332.1
A
2017-06-09
国家科技支撑计划项目(2006BAD21B04、2009BADA8B03、2014BAD11B04);国家现代农业产业技术体系(CARS-14);湖南省研究生科研创新项目(CX2015B230)
易静(1992-),女,湖南岳阳人,湖南农业大学农学院在读硕士,主要从事花生生理生态研究。
*通讯作者:李林,教授,博士生导师,主要从事花生栽培育种和农业生态研究。E-mail: lilindw@163.com 万书波,研究员,博士生导师,主要从事花生栽培生理研究。E-mail: wansb@ saas.ac.cn
猜你喜欢
种子(2023年8期)2023-11-15 10:42:20
江西农业大学学报(2022年3期)2022-07-07 04:00:52
快乐语文(2020年26期)2020-10-16 03:37:00
草地学报(2019年2期)2019-05-31 03:33:44
农机化研究(2019年9期)2019-05-24 09:53:28
农业机械学报(2019年4期)2019-04-29 02:20:48
畜牧与饲料科学(2018年5期)2018-06-13 12:03:48
食品工业科技(2014年15期)2014-03-11 18:17:33
土壤与作物(2013年3期)2013-03-11 18:47:31
植物营养与肥料学报(2010年4期)2010-11-06 07:28:54
