
拔节期光强对水稻抗倒伏能力的影响及机理
2017-12-01 09:50李晓娟梁开明钟旭华彭少兵黄农荣潘俊峰刘彦卓
华南农业大学学报 2017年6期
李晓娟,梁开明,钟旭华,田 卡,彭少兵,黄农荣,潘俊峰,刘彦卓
(1 广东省水稻育种新技术重点实验室/广东省农业科学院 水稻研究所,广东 广州 510640;2 华中农业大学 植物科学技术学院,湖北 武汉 430070)
拔节期光强对水稻抗倒伏能力的影响及机理
李晓娟1,2†,梁开明1†,钟旭华1,田 卡1,彭少兵2,黄农荣1,潘俊峰1,刘彦卓1
(1 广东省水稻育种新技术重点实验室/广东省农业科学院 水稻研究所,广东 广州 510640;2 华中农业大学 植物科学技术学院,湖北 武汉 430070)
【目的】探讨拔节期光强对水稻Oryza sativa L.抗倒伏能力的影响。【方法】通过人工气候箱光强控制试验,在基部第1、第2、第3节间伸长期及第1~3节间伸长期进行不同光强处理,测定水稻茎秆倒伏指数和基部节间的形态和材料力学性状。【结果】倒伏指数随着光强增加显著下降。在第1~3节间伸长期连续用中、高光强处理,茎秆倒伏指数比低光强处理降低34.8%~73.9%。倒伏指数与节间长度呈极显著正相关,与节间粗度、茎壁厚度和节间充实度呈显著负相关。第2节间伸长期是茎秆抗倒伏能力对光强反应最敏感的时期。光强对水稻抗倒伏能力的影响有累积效应。第1~3节间伸长期连续处理条件下,倒伏指数和多数形态、材料力学性状的变异系数高于单个节间伸长期处理。【结论】节间长度、茎壁厚度和节间充实度是影响茎秆抗倒伏性的关键因子。光强改变了节间长度、单位体积节间干质量和茎壁厚度等形态特性,进而影响茎秆抗折力和弯曲力矩等材料力学性状,最终影响水稻抗倒伏能力。
水稻;倒伏指数;光强;形态性状;材料力学性状
倒伏是限制水稻Oryza sativa L.高产的重要因子之一,可导致10%~30%的减产甚至绝产,同时导致稻米品质下降,增加收割成本[1-2]。倒伏分为根倒伏和茎倒伏2种。根倒伏多数发生在旱稻和直播稻中,是由于根系入土浅、固持能力差引起的;茎倒伏则多发生在移栽稻中,是由于茎秆基部节间折断引起的。华南地区以移栽稻为主,水稻倒伏主要是茎倒伏。特别是早稻,由于多雨少日照,台风暴雨频繁,倒伏问题更为严重。水稻的抗倒伏能力由茎秆形态和材料力学特性决定。茎秆形态性状包括株高、重心高度、基部节间长度、茎壁厚度、节间粗度、节间充实度和维管束数目等,而材料力学性状包括机械强度、抗折力、弹性模量、弯曲力矩和抗弯刚度等[3-5]。株高是影响水稻抗倒伏能力的重要形态性状[6]。20世纪60年代,矮秆品种的育成和应用大幅降低了水稻株高,提升了水稻抗倒伏能力,水稻产量也随之上了一个台阶。但是,随着水稻产量的持续提高,水稻的株高逐步增加,倒伏风险也随之加大,倒伏问题再次成为水稻生产的一大难题。因此,如何在株高适度增加的情况下,通过改善茎秆的形态和材料力学特性,提高水稻抗倒伏能力,实现抗倒伏与高产的协调,已成为国内外的研究热点[4-6]。水稻茎秆一般由4~7个伸长节间组成,而茎倒伏的折断部位为基部伸长节间。改善基部节间的形态和材料力学性状,可有效防止倒伏的发生[3,7-8]。光照是影响水稻抗倒伏能力的重要因子。许多栽培措施(如密度、施肥和水分管理等)对水稻抗倒伏能力的影响都与群体内部光环境的改变有关。移栽密度过高,导致群体基部光照不足,基部节间非正常伸长,茎秆纤弱,抗折力降低; 反之,低密度栽插改善了水稻群体基部的光照条件,有利于茎秆的个体发育,从而提高茎秆抗倒伏能力。茎秆基部节间倒伏指数与栽插密度显著正相关,栽插密度增加会使基部节间长度增加,茎壁厚度和茎粗降低,最终导致倒伏指数增大[9]。过量施用氮肥,导致群体叶面积过大,基部受光少,基部节间长度增加,节间充实度下降,倒伏指数增加[10-13]。张明聪等[14]研究发现,优化氮肥运筹可以缩短水稻基部各个节间长度,增加茎粗和充实度,从而显著增强水稻的抗倒伏能力。在干湿交替和控水灌溉模式下,水稻无效分蘖得到控制,群体透光率增加,倒伏指数明显低于连续淹水模式[15-16]。光强是如何影响水稻抗倒伏能力的?其形态机理和材料力学机理如何?不同节间伸长期对光强处理的敏感性有无差异?是否存在累积效应?目前尚不清楚。弄清这些问题对水稻抗倒高产栽培具有重要指导意义。关于光强对水稻抗倒伏能力的影响,已有的研究大多是在田间自然条件下进行的,主要通过设置不同的施肥水平和栽插密度,改变群体光环境,进而探讨群体内光强与抗倒伏能力的关系。但是,施肥和栽插密度的改变不仅改变了群体光环境,同时也改变了植株含氮量等一系列生理生化指标。因此,有必要开展光强控制试验,在排除其他因素干扰的情况下,进一步探讨光强对水稻抗倒伏能力的影响及机理。本研究通过人工气候箱光强控制试验,在不同节间伸长期设置不同光强环境,探讨光强对水稻抗倒伏能力的影响及材料力学机理,以期为水稻抗倒高产栽培调控提供理论依据。
1 材料与方法
1.1 材料和试验设计
试验于2014年早季在广东省农业科学院水稻研究所的网室和人工气候箱内进行。供试水稻品种为银晶软占,属感温型常规稻品种,株高101~105 cm,早季全生育期125~128 d,5个伸长节间,主茎总叶片数为15片。试验材料采用盆栽方式种植于网室,培养钵为封底的PVC管,管高和内径分别为22和12 cm。每盆装过筛(10目)的水稻干土1.9 kg。土壤的主要理化性状为:pH 5.98,有机质、全氮、全磷和全钾分别为 21.5、1.19、1.11 和 11.3 g·kg–1,碱解氮、有效磷和速效钾分别为 88.5、43.9 和 45.2 mg·kg–1。2014年3月14日播种,育秧盘育秧,4月15日(5叶期)选取生长均匀的秧苗移栽,每盆1穴,每穴1苗。在水稻生长过程中各处理的水肥管理保持一致,每盆施肥总量为:尿素0.94 g、氯化钾0.90 g、过磷酸钙0.42 g、七水硫酸锌0.025 g。磷肥、钾肥和锌肥均作基肥在移栽前1 d一次性施入,氮肥分2次施入,移栽前1 d施2/3,11叶期施1/3。整个生育期保持浅水层灌溉,及时防治病虫害。为准确记录叶龄,移栽后每3 d标记1次叶龄,接近拔节时每天观测叶龄。根据叶茎生长同伸规律,以叶龄确定各节间伸长的起止时间。在基部特定节间伸长期,选取长势和叶龄一致的20 盆材料移入Conviron PGs–36型人工气候箱内,进行不同光强处理,具体方法参照文献[17-18]。试验期间去除分蘖,只保留主茎,防止因分蘖数量和大小不同而改变叶面积和基部光环境。试验设置4个光照度水平和4个处理时期,光照度水平分别为 100、200、400 和 750 μmol·m–2·s–1,依次用L100、L200、L400和L750表示。4个处理时期分别为:1) 第1节间伸长期(叶龄余数为3.0~2.0,用 S1表示);2) 第 2节间伸长期(叶龄余数 2.0~1.0,用 S2表示);3) 第 3节间伸长期(叶龄余数 1.0~0.0,用 S3表示);4) 第 1~3节间伸长期 (叶龄余数3.0~0.0,用S123表示)。人工气候箱内各处理的温湿度条件设置一致,相对湿度不低于75%,日平均温度 26 ℃(最高 31 ℃、最低21 ℃,按正弦曲线变温),光照时间为12.5 h (06:30—19:00)。
1.2 测定项目和方法
抽穗后15 d取12盆材料对其茎秆形态和材料力学性状指标进行测定。
1) 株高(h):测量地表到穗部顶端的长度(cm)。
2) 重心高度(H)和单茎鲜质量(M):将茎秆剪去地下部分后置于一个支点上,左右方向移动茎秆使其保持水平,支点到茎秆第1节间末端长度为重心高度(cm)。用天平称量单茎鲜质量(g)。
3) 节间长度(l)和节间鲜质量(w):从节点处剪断茎秆使其分成独立节间,用直尺测量茎秆基部向上第1、2、3节间长度(cm)并称量每个节间鲜质量(g)。
4) 节间粗度(d)和茎壁厚度(D):将节间从中部截断,用游标卡尺测量长轴和短轴外径,取平均值为节间粗度(mm),茎壁厚度为长轴和短轴与茎壁4个交点处的平均厚度(mm)。
5) 单位长度节间干质量(m1)和单位体积节间干质量(m2):将各节间分装,置烘箱于105 ℃杀青30 min,80 ℃下烘至恒质量并称量得节间质量(m);把节间看作一个近似空心圆柱体,计算节间体积 (V)。
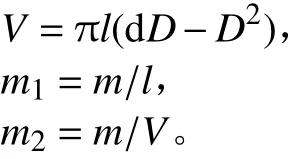
6) 弹性模量(E):采用YYD–1型茎秆强度测量仪(浙江托普仪器有限公司生产) 测定。将节间两端水平放置在2个支点上,2支点间距离为节间长度(l),在l/2处垂直施加力F使节间适当弯曲但不折断,然后测量l/2处的垂直位移δ。按以下公式计算弹性模量[19]:
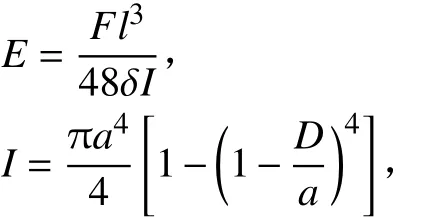
式中,I为截面惯性矩,a为节间半径(mm)。
7) 节间抗折力 (BR):用YYD–1 型茎秆强度测量仪(浙江托普仪器有限公司生产)测定。固定基部节间2个支点间距离为4 cm,将节间水平放置在2个支点上,在节间中点施力使其折断,力的大小即为该节间抗折力(N)。若节间长度小于4 cm则不测。
8) 抗弯刚度(BS):抗弯刚度为弹性模量和截面惯性矩的乘积,即
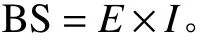
9)倒伏指数(LI):根据濑古秀生[20]的公式计算,即
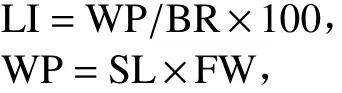
式中,WP为全株加在基部节间的弯曲力矩(g·cm),SL为基部节间折断部位到穗顶的距离(cm),FW为基部节间折断部位至穗顶的鲜质量(g)。
1.3 数据处理
使用Statistix 8 for Windows进行方差分析和回归分析,LSD法进行多重比较。采用不同光强处理下各形态及材料力学性状的变异系数(CV)作为衡量各性状对光强反应敏感性的指标,CV=标准差/平均值×100%。
2 结果与分析
2.1 不同光强处理对水稻茎秆抗倒伏能力的影响
由表1可知,不论在哪个节间伸长期间进行光强处理,倒伏指数均有随着光强增加而下降的趋势,表明改善节间伸长期间的光照条件,可以降低倒伏风险。在S1进行不同光强处理,倒伏指数随着光强的增加而下降,但差异未达到显著水平。在S2处理,不同光强处理的倒伏指数表现为L100> L200>L400,处理间存在显著差异。在S3处理,倒伏指数也随着光强的增加而显著减小。在S123处理,L200、L400光强处理下的倒伏指数比L100处理分别减小了34.8%和73.9%,降幅明显高于S1、S2和S3。从不同节间伸长期处理的倒伏指数变异系数(CV)来看,最高CV值出现S2,表明在第2节间伸长期进行光强处理对倒伏指数影响最大。在S123连续处理下,倒伏指数的CV值明显高于S1、S2和S3单个节间伸长期处理,表明光强对倒伏指数的影响具有累积效应。
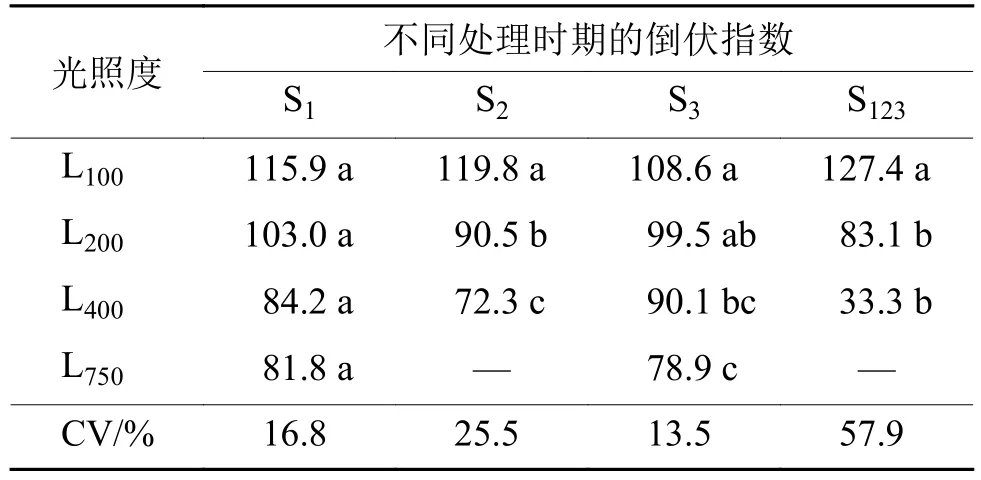
表1 不同光强处理下水稻茎秆的倒伏指数1)Tab. 1 The lodging index of rice stem under different light intensity treatments
以第2节间伸长期光强为自变量(x),倒伏指数(LI)为因变量(y),得到回归方程:y=–0.100x+120.7,r=0.663,n=31。以第 1~3节间伸长期间光强为自变量(x),得到回归方程:y=–0.133x+135.8,r=0.647,n=40。2个回归方程都达到极显著水平(p<0.001),倒伏指数随节间伸长期间光强的增加而下降。
2.2 不同光强处理对水稻茎秆形态性状的影响
2.2.1 不同光强处理对株高和重心高度的影响 由图1可知,光强对株高和重心高度有显著影响。在S1处理,株高和重心高度在不同光强处理间无显著差异,而在S2、S3和S123处理,株高和重心高度均随着光强的增加而下降。其中在S123处理,L750处理的株高和重心高度分别比L100处理降低了7.4%和26.9%,差异达到显著水平。此外,在S123连续处理,株高和重心高度的CV均明显高于S1、S2和S3单个节间伸长期处理。
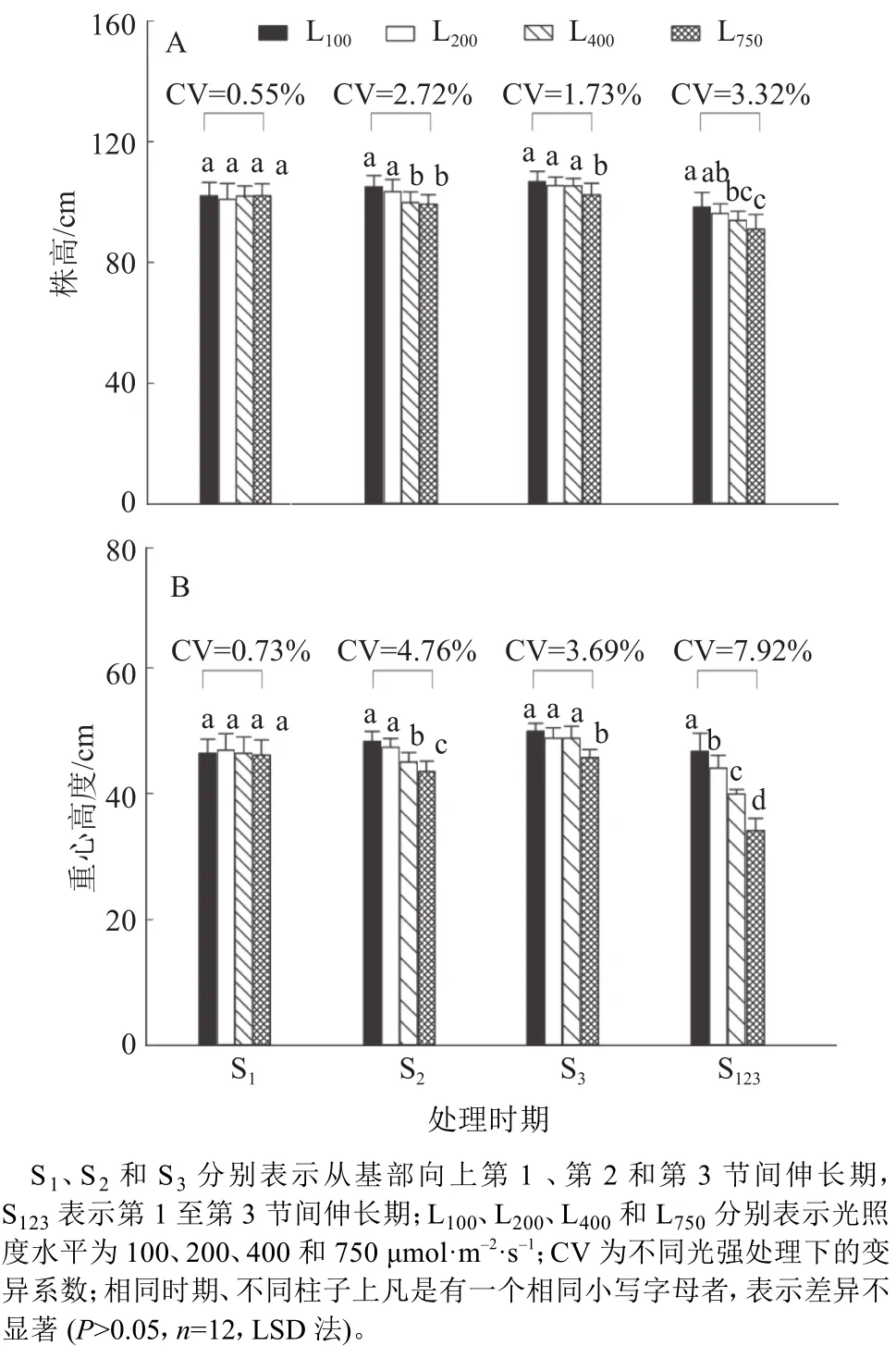
图1 不同光强处理的水稻株高和重心高度Fig. 1 The plant height and gravity center height of rice under different light intensity treatments
2.2.2 不同光强处理对基部节间长度、茎壁厚度和节间粗度的影响 由表2可知,在S1处理,第1节间长度随着光强的增加而显著缩短,第2节间长度在不同光强处理间差异不显著,而第3节间则反而随着光强的增加而显著拉长。在S2处理,光强对第1节间长度无显著影响,第2节间长度随着光强的增加而显著缩短,而第3节间长度则显著拉长。在S3处理,光强对第1、2节间长度无显著影响,第3节间长度则随着光强的增加而显著缩短。在S123处理,第1、2节间长度都以L100最长,L750最短或接近最短,对第3节间长度则无显著影响。由此可见,在基部 3个节间伸长期间(S1、S2、S3、S123)进行光强处理,对正在伸长节间的长度具有显著影响,其节间长度随着光强的增加而显著缩短。但在S1和S2进行光强处理,随着光强的增加,第3节间长度不仅不缩短,反而拉长。在S1处理,基部第1、2节间的茎壁厚度均随着光强的升高而增加,但第3节间反而变薄。在S2处理,第1节间的茎壁厚度在不同光强处理间无显著差异,第2、3节间的茎壁厚度则随着光强的升高而增加。在S3处理,光强对第1、2节间的茎壁厚度无显著影响,而第3节间的茎壁厚度则以L100最薄。在S123连续处理,基部3个节间的茎壁厚度均随着光强的升高而显著增加。这表明,在各个节间伸长阶段,光强增加显著增加了正在伸长节间及其上位节间的茎壁厚度。光强处理对节间粗度亦具有显著影响,在某个节间伸长期进行处理,则该节间及其上位节间的粗度均随着光强的增加而增加。
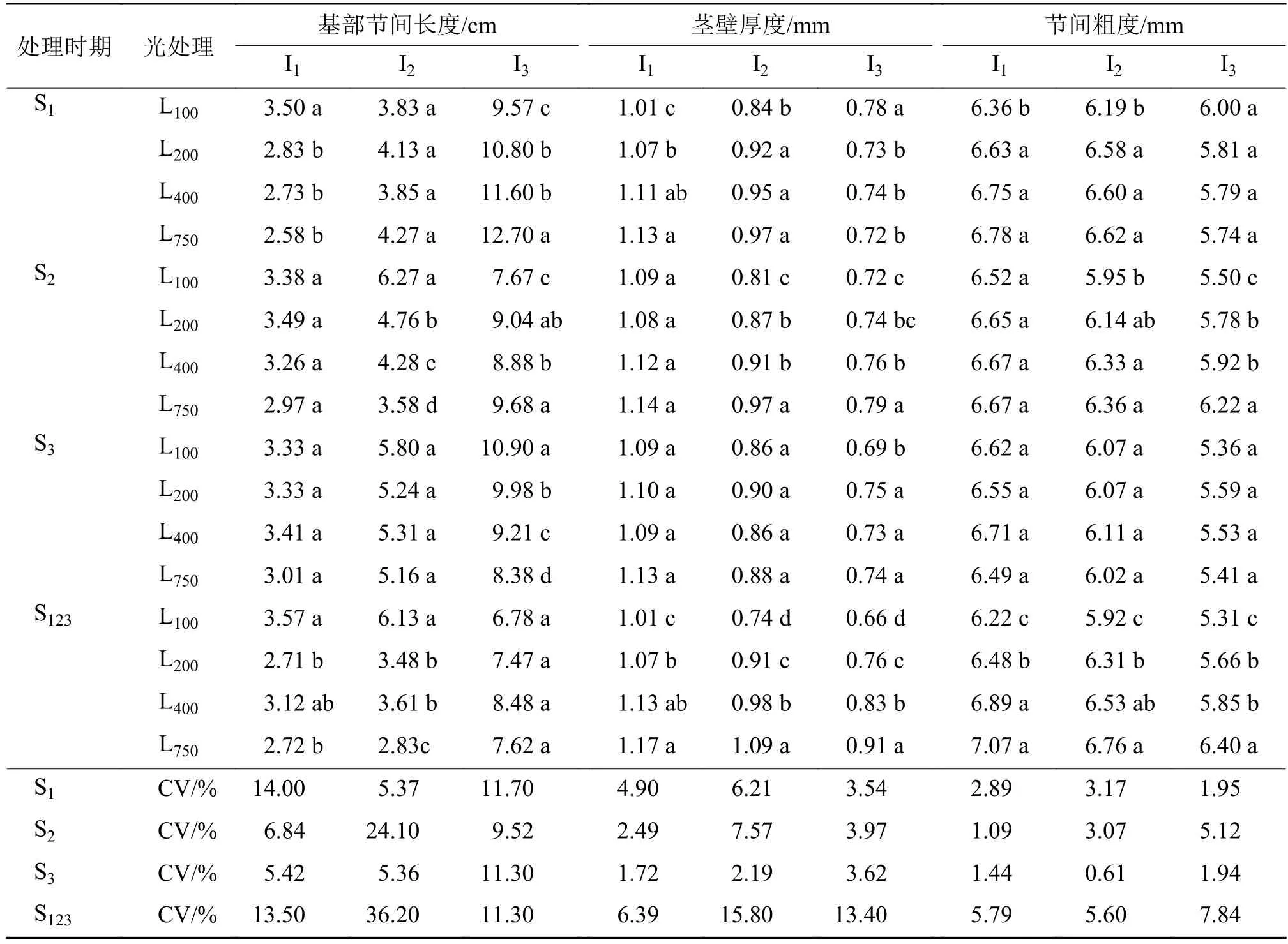
表2 不同光强处理下水稻植株基部节间长度、茎壁厚度和节间粗度1)Tab. 2 The basal internode length, culm wall thickness and basal internode diameter of rice plant under different light intensity treatments
基部节间长度的变异系数(CV)比较表明,CV的最高值和次高值分别出现在S123和S2处理,均以第2节间最高,说明第2节间长度对光强的反应比第1节间和第3节间更为敏感。茎壁厚度的CV在S1、S2和S123处理下多以第2节间最高。节间粗度的CV在S1处理以第2节间最高,而在S2、S3和S123处理则以第3节间最高,表明茎壁厚度对光强的反应以第2节间最为敏感,而节间粗度则以第3节间最为敏感。
2.2.3 不同光强处理对基部节间干质量和充实度的影响 由表3可见,光强处理显著影响了水稻茎秆的基部节间干质量(m)。在S1处理,对第2节间的m有显著影响,且以L200处理最高,而对第1、3节间的m无显著影响。在S2处理,随着光强的提高,基部3个节间的m均有不同程度的增加,处理间存在显著差异。在S3处理,光强对基部3个节间的m均有显著影响,且均以L400处理下最高。在S123处理,基部3个节间的m均以L400和L750较高,L100和L200较低。
单位长度节间干质量(m1)和单位体积节间干质量(m2)反映了茎秆基部节间的充实度高低。在S1处理,光强对第1、2节间的m1无显著影响,第3节间的m1则随着光强的增加而降低。在S2、S3和S123处理,基部3个节间的m1均以L400和L750较高,L100和L200较低,处理间存在显著差异。m2变化趋势与m1基本一致。上述结果表明,在S2、S3和S123处理,基部节间充实度有随着光强的升高而增加的趋势。光强处理不仅对正在伸长节间的充实度有显著影响,对其他节间的充实度亦产生显著影响。
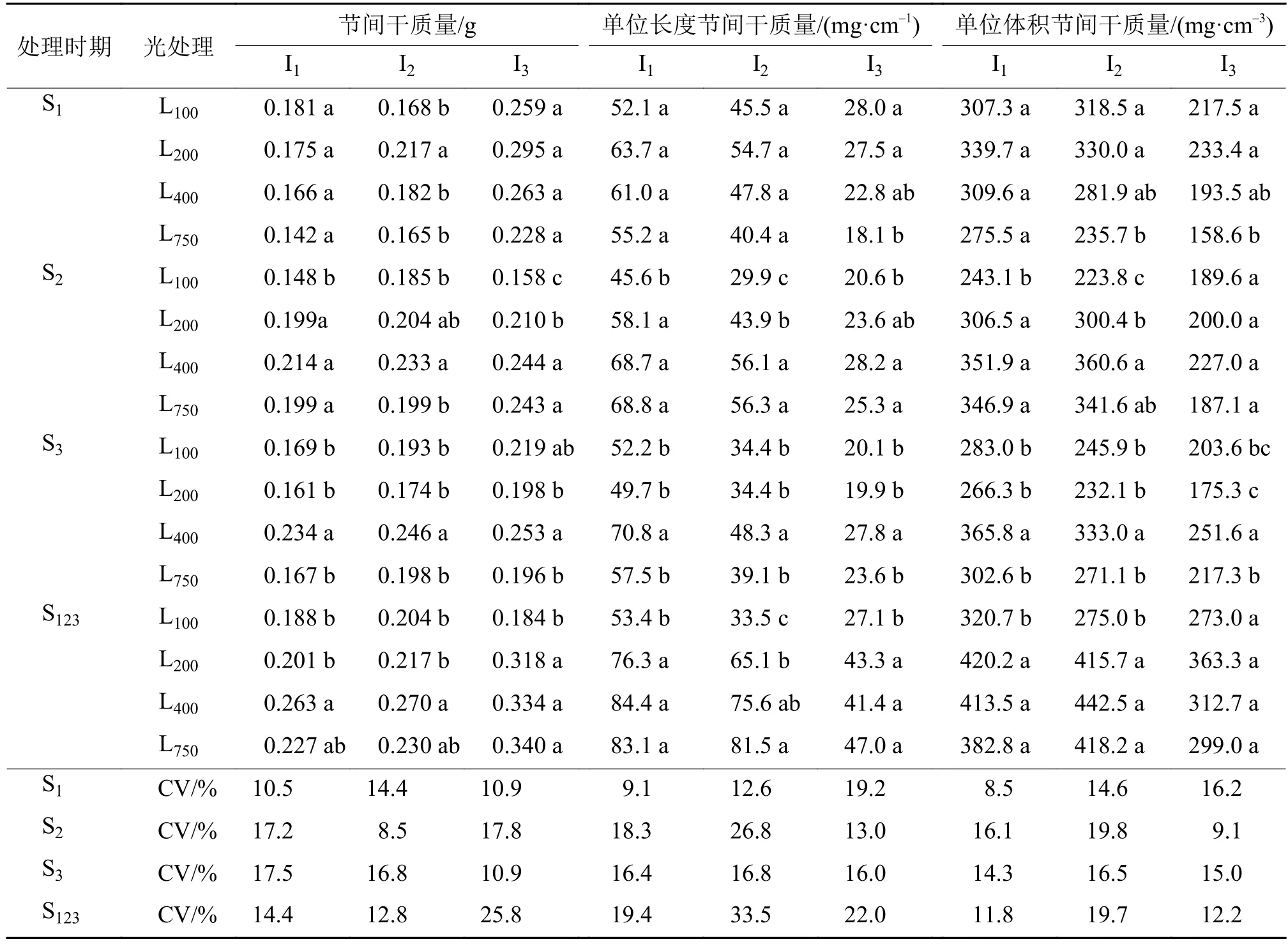
表3 不同光强处理的基部节间干质量、单位长度节间干质量和单位体积节间干质量1)Tab. 3 The dry weight, dry weight per unit length and dry weight per unit volume of basal internode under different light intensity treatment
不同光强处理间m1和m2的变异系数,多数以第2节间最高,表明在3个基部节间中,第2节间充实度对光强处理的反应最为敏感。
2.2.4 茎秆形态性状与倒伏指数的相关分析 相关分析表明,倒伏指数与节间长度呈极显著正相关(r=0.297,p<0.01),与重心高度呈显著正相关(r=0.210,p<0.05),与茎壁厚度、单位长度节间干质量和单位体积节间干质量呈极显著负相关(r分别为–0.249、–0.253 和–0.304,p<0.01),与节间粗度呈显著负相关 (r=–0.227,p<0.05),与株高、穗鲜质量和单茎鲜质量的相关性则未达到显著水平(r分别为0.166、0.146 和 0.145,P>0.05)。以倒伏指数为因变量(y),以茎秆形态性状为自变量,以P=0.05为变量入选和剔除条件进行逐步回归,得回归方程:y=235.46−195.451x1+10.495x2−0.103x3,相关系数 (r)=0.861(F=38.93,p<0.001),达极显著水平。式中 x1、x2和x3分别为茎壁厚度、节间长度和单位体积节间干质量。表明在不同光强处理下,影响水稻抗倒伏能力的关键性状是茎壁厚度、节间长度和节间充实度。
2.3 不同光强处理对水稻茎秆材料力学特性的影响
由表4可知,在第2和第3节间伸长期,光强增加提高了伸长节间的抗折力。在S1处理,光强对第2节间和第3节间抗折力未产生显著影响。在S2处理,第2节间抗折力随着光强的增加而显著提高,但对第3节间抗折力无显著影响。在S3处理,光强增加不仅提高了第3节间抗折力,同时也提高了第2节间抗折力。由变异系数可见,在S3处理,第3节间的抗折力变异系数较高,但在S1、S2期以及S123处理,抗折力的变异系数则以第2节间较高,即第2节间的抗折力对光强的反应较为敏感。
光强主要影响了第3节间的弹性模量,而对第2节间的弹性模量未造成显著影响。在S1、S2、S3处理和S123连续处理,第2节间的弹性模量在不同光强处理间的差异均未达到显著水平。在S1和S3处理,对第3节间弹性模量也无显著影响。但在S2和S123处理,L200、L400和L750处理下的第3节间弹性模量均显著低于L100处理。弹性模量在不同光强处理下的变异系数以第3节间较高,表明光强对第3节间弹性模量的影响大于第2节间。
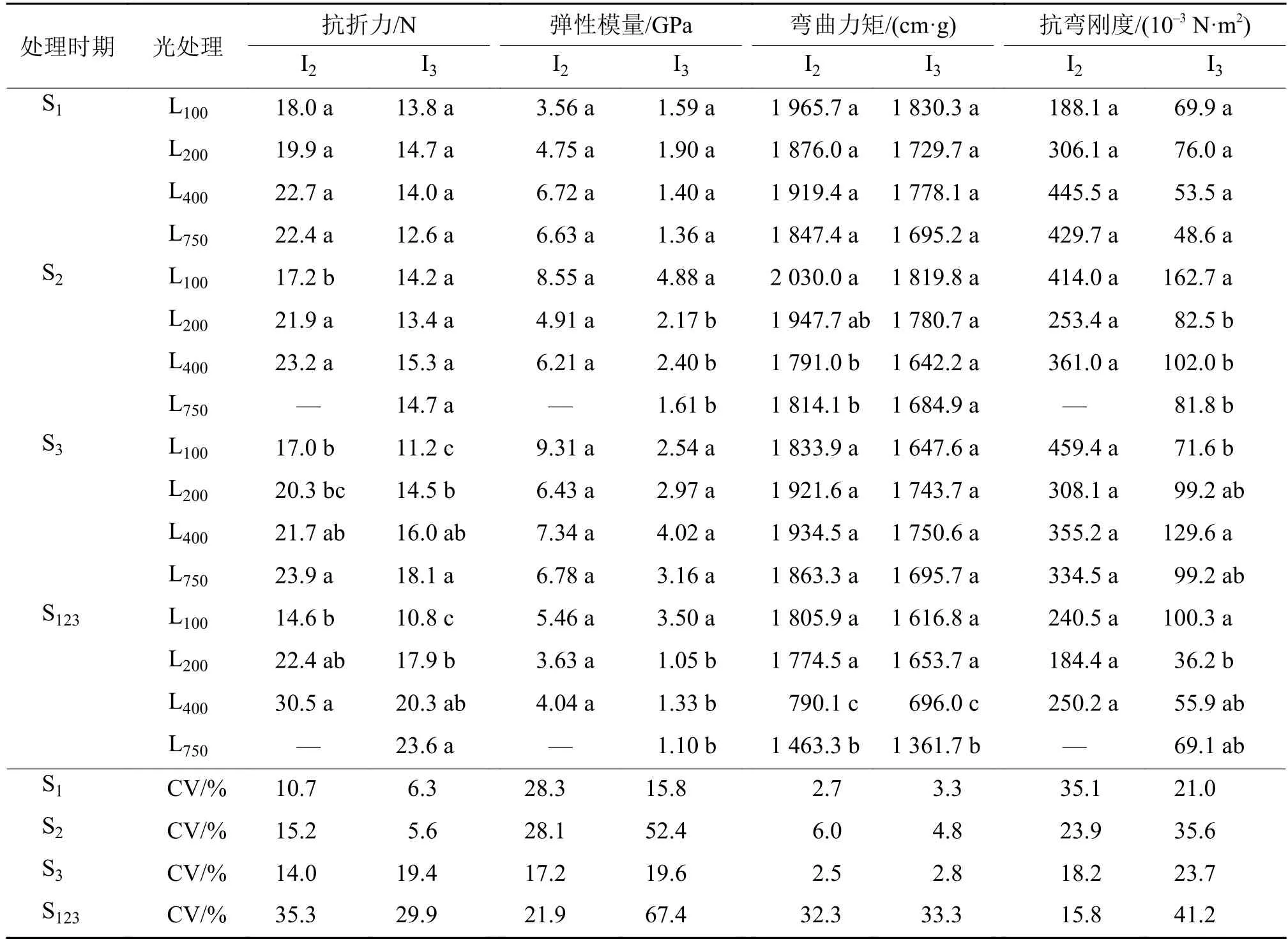
表4 不同光强处理的基部节间抗折力、弹性模量、弯曲力矩和抗弯刚度1)Tab. 4 The breaking resistance, elastic modulus, bending moment and bending stiffness of basal internode under different light intensity treatment
弯曲力矩测定结果表明,在第2和第3节间伸长期处理,光强的增加降低了伸长节间的弯曲力矩。在S2处理,随着光强的增加,第2节间和第3节间弯曲力矩均呈下降的趋势,其中L400和L750处理下的第2节间弯曲力矩分别比L100处理降低了11.8%和10.6%,差异达到显著水平。在S123处理,L400和L750处理下基部第2、3节间的弯曲力矩均显著低于L100和L200处理。
抗弯刚度对于光强处理的反应没有表现出明显规律性。对第2节间而言,在S1处理,其抗弯刚度随着光强的升高而增加,而在S2和S3处理,反而随着光强的升高而有下降趋势。对第3节间而言,在S3处理,L100处理的抗弯刚度低于其他光强处理,表明在第3节间伸长期,光强的增加提高了该节间的抗弯刚度,但在S2和S123处理,光强增加则降低了该节间的抗弯刚度。
相关分析表明,倒伏指数与节间抗折力呈极显著负相关 (r= – 0.377,p<0.01),与节间的弯曲力矩呈极显著正相关(r=0.958,p<0.01),与相应节间抗弯刚度和弹性模量相关性不显著(r分别为– 0.005和 0.091,P>0.05)。由此可知,在不同光强条件下,基部节间抗折力和弯曲力矩是影响水稻抗倒伏能力的主要因素,而弹性模量和抗弯刚度的影响不大。
3 讨论与结论
3.1 光强对水稻抗倒伏能力的影响
光强是影响水稻生长发育的重要因子之一,光强改变作物的株高、节间长度和茎壁厚度等形态性状,而这些变化将影响茎秆的抗倒伏能力[21-23]。关于光强对水稻抗倒伏能力的影响,国内外已有不少研究,但是这些工作大多是在田间条件下进行,通过设置不同栽插密度和施肥处理,改变群体叶面积指数和群体透光率,进而探讨群体内光强与水稻抗倒伏能力的关系。这种方法的优点是处理条件与生产实际接近,但不同栽插密度和施肥条件下,除群体基部光强外,光质也发生了变化[24],植株含氮量也可能受到影响[8-9]。本研究采用人工气候箱控制试验,避免了光强以外的因素干扰,系统地研究了光强对水稻抗倒伏能力及其相关性状的影响。研究结果表明,倒伏指数随光强的升高而显著下降。在第2节间和第1~3节间伸长期处理,L200和L400处理的倒伏指数比L100处理降低24.5%~73.8%,这与前人研究结果一致[8,25-27]。在水稻生长过程中,增加光强可抑制基部节间的伸长,提高茎秆抗倒伏能力[25-26]。
3.2 光强影响水稻抗倒伏能力的机理
本研究结果表明,光强通过改变茎秆形态和材料力学性状影响茎秆的抗倒伏能力。随着光强的升高,基部节间缩短,茎壁厚度、节间粗度和充实度均有不同程度的增加。相关分析显示,倒伏指数与节间长度呈极显著正相关,与节间粗度、茎壁厚度和充实度呈极显著的负相关。逐步回归分析表明,节间长度、茎壁厚度以及单位体积节间干质量是影响倒伏指数的关键因子。
缩短基部节间长度对提高茎秆的抗折力和抗倒伏能力具有重要作用[28],而光是影响植物节间伸长的重要因素[29]。光强可影响植株内激素的产生、活性和分布,低光强提高赤霉素活性,促进节间伸长[24]。结合本试验结果来看,在基部3个节间伸长期,随着光强增加,伸长节间的长度均缩短,抗折力有所提高,因此高光强下基部节间缩短是水稻抗倒伏能力增强的重要原因之一。
植株茎秆机械组织的厚薄直接影响节间的粗细和抗折力,茎壁越厚则维管束越多,抗折力越强[30],而环境光强能调控维管组织横向和纵向发育的平衡,对茎壁厚度具有重要影响。纤维素和木质素是细胞壁和茎壁的主要成分,增加纤维素和木质素含量可提高茎壁的机械强度和韧度[25,31]。光强不足将影响细胞壁的发育和木质化程度,降低茎壁厚度[32]。本研究结果表明,随着光强增加,正在伸长节间的茎壁厚度有所增加,说明光强提高有利于增加茎壁厚度,提高茎杆抗倒伏能力。
单位体积节间干质量反映节间充实度的高低。节间贮存的碳水化合物与茎秆充实度密切相关,而碳水化合物的多少对维持茎秆强度有重要作用[33]。充足的光照能促进碳水化合物的积累以及纤维素和木质素的生成,增加茎壁厚度[32,34]。在本研究中,高光强处理下节间干质量和充实度均显著提高,茎秆抗倒伏能力增强,表明基部节间充实度对水稻抗倒伏能力有较大作用。
郭玉明等[35]研究表明,抗弯刚度和弹性模量对作物抗倒伏能力影响较小。本研究亦表明,抗折力随着光强增大而增强。倒伏指数与抗折力呈极显著负相关,与节间的弯曲力矩呈极显著正相关,但与抗弯刚度和弹性模量相关性不显著。因此在茎秆力学性状中,抗折力和弯曲力矩与水稻抗倒伏能力关系密切,而弹性模量和抗弯刚度对抗倒伏能力的影响较小。
根据上述结果,我们认为光强通过影响茎秆同化产物的合成和分配,改变节间长度、节间充实度和茎壁厚度,进而改变节间抗折力和弯曲力矩等材料力学性状,最终影响水稻的抗倒伏能力。
株高是影响抗倒伏性的重要因素,但在本研究中,倒伏指数与株高的相关性并未达到显著水平,这与张忠旭等[30]研究结果一致。从力学角度分析,重心高度比株高与抗倒伏能力的关系更为密切,重心上移使茎秆头重脚轻,易于倒伏。本研究中,倒伏指数与茎秆重心高度呈极显著正相关,随着光强增加,茎秆重心高度下降,抗倒伏能力增强。如何降低茎杆重心高度,在不降低株高的情况下提高抗倒伏能力,从而协调高产与抗倒的矛盾,值得进一步探讨。
3.3 水稻抗倒伏能力对光强反应的敏感期
已有研究表明,茎倒伏多发生于基部3个节间,因此提高基部3个节间的抗倒伏能力是解决倒伏问题的关键[3,7]。但对于倒伏最敏感的具体节位,不同研究者有不同的观点。杨艳华等[36]认为,倒伏的折断节位多发生于第1或第2节间。华泽田等[37]研究表明,发生倒伏的敏感节位多在茎秆基部的第2和第3节间。不同结果可能是供试品种以及材料种植方式不同所致。
本研究采用不同处理间的变异系数作为反映抗倒伏能力对光强处理敏感性大小的指标。结果表明,在第2节间伸长期间进行不同光强处理,倒伏指数变异系数明显大于其他节间伸长期间处理,表明第2节间伸长期是对光强最敏感的时期。而多数性状的最大变异系数也出现在第2节间伸长期处理。已有研究亦表明,基部第2节间短而粗的品种具有较强的抗倒伏性[38],在生产实践中,水稻倒伏大多都发生在第2节间[13,21,39-40]。综合上述结果,我们认为水稻抗倒伏能力对光强反应最敏感的时期是第2节间伸长期。在生产实践中,应通过氮肥后移、适度晒田等措施,提高基部节间伸长期的群体透光率,避免基部节间(尤其是第2节间)过度伸长,从而达到减少倒伏的目的。
3.4 光强对水稻抗倒伏能力影响的累积效应
目前为止,关于光强对水稻抗倒伏能力影响的研究尚未考虑到处理时间的累积效应。本研究发现,在基部3个节间伸长期进行连续光强处理的情况下,大多数形态和材料力学性状的变异系数均明显高于单个节间伸长期处理。由此可见,光强对茎秆抗倒伏能力的影响有累积效应。光强对节间的影响,不仅作用于该节间的伸长期间,在该节间伸长过程完成以后仍有影响,这可能是影响了节间充实度和机械组织横向发育过程,其机理仍需进一步研究。此外,本研究发现,随着第1和第2节间伸长期光强的增加,第3节间不仅没有缩短,反而拉长,且差异达到显著水平,茎壁厚度和节间粗度则反而降低。谷海东等[41]也发现类似现象,其原因值得进一步探究。
[1]ISLAM M S, PENG S B, VISPERAS R M, et al.Lodging-related morphological traits of hybrid rice in a tropical irrigated ecosystem[J]. Field Crops Res, 2007,101: 240-248.
[2]SETTER T L, LAURELES E V, MAZAREDO A M.Lodging reduces yield of rice by self-shading and reductions in canopy photosynthesis[J]. Field Crops Res, 1997,49: 95-106.
[3]申广勒, 石英尧, 黄艳玲, 等. 水稻抗倒伏特性及其与茎秆性状的相关性研究[J]. 中国农学通报, 2007, 23: 58-62.
[4]ZHANG W J, WU L M, WU X R, et al. Lodging resistance of japonica rice (Oryza Sativa L.): Morphological and anatomical traits due to top-dressing nitrogen application rates[J]. Rice, 2016, 9: 1-11.
[5]ZHANG J, LI G, SONG Y, et al. Lodging resistance characteristics of high-yielding rice populations[J]. Field Crops Res, 2014, 161: 64-74.
[6]陈友订, 万邦惠, 张旭. 华南双季超高产水稻抽穗期理想株型结构研究[J]. 中国水稻科学, 2005, 19(1): 52-58.
[7]张秋英, 欧阳由男, 戴伟民, 等. 水稻基部伸长节间性状与倒伏相关性分析及QTL定位[J]. 作物学报, 2005, 31(6): 712-717.
[8]郑亭, 陈溢, 樊高琼, 等. 株行配置对带状条播小麦群体光环境及抗倒伏性能的影响[J]. 中国农业科学, 2013, 46(8): 1571-1582.
[9]杨世民, 谢力, 郑顺林, 等. 氮肥水平和栽插密度对杂交稻茎秆理化特性与抗倒伏性的影响[J]. 作物学报, 2009,35(1): 93-103.
[10]ZHANG W J, LI G H, YANG Y M, et al. Effects of nitrogen application rate and ratio on lodging resistance of super rice with different genotypes[J]. J Integr Agr, 2014,13(1): 63-72.
[11]MILLER F L. Studies on the relationships of lodging stem physical character and fertilization in wheat[J].Crop Sci, 1963(6): 468-471.
[12]HWANG S J, HAMAYUN M, KIM H Y, et al. Effect of nitrogen and silicon nutrition on bioactive gibberellin and growth of rice under field conditions[J]. J Crop Sci Biotech, 2007, 10(4): 281-286.
[13]李国辉, 钟旭华, 田卡, 等. 施氮对水稻茎秆抗倒伏能力的影响及其形态和力学机理[J]. 中国农业科学, 2013, 46(7): 1323-1334.
[14]张明聪, 刘元英, 罗盛国, 等. 养分综合管理对寒地水稻抗倒伏性能的影响[J]. 中国农业科学, 2010, 43(21): 4536-4542.
[15]王振昌, 郭相平, 杨静晗, 等. 旱涝交替胁迫对水稻干物质生产分配及倒伏性状的影响[J]. 农业工程学报, 2016,32(24): 114-123.
[16]王抄抄, 孔雷蕾, 李妹娟, 等. 分蘖期控水处理对超级稻产量和生理特性的影响[J]. 华北农学报, 2015, 30(5):146-152.
[17]KAHLEN K, STÜTZEL H. Simplification of a lightbased model for estimating final internode length in greenhouse cucumber canopies[J]. Ann Bot, 2011, 108(6): 1055-1063.
[18]戴云云, 丁艳锋, 刘正辉, 等. 花后水稻穗部夜间远红外增温处理对稻米品质的影响[J]. 中国水稻科学, 2009, 23(4): 414-420.
[19]NIKLAS K J. A statistical approach to biological factors of safety: Bending and shearing in Psilotum axes[J]. Ann Bot, 1998, 82: 177-187.
[20]濑古秀生. 水稻の倒伏に关する研究[J]. 九州农试学报,1962, 7: 419-495.
[21]艾治勇. 超级杂交稻形态及生理特性与抗倒性关系的研究[D]. 长沙: 湖南农业大学, 2006.
[22]陈喜凤, 孙宁, 谷岩, 等. 不同群体结构下大豆植株抗倒性能的比较[J]. 华南农业大学学报, 2015, 36(1): 33-41.
[23]XUE J, GOU L, ZHAO Y S, et al. Effects of light intensity within the canopy on maize lodging[J]. Field Crops Res, 2016, 188: 133-141.
[24]SPARKES D L, KING M. Disentangling the effects of PAR and R: FR on lodging associated characters of wheat (Triticum aestivum)[J]. Ann Appl Biol, 2008, 152:1-9.
[25]OOKAWA T, ISHIHARA K. Varietal difference of the cell wall components affecting the bending stress of the culm relating to the lodging resistance in paddy rice[J].Jpn J Crop Sci, 1993, 62: 378-384.
[26]吴耀民, 卓亚男. 水稻倒伏及栽培技术对策[J]. 垦殖与稻作, 1999(3): 12-14.
[27]闫川, 丁艳峰, 王强盛, 等. 行株距配置对水稻茎秆形态生理与群体生态的影响[J]. 中国水稻科学, 2007, 21(5):530-536.
[28]李杰, 张洪程, 龚金龙, 等. 不同种植方式对超级稻植株抗倒伏能力的影响[J]. 中国农业科学, 2011, 44(11):2234-2243.
[29]王小菁, 潘瑞炽. 红光、远红光、钙及IAA对绿豆下胚轴切段伸长的影响[J]. 植物生理学通讯, 1990, 26(5): 13-16.
[30]张忠旭, 陈温福, 杨振玉, 等. 水稻抗倒伏能力与茎秆物理性状的关系及其对产量的影响[J]. 沈阳农业大学学报, 1999, 30(2): 81-85.
[31]LIU W, DENG Y, HUSSAIN S, et al. Relationship between cellulose accumulation and lodging resistance in the stem of relay intercropped soybean [Glycine max (L.)Merr. ][J]. Field Crops Res, 2016, 196: 261-267.
[32]CAMPBELL C A, READ D W L. Influence of air temperature, light intensity and soil moisture on the growth,yield and some growth analysis characteristics of Chinook wheat grown in the growth chamber[J]. Can J Plant Sci, 1968, 48(3): 299-311.
[33]徐正进, 张树林, 周淑清, 等. 水稻穗型与抗倒伏性关系的初步分析[J]. 植物生理学通讯, 2004, 40(5): 561-563.
[34]田保明, 杨光圣, 曹刚强, 等. 农作物倒伏及其影响因素分析[J]. 中国农学通报, 2006, 22(4): 163-167.
[35]郭玉明, 袁红梅, 阴妍, 等. 茎秆作物抗倒伏生物力学评价研究及关联分析[J]. 农业工程学报, 2007, 23(7): 14-18.
[36]杨艳华, 朱镇, 张亚东, 等. 不同水稻品种(系)抗倒伏能力与茎秆形态性状的关系[J]. 江苏农业学报, 2011, 27(2): 231-235.
[37]华泽田, 郝宪彬, 沈枫, 等. 东北地区超级杂交粳稻倒伏性状的研究[J]. 沈阳农业大学学报, 2003, 34(3): 161-164.
[38]陈书强, 杜晓东, 杨丽敏, 等. 水稻倒伏粒重损失及不同品种抗倒能力差异比较[J]. 黑龙江农业科学, 2015(10):34-37.
[39]杨静晗, 郭相平, 杨骕, 等. 旱涝交替胁迫对水稻抗折力的影响及其机理分析[J]. 山东农业大学学报(自然科学版), 2014, 45(3): 328-333.
[40]莫永生, 杨亲琼, 刘丕庆, 等. 高大韧稻的抗折力与茎秆结构的关系[J]. 作物杂志, 2007(2): 40-42.
[41]谷海东, 赵宏伟, 刘洋, 等. 插秧密度对寒地粳稻抗倒伏能力的影响[J]. 作物杂志, 2014(2): 101-106.
Effects of light intensity during internode elongation on rice lodging resistance and its mechanism
LI Xiaojuan1,2†, LIANG Kaiming1†, ZHONG Xuhua1, TIAN Ka1, PENG Shaobing2,HUANG Nongrong1, PAN Junfeng1, LIU Yanzhuo1
(1 Guangdong Key Laboratory of New Technology in Rice Breeding/Rice Research Institute, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, China; 2 College of Plant Science amp; Technology,Huazhong Agricultural University, Wuhan 430070, China)
【Objective】To explore the effects of light intensity during internode elongation on rice (Oryza sativa L.) lodging resistance.【Method】Rice plants were subjected to different light intensities in phytotrons at the 1st(S1), 2nd(S2), 3rd(S3) and 1stto 3rd(S123) internode elongation stages. The lodging resistance of rice stem and the morphological and material mechanical traits of basal internodes were measured.【Result】The lodging index (LI) of rice stem significantly decreased with the increasing of light intensity. LI of medium or high light intensity treatments was 34.8%–73.9% lower than that of low light intensity treatment at the S123internodes elongation stage. LI was significantly positively correlated with internode length, and negatively correlated with internode diameter, culm wall thickness and internode plumpness. The S2stage was the most sensitive period oflodging resistance to light intensity variation. Light intensity had accumulative effects on lodging resistance.The variation coefficient of lodging index at the S123stage was greater than those at the S1, S2and S3stages, and similar results were found in most morphological and material mechanical traits.【Conclusion】The key factors to determine the lodging resistance are internode length, culm wall thickness and internode plumpness. Light intensity can significantly change the three morphological traits of basal internodes, which further affect the material mechanical characteristics of rice stem and lodging resistance.
rice (Oryza sativa L. ); lodging index; light intensity; morphological trait; material mechanical trait
李晓娟, 梁开明, 钟旭华, 等. 拔节期光强对水稻抗倒伏能力的影响及机理[J]. 华南农业大学学报, 2017, 38(6): 34-43.
S511; S183
A
1001-411X(2017)06-0034-10
2017-01-20 优先出版时间:2017-11-06
优先出版网址:http://kns.cnki.net/kcms/detail/44.1110.S.20171106.1412.014.html
李晓娟(1988—),女,硕士研究生,E-mail: lixiaojuanhzau@163.com;梁开明(1979—),男,副研究员,博士,E-mail:kaiming-liang@163.com;† 对本文贡献相同; 通信作者: 钟旭华 (1963—),男,研究员,博士,E-mail:xzhong8@163.com
广东省自然科学基金重点项目(S2012020011043);863计划(2014AA10A605);广东省科技计划项目(2013B050800014,粤科规财字[2014]208号)
【责任编辑 周志红】
猜你喜欢
江西农业大学学报(2022年3期)2022-07-07
湖北农业科学(2022年1期)2022-02-11
农业机械学报(2019年4期)2019-04-29
江西农业大学学报(2018年5期)2018-11-22
畜牧与饲料科学(2018年5期)2018-06-13
果农之友(2017年10期)2018-02-08
中国果业信息(2018年12期)2018-01-17
热带农业科学(2017年9期)2017-10-23
农村农业农民·B版(2016年7期)2016-10-21
土壤与作物(2013年3期)2013-03-11
